
Atypical Bacilliredoxin AbxC Plays a Role in Responding to Oxidative Stress in Radiation-Resistant Bacterium Deinococcus radiodurans

 , , , ,
, , , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Conditions
2.2. Construction of Mutants and Plasmids
2.3. Measurement of BSH
2.4. Survival Assay
2.5. Protein Carbonylation Assay
2.6. Protein Purification
2.7. NADPH Consumption Assay
2.8. Quantification of Thiols
2.9. Western Blotting
2.10. Preparation of AbxC Affinity Column and Capturing of Target Proteins
3. Results and Discussions
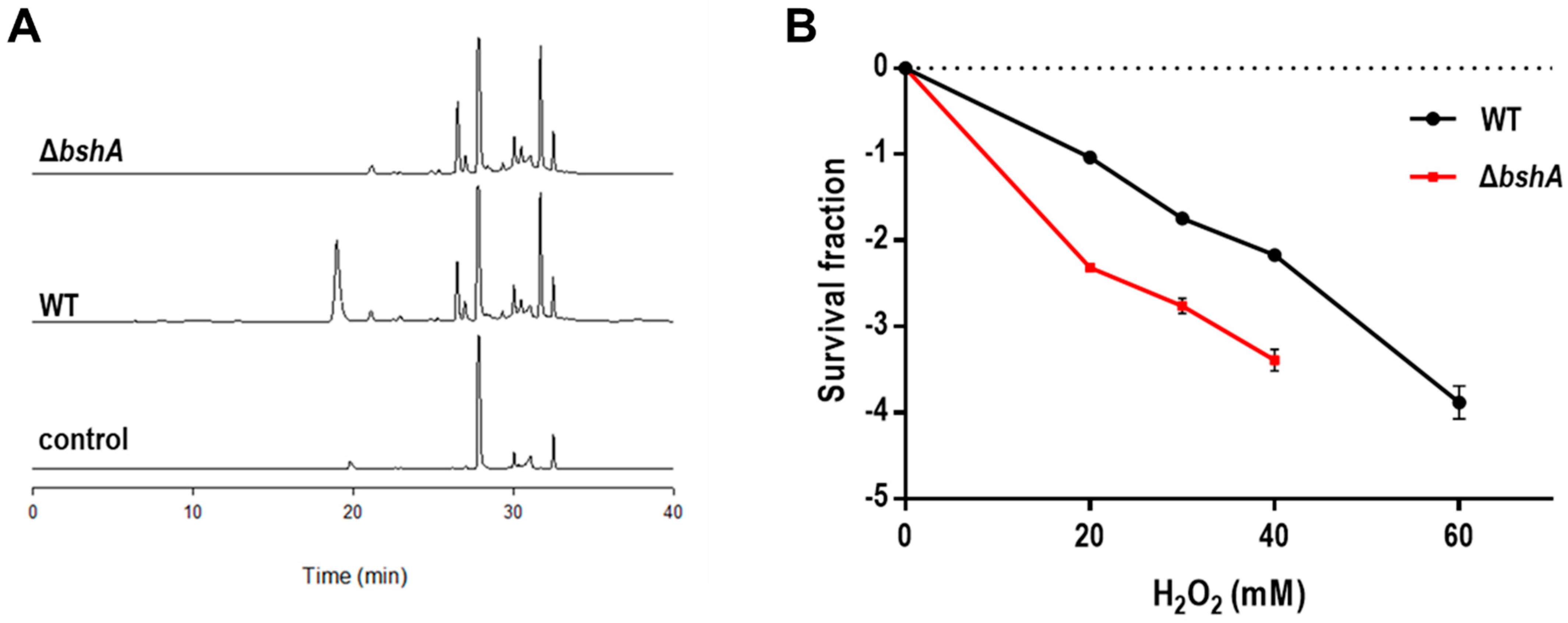
3.1. A BSH-Deficient D. radiodurans Strain Is Sensitive to H2O2
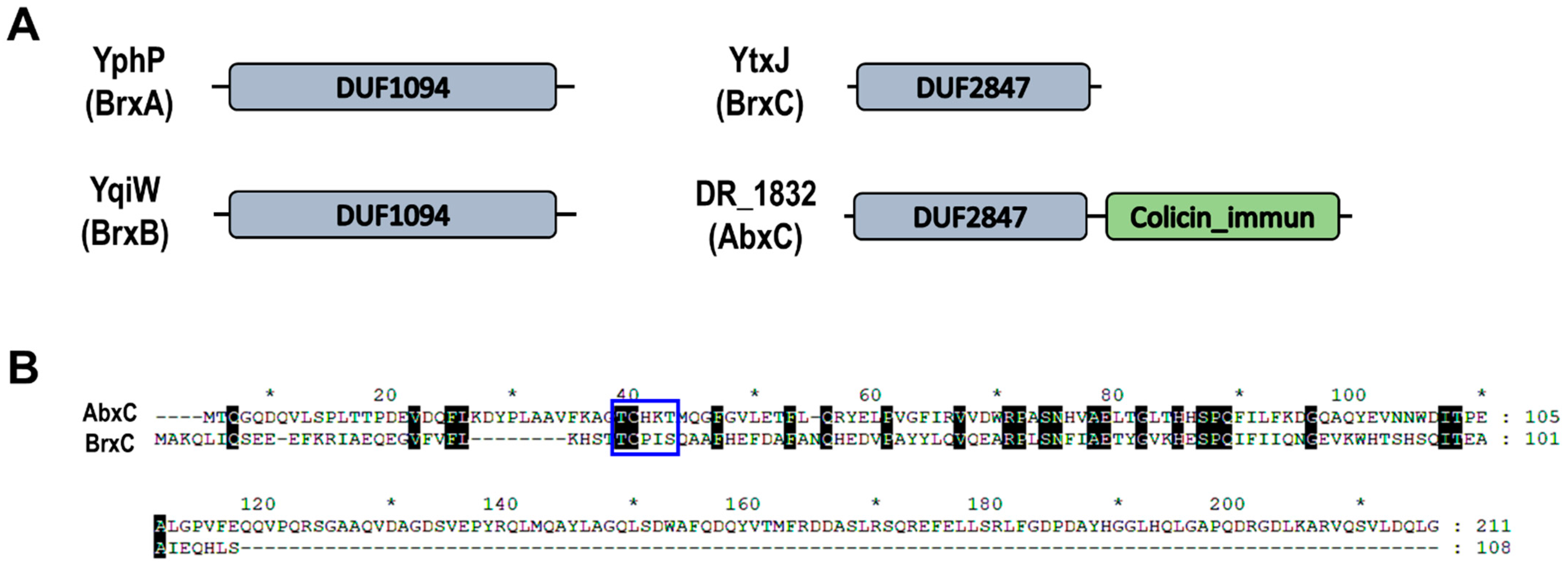
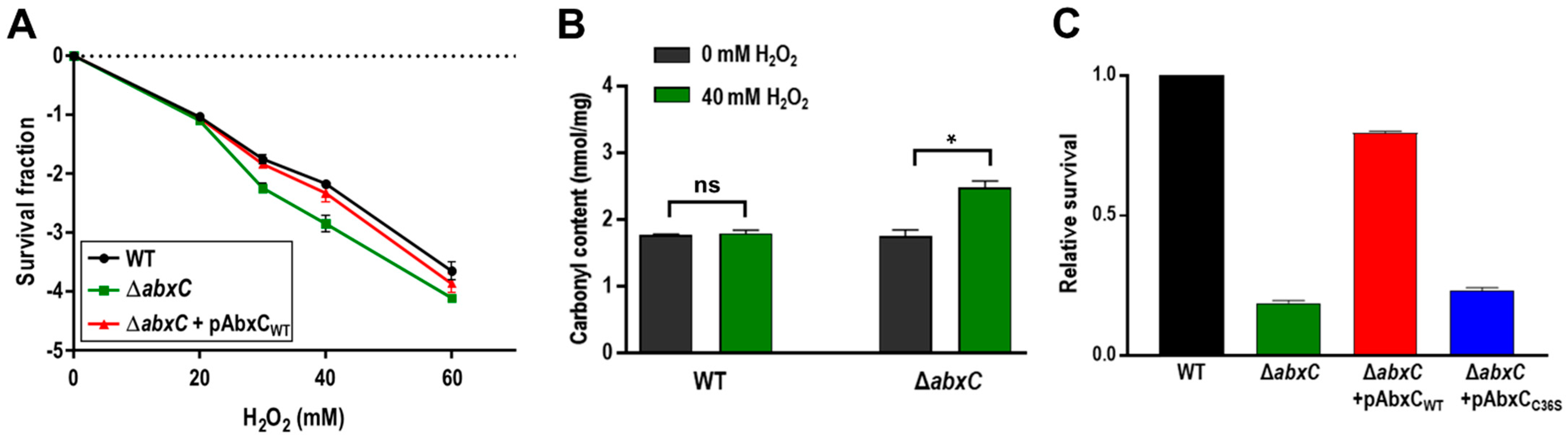
3.2. Mutation of abxC Decreases H2O2 Resistance of D. radiodurans
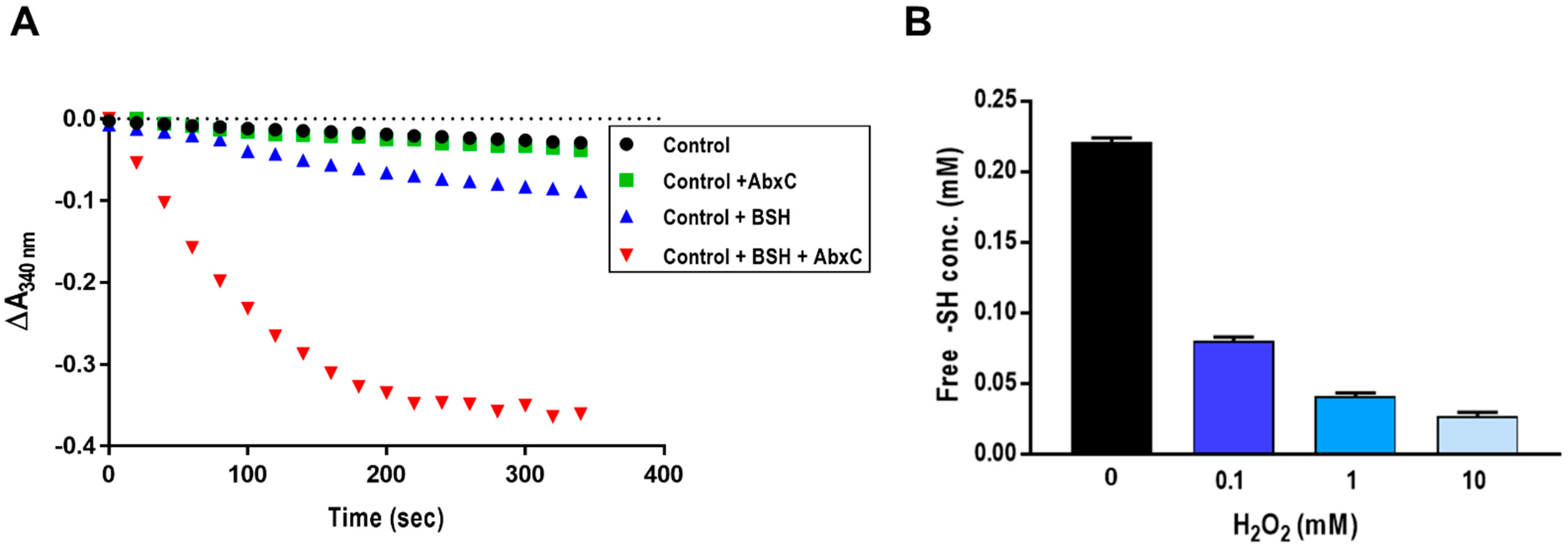
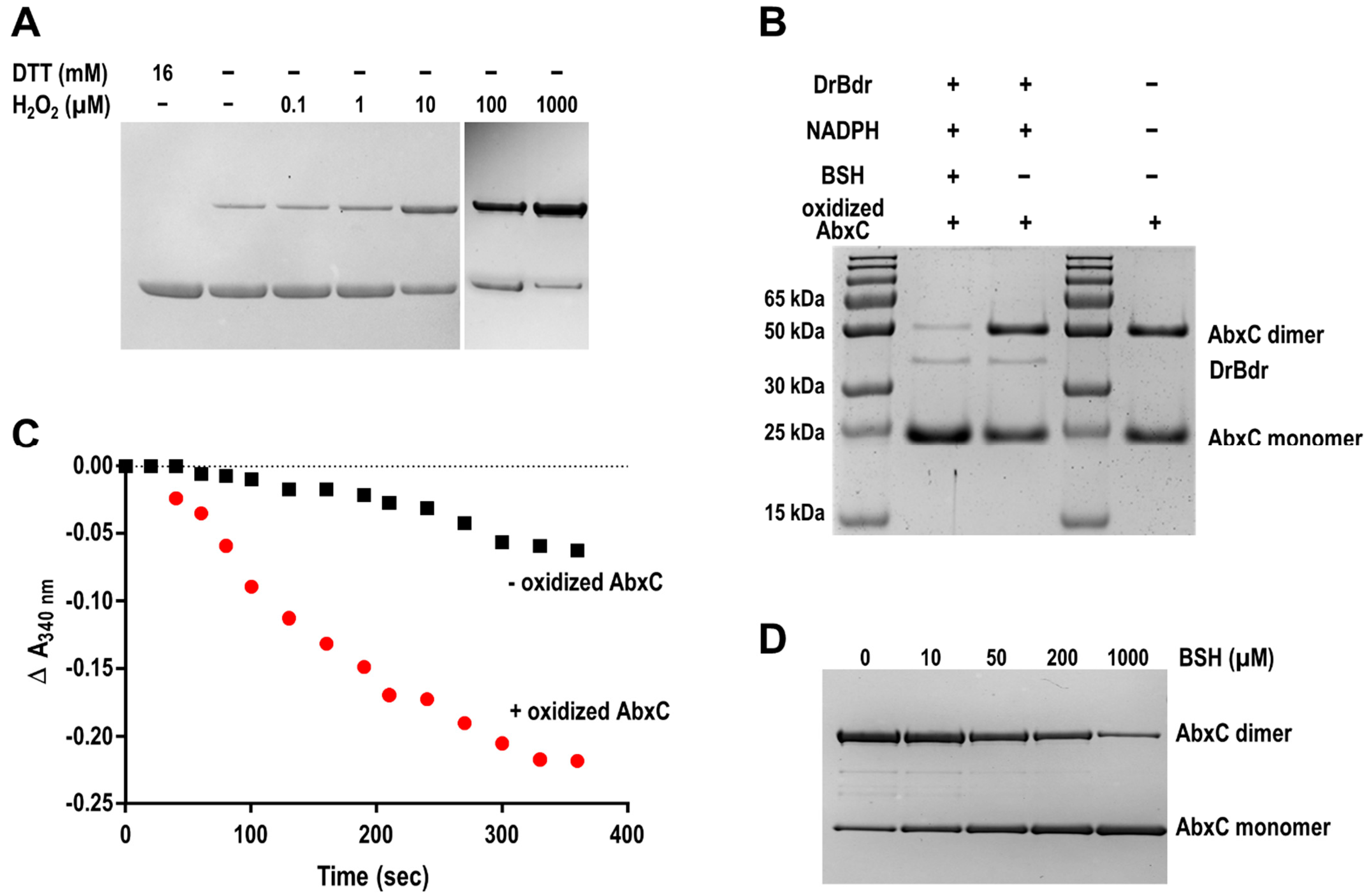
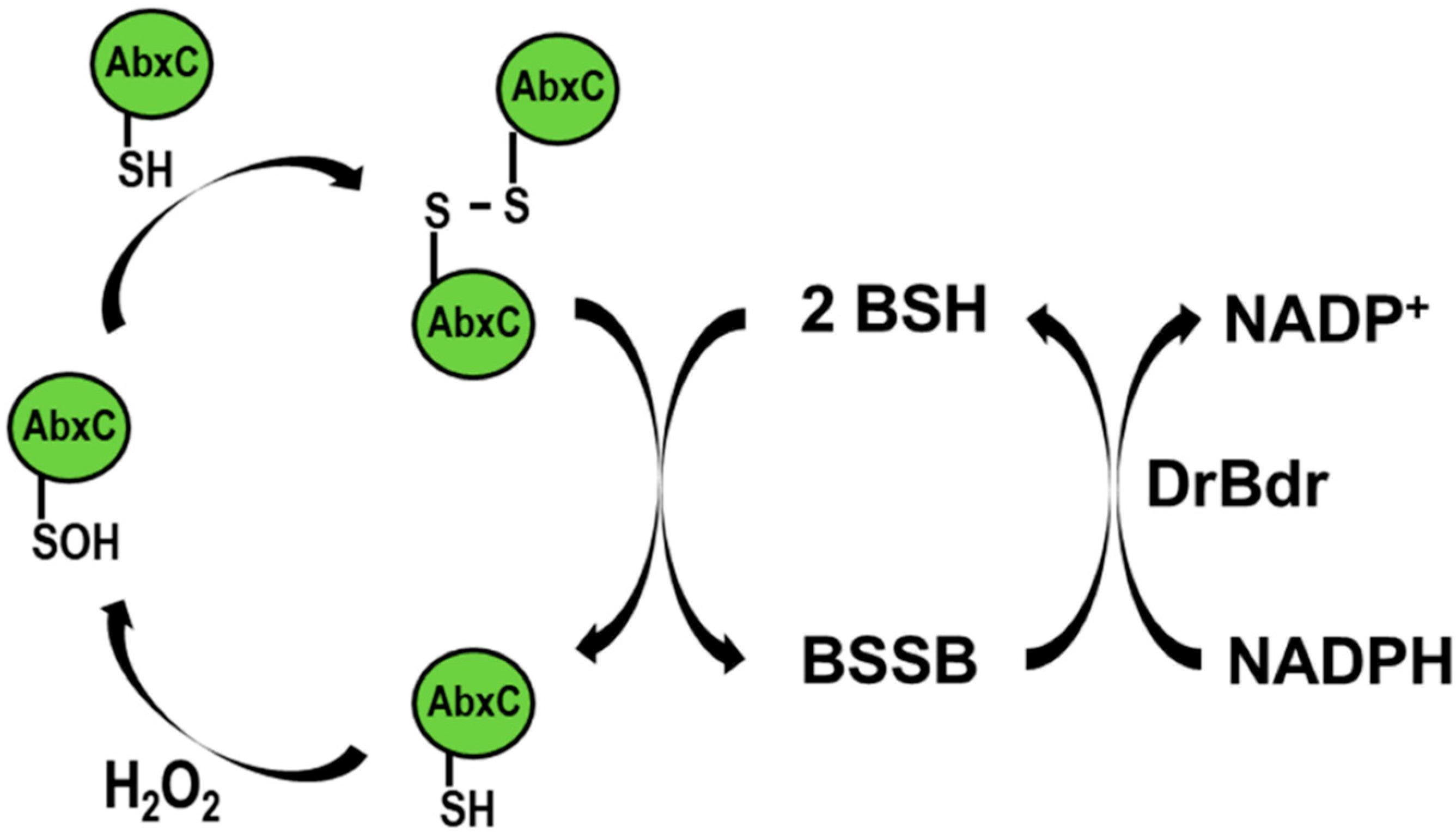
3.3. AbxC Exhibits H2O2 Scavenging Activity
3.4. H2O2 Stimulates AbxC Dimerization
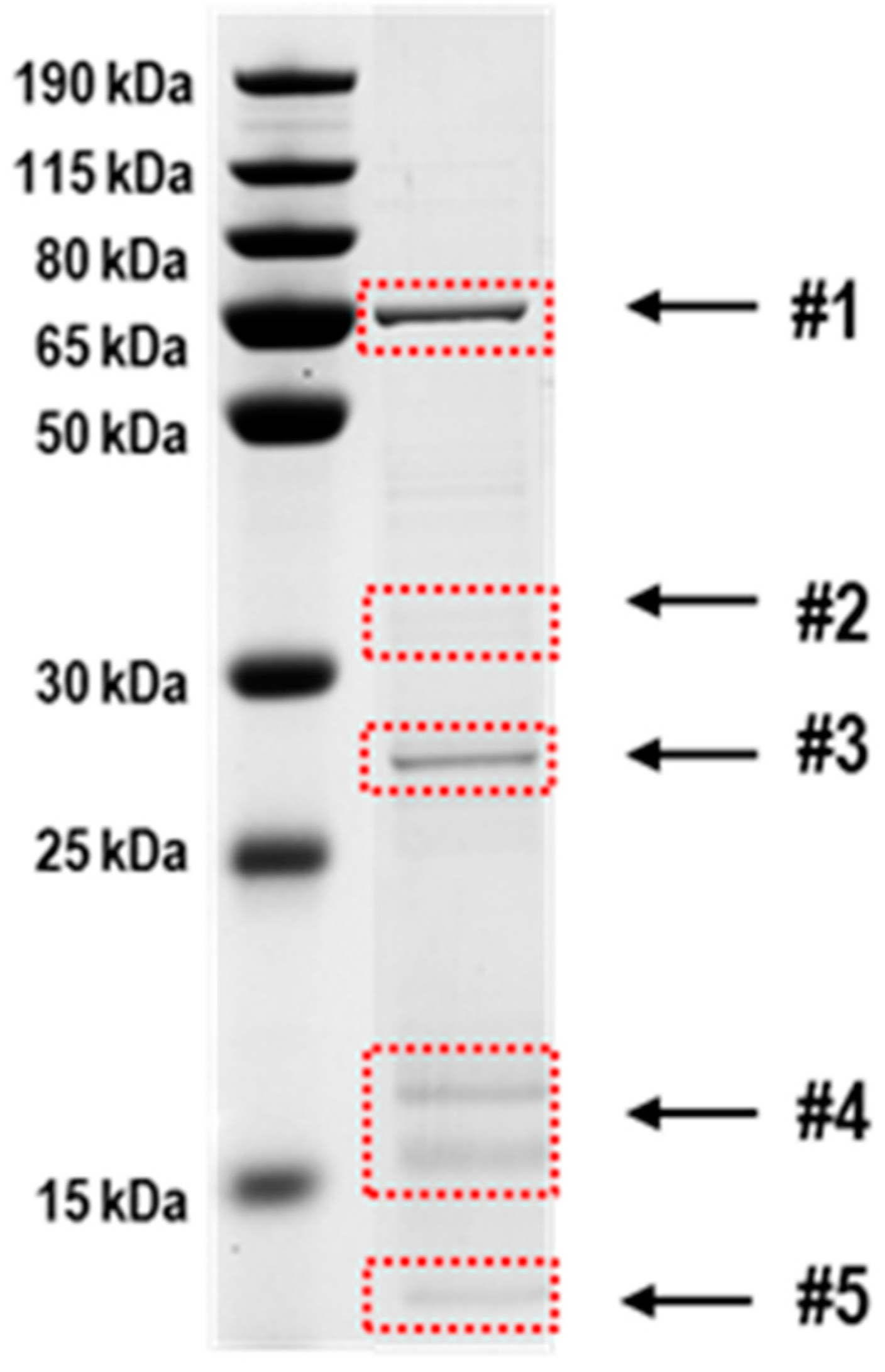
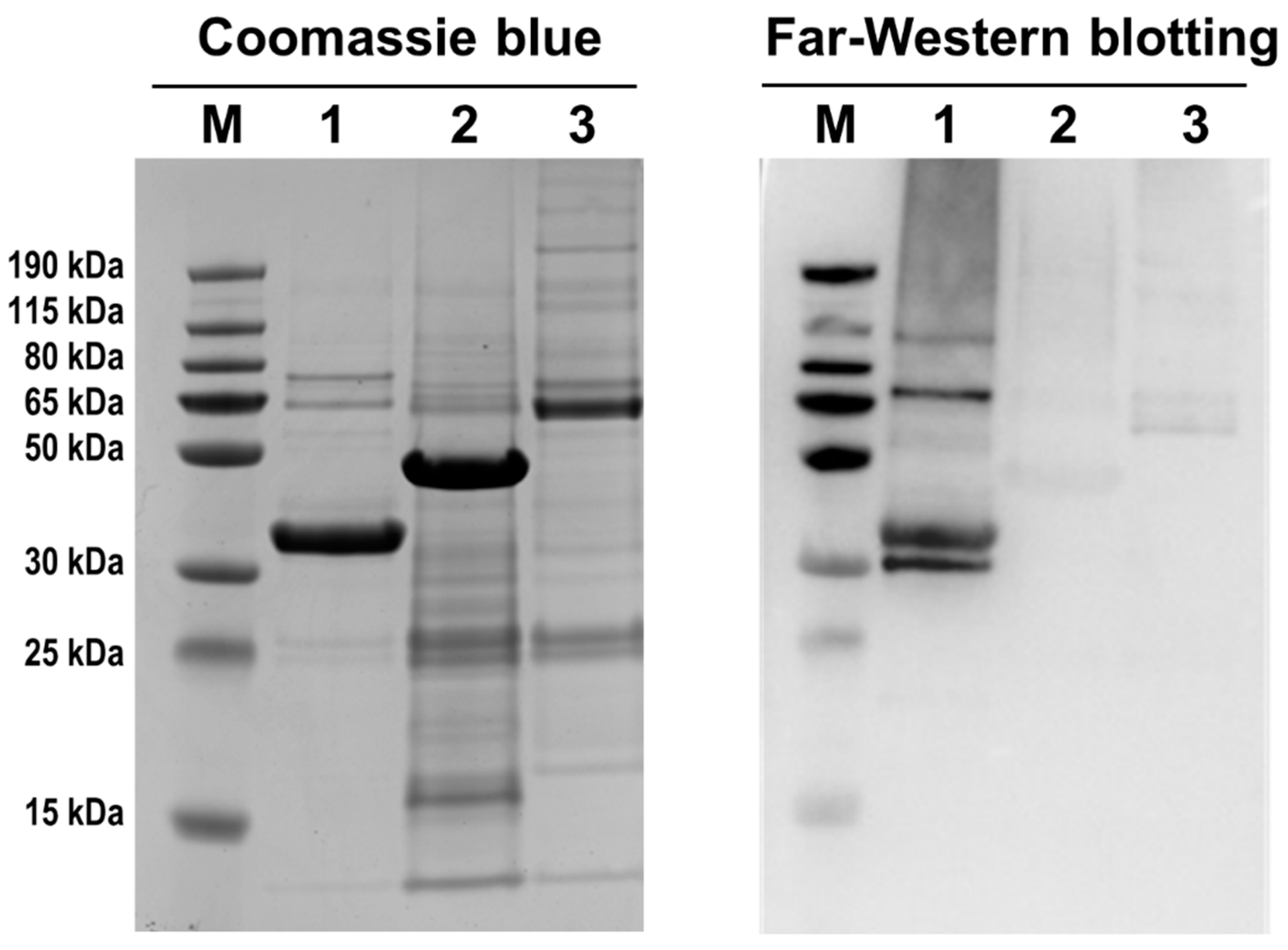
3.5. Identification of Proteins Potentially Interacting with AbxC
3.6. AbxC Interacts with DrOxyR
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Dalle-Donne, I.; Milzani, A.; Gagliano, N.; Colombo, R.; Giustarini, D.; Rossi, R. Molecular mechanisms and potential clinical significance of S-glutathionylation. Antioxid. Redox Signal. 2008, 10, 445–473. [Google Scholar] [CrossRef] [PubMed]
- Zamocky, M.; Gasselhuber, B.; Furtmuller, P.G.; Obinger, C. Molecular evolution of hydrogen peroxide degrading enzymes. Arch. Biochem. Biophys. 2012, 525, 131–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pannala, V.R.; Dash, R.K. Mechanistic characterization of the thioredoxin system in the removal of hydrogen peroxide. Free Radic. Biol. Med. 2015, 78, 42–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Xu, G.; Zhao, Y.; Tian, B.; Lu, H.; Yu, X.; Xu, Z.; Ying, N.; Hu, S.; Hua, Y. A novel OxyR sensor and regulator of hydrogen peroxide stress with one cysteine residue in Deinococcus radiodurans. PLoS ONE 2008, 3, e1602. [Google Scholar] [CrossRef] [PubMed]
