
Changes in Antioxidative, Oxidoreductive and Detoxification Enzymes during Development of Aphids and Temperature Increase



Abstract
:1. Introduction
2. Materials and Methods
2.1. Aphids
2.2. Host Plants
2.3. Effect of Temperature on the Enzymatic Activity in Nymph and Adult Aphid Tissues
2.3.1. Antioxidant Enzymes
2.3.2. Detoxifying Enzymes
2.3.3. Oxidoreductive Enzymes
2.3.4. Protein Content in Aphid Extracts
2.4. Effect of Temperature on Aphid Longevity
2.5. Statistical Analyses
3. Results
3.1. Antioxidant Enzyme Activity of Nymphs and Adult Aphids during Temperature Increase
3.2. Detoxifying Enzyme Activity of Nymphs and Adult Aphids during Temperature Increase
3.3. Oxidoreductive Enzyme Activity of Nymphs and Adult Aphids during Temperature Increase
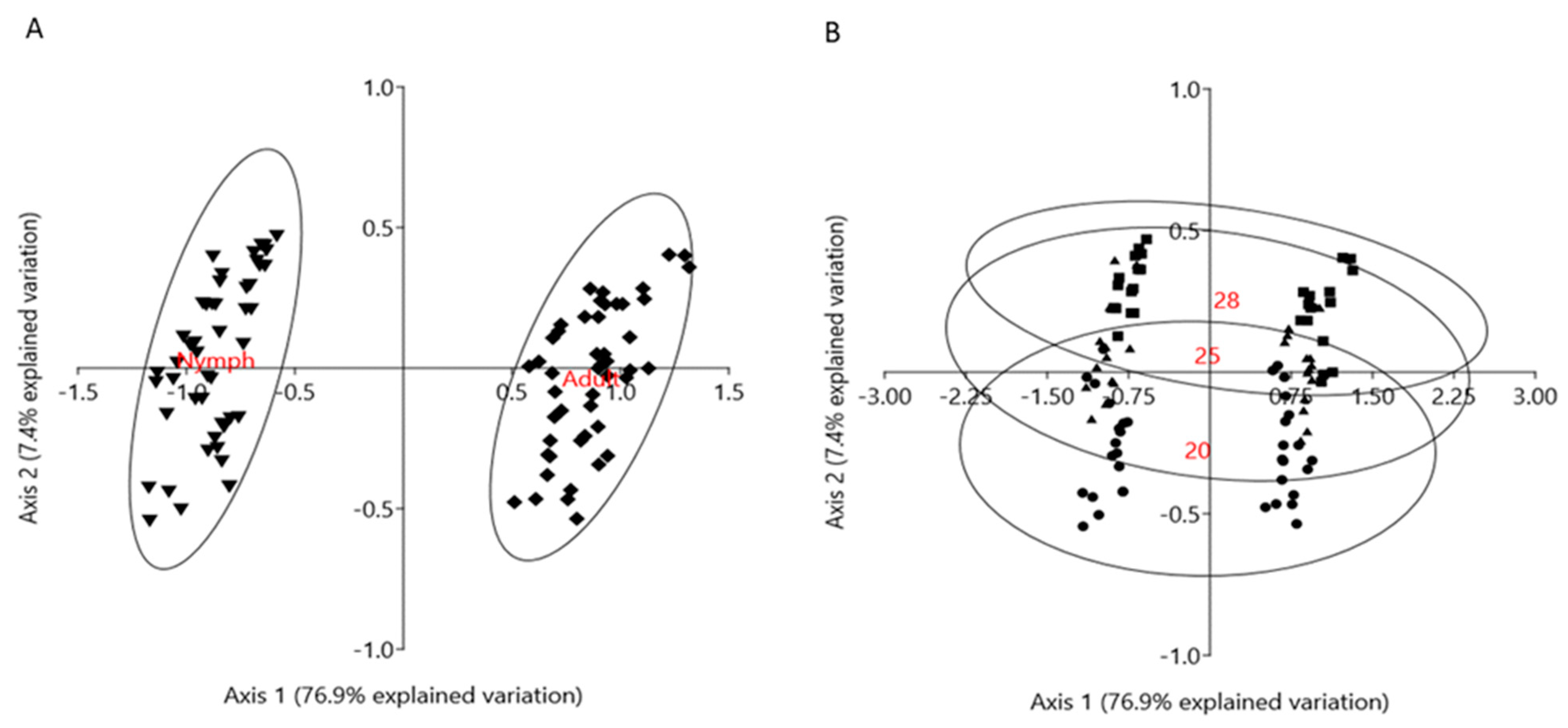
3.4. Enzymatic Differences between Nymphs and Adult Aphids during Temperature Increase
3.5. Effect of Temperature on the Longevity of Aphids
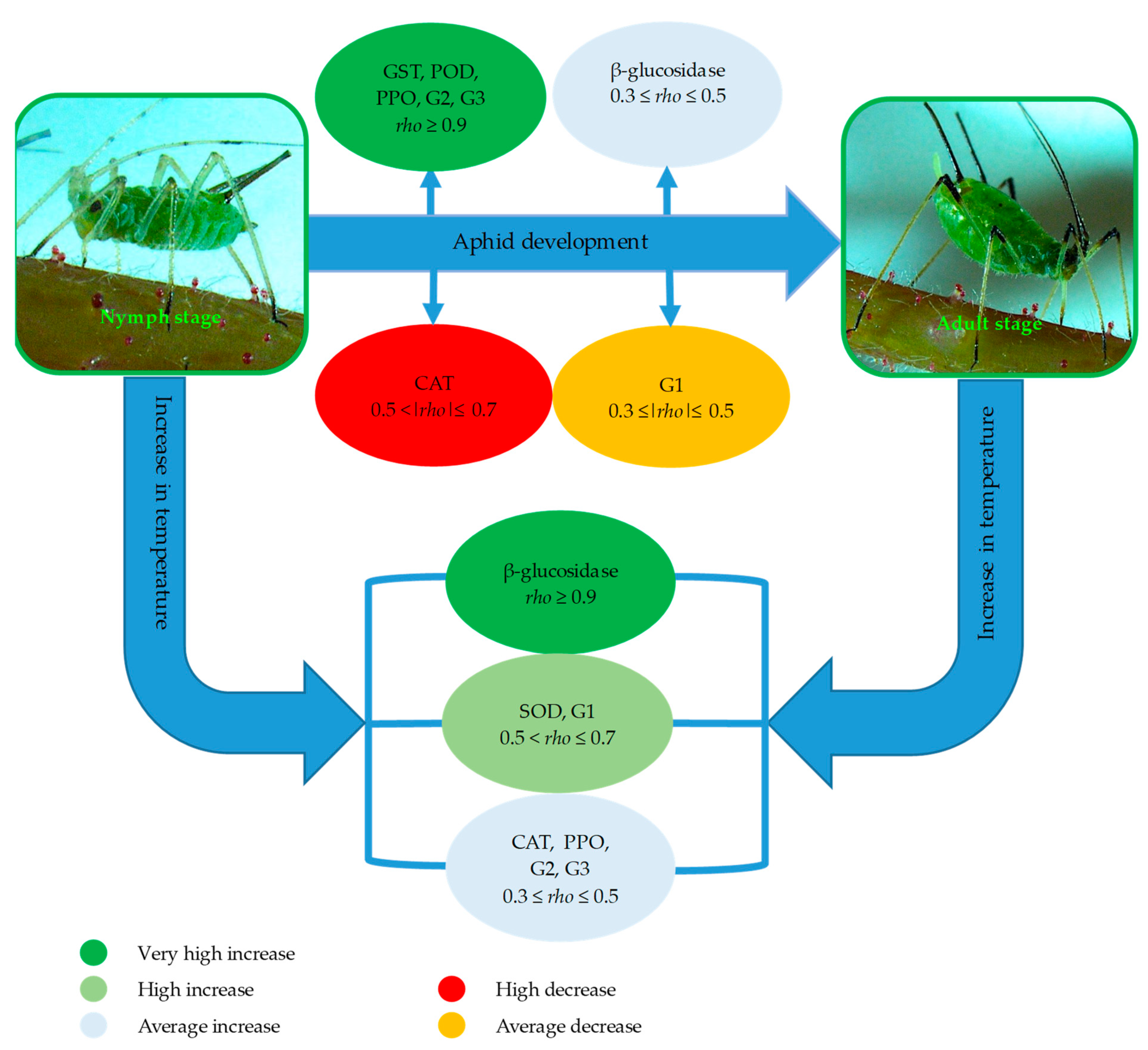
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bale, J.S.; Gerday, C.; Parker, A.; Marahiel, M.A.; Shanks, I.A.; Davies, P.L.; Warren, G. Insects and low temperatures: From molecular biology to distributions and abundance. Philos. Trans. R. Soc. B Biol. Sci. 2002, 357, 849–862. [Google Scholar] [CrossRef] [Green Version]
- Hullé, M.; Coeur d’Acier, A.; Bankhead-Dronnet, S.; Harrington, R. Aphids in the face of global changes. Comptes Rendus Biol. 2010, 333, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Rukke, B.A.; Sivasubramaniam, R.; Birkemoe, T.; Aak, A. Temperature stress deteriorates bed bug (Cimex lectularius) populations through decreased survival, fecundity and offspring success. PLoS ONE 2018, 13, e0193788. [Google Scholar] [CrossRef] [Green Version]
- Harrington, R.; Fleming, R.A.; Woiwod, I.P. Climate change impacts on insect management and conservation in temperate regions: Can they be predicted? Agric. For. Entomol. 2001, 3, 233–240. [Google Scholar] [CrossRef]