- Basu, B.; Apte, S.K. Gamma radiation-induced proteome of Deinococcus radiodurans primarily targets DNA repair and oxidative stress alleviation. Mol. Cell Proteomics. 2012, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S.W.; Jung, J.H.; Kim, M.K.; Seo, H.S.; Lim, H.M.; Lim, S. The three catalases in Deinococcus radiodurans: Only two show catalase activity. Biochem. Biophys. Res. Commun. 2016, 469, 443–448. [Google Scholar] [CrossRef]
- Wood, Z.A.; Schroder, E.; Robin Harris, J.; Poole, L.B. Structure, mechanism and regulation of peroxiredoxins. Trends Biochem. Sci. 2003, 28, 32–40. [Google Scholar] [CrossRef]
- Lim, S.; Jung, J.H.; Blanchard, L.; de Groot, A. Conservation and diversity of radiation and oxidative stress resistance mechanisms in Deinococcus species. FEMS Microbiol. Rev. 2019, 43, 19–52. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Jeong, S.; Zhang, J.; Jung, J.H.; Choi, J.I.; Lim, S.; Kim, M.K. Crystal structure of the AhpD-like protein DR1765 from Deinococcus radiodurans R1. Biochem. Biophys. Res. Commun. 2020, 529, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Obiero, J.; Pittet, V.; Bonderoff, S.A.; Sanders, D.A. Thioredoxin system from Deinococcus radiodurans. J. Bacteriol. 2010, 192, 494–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, C.; Lee, G.W.; Hong, S.H.; Kaur, S.; Jung, K.W.; Jung, J.H.; Lim, S.; Chung, B.Y.; Lee, S.S. Novel functions of peroxiredoxin Q from Deinococcus radiodurans R1 as a peroxidase and a molecular chaperone. FEBS Lett. 2019, 593, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, Q.; Liu, W. The versatile low-molecular-weight thiols: Beyond cell protection. Bioessays 2015, 37, 1262–1267. [Google Scholar] [CrossRef] [PubMed]
- Loi, V.V.; Rossius, M.; Antelmann, H. Redox regulation by reversible protein S-thiolation in bacteria. Front. Microbiol. 2015, 6, 187. [Google Scholar] [CrossRef] [Green Version]
- Newton, G.L.; Rawat, M.; La Clair, J.J.; Jothivasan, V.K.; Budiarto, T.; Hamilton, C.J.; Claiborne, A.; Helmann, J.D.; Fahey, R.C. Bacillithiol is an antioxidant thiol produced in Bacilli. Nat. Chem. Biol. 2009, 5, 625–627. [Google Scholar] [CrossRef] [PubMed]
- Linzner, N.; Loi, V.V.; Fritsch, V.N.; Tung, Q.N.; Stenzel, S.; Wirtz, M.; Hell, R.; Hamilton, C.J.; Tedin, K.; Fulde, M.; et al. Staphylococcus aureus Uses the Bacilliredoxin (BrxAB)/Bacillithiol Disulfide Reductase (YpdA) Redox Pathway to Defend Against Oxidative Stress Under Infections. Front. Microbiol. 2019, 10, 1355. [Google Scholar] [CrossRef] [Green Version]
- Dickerhof, N.; Paton, L.; Kettle, A.J. Oxidation of bacillithiol by myeloperoxidase-derived oxidants. Free Radic. Biol. Med. 2020, 158, 74–83. [Google Scholar] [CrossRef]
- Chandrangsu, P.; Loi, V.V.; Antelmann, H.; Helmann, J.D. The Role of Bacillithiol in Gram-Positive Firmicutes. Antioxid. Redox Signal. 2018, 28, 445–462. [Google Scholar] [CrossRef] [Green Version]
- Gaballa, A.; Chi, B.K.; Roberts, A.A.; Becher, D.; Hamilton, C.J.; Antelmann, H.; Helmann, J.D. Redox regulation in Bacillus subtilis: The bacilliredoxins BrxA(YphP) and BrxB(YqiW) function in de-bacillithiolation of S-bacillithiolated OhrR and MetE. Antioxid. Redox Signal. 2014, 21, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Imber, M.; Huyen, N.T.T.; Pietrzyk-Brzezinska, A.J.; Loi, V.V.; Hillion, M.; Bernhardt, J.; Tharichen, L.; Kolsek, K.; Saleh, M.; Hamilton, C.J.; et al. Protein S-Bacillithiolation Functions in Thiol Protection and Redox Regulation of the Glyceraldehyde-3-Phosphate Dehydrogenase Gap in Staphylococcus aureus Under Hypochlorite Stress. Antioxid. Redox Signal. 2018, 28, 410–430. [Google Scholar] [CrossRef] [Green Version]