- Wu, Y.; Li, J.; Liu, H.; Qiao, G.; Huang, X. Investigating the impact of climate warming on phenology of aphid pests in China using long-term historical data. Insects 2020, 11, 167. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Fu, W.; Li, N.; Zhang, F.; Liu, T.X. Antioxidant responses of Propylaea japonica (Coleoptera: Coccinellidae) exposed to high temperature stress. J. Insect Physiol. 2015, 73, 47–52. [Google Scholar] [CrossRef]
- Jena, K.; Kumar Kar, P.; Kausar, Z.; Babu, C.S. Effects of temperature on modulation of oxidative stress and antioxidant defenses in testes of tropical tasar silkworm Antheraea mylitta. J. Therm. Biol. 2013, 38, 199–204. [Google Scholar] [CrossRef]
- Chen, H.; Solangi, G.S.; Guo, J.; Wan, F.; Zhou, Z. Antioxidant responses of ragweed leaf beetle Ophraella communa (Coleoptera: Chrysomelidae) exposed to thermal stress. Front. Physiol. 2018, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.H.; Liu, H.; Wang, J.J.; Wang, Z.Y. Effects of thermal stress on lipid peroxidation and antioxidant enzyme activities of the predatory mite, Neoseiulus cucumeris (Acari: Phytoseiidae). Exp. Appl. Acarol. 2014, 64. [Google Scholar] [CrossRef]
- Kang, Z.-W.; Liu, F.-H.; Liu, X.; Yu, W.-B.; Tan, X.-L.; Zhang, S.-Z.; Tian, H.-G.; Liu, T.-X. The potential coordination of the heat-shock proteins and antioxidant enzyme genes of Aphidius gifuensis in response to thermal stress. Front. Physiol. 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Rashid, M.A.; Huang, Q.Y.; Wong, C.; Lei, C.-L. Response of antioxidant enzymes in Mythimna separata (Lepidoptera: Noctuidae) exposed to thermal stress. Bull. Entomol. Res. 2017, 107, 382–390. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: New York, NY, USA, 2015; ISBN 9780198717478. [Google Scholar]
- Barbehenn, R.V. Gut-based antioxidant enzymes in a polyphagous and a graminivorous grasshopper. J. Chem. Ecol. 2002, 28, 1329–1347. [Google Scholar] [CrossRef]
- Wang, Y.; Oberley, L.W.; Murhammer, D.W. Antioxidant defense systems of two lipidopteran insect cell lines. Free Radic. Biol. Med. 2001, 30, 1254–1262. [Google Scholar] [CrossRef]