- Gaballa, A.; Newton, G.L.; Antelmann, H.; Parsonage, D.; Upton, H.; Rawat, M.; Claiborne, A.; Fahey, R.C.; Helmann, J.D. Biosynthesis and functions of bacillithiol, a major low-molecular-weight thiol in Bacilli. Proc. Natl. Acad Sci. USA 2010, 107, 6482–6486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaballa, A.; Su, T.T.; Helmann, J.D. The Bacillus subtilis monothiol bacilliredoxin BrxC (YtxJ) and the Bdr (YpdA) disulfide reductase reduce S-bacillithiolated proteins. Redox Biol. 2021, 101935. [Google Scholar] [CrossRef] [PubMed]
- Hammerstad, M.; Gudim, I.; Hersleth, H.P. The Crystal Structures of Bacillithiol Disulfide Reductase Bdr (YpdA) Provide Structural and Functional Insight into a New Type of FAD-Containing NADPH-Dependent Oxidoreductase. Biochemistry 2020, 59, 4793–4798. [Google Scholar] [CrossRef]
- Joe, M.H.; Lee, K.H.; Lim, S.Y.; Im, S.H.; Song, H.P.; Lee, I.S.; Kim, D.H. Pigment-based whole-cell biosensor system for cadmium detection using genetically engineered Deinococcus radiodurans. Bioprocess. Biosyst. Eng. 2012, 35, 265–272. [Google Scholar] [CrossRef]
- Udupa, K.S.; O’Cain, P.A.; Mattimore, V.; Battista, J.R. Novel ionizing radiation-sensitive mutants of Deinococcus radiodurans. J. Bacteriol. 1994, 176, 7439–7446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohba, H.; Satoh, K.; Yanagisawa, T.; Narumi, I. The radiation responsive promoter of the Deinococcus radiodurans pprA gene. Gene 2005, 363, 133–141. [Google Scholar] [CrossRef]
- Jeong, S.W.; Kim, M.K.; Zhao, L.; Yang, S.K.; Jung, J.H.; Lim, H.M.; Lim, S. Effects of Conserved Wedge Domain Residues on DNA Binding Activity of Deinococcus radiodurans RecG Helicase. Front. Genet. 2021, 12, 634615. [Google Scholar] [CrossRef]
- Meima, R.; Lidstrom, M.E. Characterization of the minimal replicon of a cryptic Deinococcus radiodurans SARK plasmid and development of versatile Escherichia coli-D. radiodurans shuttle vectors. Appl. Environ. Microbiol. 2000, 66, 3856–3867. [Google Scholar] [CrossRef] [Green Version]
- Newton, G.L.; Fahey, R.C.; Rawat, M. Detoxification of toxins by bacillithiol in Staphylococcus aureus. Microbiology 2012, 158, 1117–1126. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.S.; Xiong, Y.Q.; Mitchell, J.; Seepersaud, R.; Bayer, A.S.; Sullam, P.M. Bacteriophage lysin mediates the binding of streptococcus mitis to human platelets through interaction with fibrinogen. PLoS Pathog. 2010, 6, e1001047. [Google Scholar] [CrossRef]
- Moreno-Sanchez, R.; Marin-Hernandez, A.; Gallardo-Perez, J.C.; Vazquez, C.; Rodriguez-Enriquez, S.; Saavedra, E. Control of the NADPH supply and GSH recycling for oxidative stress management in hepatoma and liver mitochondria. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 1138–1150. [Google Scholar] [CrossRef] [PubMed]
- Winther, J.R.; Thorpe, C. Quantification of thiols and disulfides. Biochim. Biophys. Acta 2014, 1840, 838–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, K.; Tu, Z.; Ohba, H.; Narumi, I. Development of versatile shuttle vectors for Deinococcus grandis. Plasmid 2009, 62, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, Q.; Chen, X.Z. Detecting protein-protein interactions by Far western blotting. Nat. Protoc. 2007, 2, 3278–3284. [Google Scholar] [CrossRef] [PubMed]
- Sturm, N.; Jortzik, E.; Mailu, B.M.; Koncarevic, S.; Deponte, M.; Forchhammer, K.; Rahlfs, S.; Becker, K. Identification of proteins targeted by the thioredoxin superfamily in Plasmodium falciparum. PLoS Pathog. 2009, 5, e1000383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahm, J.Y.; Kim, J.Y.; Park, J.W.; Kang, J.Y.; Kim, K.B.; Kim, S.R.; Cho, H.; Seo, S.B. Methylation of UHRF1 by SET7 is essential for DNA double-strand break repair. Nucleic. Acids. Res. 2019, 47, 184–196. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kang, J.; Kang, Y.L.; Woo, J.; Kim, Y.; Huh, J.; Park, J.W. Ketohexokinase-A acts as a nuclear protein kinase that mediates fructose-induced metastasis in breast cancer. Nat. Commun. 2020, 11, 5436. [Google Scholar] [CrossRef]