- Krishnan, N.; Sehnal, F. Compartmentalization of oxidative stress and antioxidant defense in the larval gut of Spodoptera littoralis. Arch. Insect Biochem. Physiol. 2006, 63, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, C.C.; Koenig, C.; Araya, C.; Santos, M.J.; Niemeyer, H.M. Effect of DIMBOA, a hydroxamic acid from cereals, on peroxisomal and mitochondrial enzymes from aphids: Evidence for the presence of peroxisomes in aphids. J. Chem. Ecol. 1999, 25, 2465–2475. [Google Scholar] [CrossRef]
- Loayza-Muro, R.; Figueroa, C.C.; Niemeyer, H.M. Effect of two wheat cultivars differing in hydroxamic acid concentration on detoxification metabolism in the aphid Sitobion avenae. J. Chem. Ecol. 2000, 26, 2725–2736. [Google Scholar] [CrossRef]
- Lukasik, I. Changes in activity of superoxide dismutase and catalase within cereal aphids in response to plant o-dihydroxyphenols. J. Appl. Entomol. 2007, 131, 209–214. [Google Scholar] [CrossRef]
- Łukasik, I.; Goławska, S.; Wójcicka, A.; Goławski, A. Effect of host plants on antioxidant system of pea aphid Acyrthosiphon pisum. Bull. Insectol. 2011, 64, 153–158. [Google Scholar]
- Łukasik, I.; Goławska, S. Effect of host plant on levels of reactive oxygen species and antioxidants in the cereal aphids Sitobion avenae and Rhopalosiphum padi. Biochem. Syst. Ecol. 2013, 51, 232–239. [Google Scholar] [CrossRef]
- Bi, J.L.; Felton, G.W. Foliar oxidative stress and insect herbivory: Primary compounds, secondary metabolites, and reactive oxygen species as components of induced resistance. J. Chem. Ecol. 1995, 21, 1511–1530. [Google Scholar] [CrossRef] [PubMed]
- Łukasik, I. Effect of host plant alternation on some adaptive enzymes of the bird cherry–oat aphid, Rhopalosiphum padi (L.). J. Pest Sci. 2009, 82, 203–209. [Google Scholar] [CrossRef]
- Dmochowska-Ślęzak, K.; Giejdasz, K.; Fliszkiewicz, M.; Żółtowska, K. Variations in antioxidant defense during the development of the solitary bee Osmia bicornis. Apidologie 2015, 46, 432–444. [Google Scholar] [CrossRef]
- Krishnan, N.; Kodrík, D.; Turanli, F.; Sehnal, F. Stage-specific distribution of oxidative radicals and antioxidant enzymes in the midgut of Leptinotarsa decemlineata. J. Insect Physiol. 2007, 53, 67–74. [Google Scholar] [CrossRef]