- Posada, A.C.; Kolar, S.L.; Dusi, R.G.; Francois, P.; Roberts, A.A.; Hamilton, C.J.; Liu, G.Y.; Cheung, A. Importance of bacillithiol in the oxidative stress response of Staphylococcus aureus. Infect. Immun. 2014, 82, 316–332. [Google Scholar] [CrossRef] [Green Version]
- Pedre, B.; Van Molle, I.; Villadangos, A.F.; Wahni, K.; Vertommen, D.; Turell, L.; Erdogan, H.; Mateos, L.M.; Messens, J. The Corynebacterium glutamicum mycothiol peroxidase is a reactive oxygen species-scavenging enzyme that shows promiscuity in thiol redox control. Mol. Microbiol. 2015, 96, 1176–1191. [Google Scholar] [CrossRef]
- Ulrich, K.; Jakob, U. The role of thiols in antioxidant systems. Free Radic. Biol. Med. 2019, 140, 14–27. [Google Scholar] [CrossRef]
- Perera, V.R.; Newton, G.L.; Pogliano, K. Bacillithiol: A key protective thiol in Staphylococcus aureus. Expert Rev. Anti. Infect. Ther. 2015, 13, 1089–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rungrassamee, W.; Liu, X.; Pomposiello, P.J. Activation of glucose transport under oxidative stress in Escherichia coli. Arch. Microbiol. 2008, 190, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Alfano, J.R.; Becker, D.F. Proline metabolism increases katG expression and oxidative stress resistance in Escherichia coli. J. Bacteriol. 2015, 197, 431–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanz, N.D.; Booker, S.J. Auxiliary iron-sulfur cofactors in radical SAM enzymes. Biochim. Biophys. Acta 2015, 1853, 1316–1334. [Google Scholar] [CrossRef] [Green Version]
- Makarova, K.S.; Aravind, L.; Daly, M.J.; Koonin, E.V. Specific expansion of protein families in the radioresistant bacterium Deinococcus radiodurans. Genetica 2000, 108, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Meunier-Jamin, C.; Kapp, U.; Leonard, G.A.; McSweeney, S. The structure of the organic hydroperoxide resistance protein from Deinococcus radiodurans. Do conformational changes facilitate recycling of the redox disulfide? J. Biol. Chem. 2004, 279, 25830–25837. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Quinn, A.M.; Wolin, S.L. Ro ribonucleoproteins contribute to the resistance of Deinococcus radiodurans to ultraviolet irradiation. Genes Dev. 2000, 14, 777–782. [Google Scholar]
- Goncalves, A.M.D.; de Sanctis, D.; McSweeney, S.M. Structural and functional insights into DR2231 protein, the MazG-like nucleoside triphosphate pyrophosphohydrolase from Deinococcus radiodurans. J. Biol. Chem. 2011, 286, 30691–30705. [Google Scholar] [CrossRef] [Green Version]
- Nagahara, N. Intermolecular disulfide bond to modulate protein function as a redox-sensing switch. Amino. Acids. 2011, 41, 59–72. [Google Scholar] [CrossRef]
- Luan, H.; Meng, N.; Fu, J.; Chen, X.; Xu, X.; Feng, Q.; Jiang, H.; Dai, J.; Yuan, X.; Lu, Y.; et al. Genome-wide transcriptome and antioxidant analyses on gamma-irradiated phases of Deinococcus radiodurans R1. PLoS ONE 2014, 9, e85649. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Locus tag | Protein | Size | Count * | No. of Cys |
|---|---|---|---|---|---|
| #1 | DR_1262 | 60 kDa SS-A/Ro ribonucleoprotein | 57 kDa | 5 | 2 |
| DR_1298 | radical SAM superfamily | 58 kDa | 6 | 5 | |
| DR_1316 | propionyl-CoA carboxylase | 57 kDa | 10 | 4 | |
| DR_1742 | glucose-6-phosphate isomerase | 60 kDa | 3 | 2 | |
| #2 | DR_0615 | transcriptional regulator OxyR | 35 kDa | 3 | 1 |
| DR_0814 | proline dehydrogenase | 35 kDa | 6 | 2 | |
| DR_1072 | acetyl-CoA acetyltransferase | 41 kDa | 7 | 4 | |
| DR_1022 | NTP pyrophosphatase MazG | 37 kDa | 8 | 3 | |
| DR_1519 | ketol-acid reductoisomerase | 37 kDa | 13 | 3 | |
| DR_1890 | oxidoreductase | 35 kDa | 3 | 4 | |
| DR_2428 | nicotinamide-nucleotide adenylyltransferase | 38 kDa | 3 | 1 | |
| DR_A0143 | 3-hydroxyacyl-CoA dehydrogenase | 38 kDa | 3 | 1 | |
| #3 | DR_0114 | enoyl-CoA hydratase, putative | 27 kDa | 7 | 2 |
| DR_1451 | S-adenosylhomocysteine nuclosidase | 25 kDa | 4 | 2 | |