- Urbańska, A. Occurrence and source of hydrogen peroxide in aphids. Electron. J. Pol. Agric. Univ. 2009, 12, 4. [Google Scholar]
- Cai, Q.-N.; Han, Y.; Cao, Y.-Z.; Hu, Y.; Zhao, X.; Bi, J.-L. Detoxification of gramine by the cereal aphid Sitobion avenae. J. Chem. Ecol. 2009, 35, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Board, P.G.; Menon, D. Glutathione transferases, regulators of cellular metabolism and physiology. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3267–3288. [Google Scholar] [CrossRef]
- Mehrabadi, M.; Bandani, A.R.; Kwon, O. Biochemical characterization of digestive α-d-glucosidase and β-d-glucosidase from labial glands and midgut of wheat bug Eurygaster maura (Hemiptera: Scutelleridae). Entomol. Res. 2011, 41, 81–87. [Google Scholar] [CrossRef]
- Dancewicz, K.; Paprocka, M.; Morkunas, I.; Gabryś, B. Struggle to survive: Aphid—plant relationships under low-light stress. A case of Acyrthosiphon pisum (Harris) and Pisum sativum L. Arthropod. Plant Interact. 2018, 12, 97–111. [Google Scholar] [CrossRef] [Green Version]
- Urbanska, A.; Freddy Tjallingii, W.; Dixon, A.F.G.; Leszczynski, B. Phenol oxidising enzymes in the grain aphid’s saliva. Entomol. Exp. Appl. 1998, 86, 197–203. [Google Scholar] [CrossRef]
- Conti, B. Considerations on temperature, longevity and aging. Cell. Mol. Life Sci. 2008, 65, 1626–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mołoń, M.; Dampc, J.; Kula-Maximenko, M.; Zebrowski, J.; Mołoń, A.; Dobler, R.; Durak, R.; Skoczowski, A. Effects of temperature on lifespan of Drosophila melanogaster from different genetic backgrounds: Links between metabolic rate and longevity. Insects 2020, 11, 470. [Google Scholar] [CrossRef]
- Dampc, J.; Kula-Maximenko, M.; Molon, M.; Durak, R. Enzymatic defense response of apple aphid Aphis pomi to increased temperature. Insects 2020, 11, 436. [Google Scholar] [CrossRef] [PubMed]
- Blackman, R.L.; Eastop, V.F. Aphids of the World’s Plants: An Online Identification and Information Guide. Available online: http://www.aphid sonworldsplants.info (accessed on 26 May 2021).