| DR_1832 | putative homolog of BrxC (YtxJ) | 24 kDa | 13 | 1 | |
| #4 | DR_0763 | predicted acetyltransferase | 18 kDa | 3 | 1 |
| DR_0876 | Nudix family | 18 kDa | 3 | 2 | |
| DR_1245 | putative sensory transduction regulator | 19 kDa | 6 | 1 | |
| DR_1418 | signal transduction response regulator | 15 kDa | 5 | 2 | |
| DR_1654 | hypothetical protein | 19 kDa | 5 | 3 | |
| DR_1776 | Nudix family | 19 kDa | 4 | 3 | |
| DR_1857 | organic hydroperoxide reductase | 15 kDa | 3 | 2 | |
| DR_2298 | acyl-CoA thioester hydrolase | 19 kDa | 4 | 2 | |
| DR_2481 | hypothetical protein | 17 kDa | 5 | 1 | |
| #5 | DR_2580 | ribosomal silencing factor RsfS | 13 kDa | 6 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, S.; Jung, J.-H.; Kim, M.-K.; de Groot, A.; Blanchard, L.; Ryu, S.; Bahn, Y.-S.; Lim, S. Atypical Bacilliredoxin AbxC Plays a Role in Responding to Oxidative Stress in Radiation-Resistant Bacterium Deinococcus radiodurans. Antioxidants 2021, 10, 1148. https://doi.org/10.3390/antiox10071148
Jeong S, Jung J-H, Kim M-K, de Groot A, Blanchard L, Ryu S, Bahn Y-S, Lim S. Atypical Bacilliredoxin AbxC Plays a Role in Responding to Oxidative Stress in Radiation-Resistant Bacterium Deinococcus radiodurans. Antioxidants. 2021; 10(7):1148. https://doi.org/10.3390/antiox10071148
Chicago/Turabian StyleJeong, Soyoung, Jong-Hyun Jung, Min-Kyu Kim, Arjan de Groot, Laurence Blanchard, Sangryeol Ryu, Yong-Sun Bahn, and Sangyong Lim. 2021. "Atypical Bacilliredoxin AbxC Plays a Role in Responding to Oxidative Stress in Radiation-Resistant Bacterium Deinococcus radiodurans" Antioxidants 10, no. 7: 1148. https://doi.org/10.3390/antiox10071148
APA StyleJeong, S., Jung, J. -H., Kim, M. -K., de Groot, A., Blanchard, L., Ryu, S., Bahn, Y. -S., & Lim, S. (2021). Atypical Bacilliredoxin AbxC Plays a Role in Responding to Oxidative Stress in Radiation-Resistant Bacterium Deinococcus radiodurans. Antioxidants, 10(7), 1148. https://doi.org/10.3390/antiox10071148