- Gupta, R.; Tara, J.S. Life history of Aphis pomi De Geer (green apple aphid) on apple plantations in Jammu Province, J&K, India. Munis Entomol. Zool. 2015, 10, 2–6. [Google Scholar]
- Mehrparvar, M.; Mansouri, S.M.; Hatami, B. Some bioecological aspects of the rose aphid, Macrosiphum rosae (Hemiptera: Aphididae) and its natural enemies. Acta Univ. Sapientiae Agric. Environ. 2016, 8, 74–88. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, K.; Świątek, P.; Durak, R. Influence of selected biogenic amines on development and demographic parameters of a temperate population of Cinara (Cupressobium) cupressi (Hemiptera, Aphididae). Arthropod. Plant Interact. 2021, 1–11. [Google Scholar] [CrossRef]
- Durak, R.; Borowiak-Sobkowiak, B.; Socha, M. Bionomy and ecology of Cinara cupressi (BUCKTON, 1881) (Hemiptera, Aphidoidea). Pol. J. Entomol. 2007, 2, 107–113. [Google Scholar]
- Alford, D.V. Pests of Ornamental Trees, Shrubs and Flowers: A Colour Handbook, 2nd ed.; Manson Publishing: London, UK, 2012. [Google Scholar]
- Bowler, K.; Terblanche, J.S. Insect thermal tolerance: What is the role of ontogeny, ageing and senescence? Biol. Rev. 2008, 83, 339–355. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Arthur Woods, H.; Buckley, L.B.; Potter, K.A.; MacLean, H.J.; Higgins, J.K. Complex life cycles and the responses of insects to climate change. Integr. Comp. Biol. 2011, 51, 719–732. [Google Scholar] [CrossRef] [Green Version]
- Durak, R.; Molon, M.; Durak, T.; Chrzanowski, G. The enzymatic markers of the adaptation of Cinara tujafilina to changing the host plant. Ethol. Ecol. Evol. 2018, 30, 416–429. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Katagiri, C. α-D-Glucosidase in the serum of the american cockroach, Periplaneta americana. Insect Biochem. 1979, 9, 199–204. [Google Scholar] [CrossRef]
- Leszczynski, B.; Dixon, A.F.G. Resistance of cereals to aphids: The interaction between hydroxamic acids and glutathione S-transferases in the grain aphid Sitobion avenae (F.) (Hom., Aphididae). J. Appl. Entomol. 1992, 113, 61–67. [Google Scholar] [CrossRef]
- Miles, P.W. Studies on the salivary physiology of plant bugs: Oxidase activity in the salivary apparatus and saliva. J. Insect Physiol. 1964, 10, 121–129. [Google Scholar] [CrossRef]
- Laurema, S.; Varis, A.-L.; Miettinen, H. Studies on enzymes in the salivary glands of Lygus rugulipennis (Hemiptera, miridae). Insect Biochem. 1985, 15, 211–224. [Google Scholar] [CrossRef]
- Fehrmann, H.; Dimond, A.E. Peroxidase activity and Phytophthora resistance in different organs of the potato plant. Phytopathology 1969, 57, 69–72. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with Folin reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Enriquez, T.; Colinet, H. Basal tolerance to heat and cold exposure of the spotted wing drosophila, Drosophila suzukii. PeerJ 2017, 5, e3122. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.S.; Wang, L.; Zhang, W.; Rudolf, V.H.W. Resolving biological impacts of multiple heat waves: Interaction of hot and recovery days. Oikos 2018, 127, 622–633. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Bai, Q.; Zheng, X.; Lu, Z. Expression and enzyme activity of catalase in Chilo suppressalis (Lepidoptera: Crambidae) is responsive to environmental stresses. J. Econ. Entomol. 2017, 110, 1803–1812. [Google Scholar] [CrossRef]
- Farjan, M.; Dmitryjuk, M.; Lipiński, Z.; Biernat-Łopieńska, E.; Zółtowska, K. Supplementation of the honey bee diet with vitamin C: The effect on the antioxidative system of Apis mellifera carnica brood at different stages. J. Apic. Res. 2012, 51, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.B.; Roberts, S.P.; Elekonich, M.M. Age and natural metabolically-intensive behavior affect oxidative stress and antioxidant mechanisms. Exp. Gerontol. 2008, 43, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Molon, M.; Zadrag-Tecza, R. Effect of temperature on replicative aging of the budding yeast Saccharomyces cerevisiae. Biogerontology 2016, 17, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Van Voorhies, W.A.; Ward, S. Genetic and environmental conditions that increase longevity in Caenorhabditis elegans decrease metabolic rate. Proc. Natl. Acad. Sci. USA 1999, 96, 11399–11403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borowiak-Sobkowiak, B.; Woźniak, A.; Bednarski, W.; Formela, M.; Samardakiewicz, S.; Morkunas, I. Brachycorynella asparagi (Mordv.) Induced—Oxidative stress and antioxidative defenses of Asparagus officinalis L. Int. J. Mol. Sci. 2016, 17, 1740. [Google Scholar] [CrossRef] [Green Version]
- Durak, R.; Dampc, J.; Dampc, J. Role of temperature on the interaction between Japanese quince Chaenomeles japonica and herbivorous insect Aphis pomi (Hemiptera: Aphidoidea). Environ. Exp. Bot. 2020, 176, 104100. [Google Scholar] [CrossRef]
- Durak, R.; Jedryczka, M.; Czajka, B.; Dampc, J.; Wielgusz, K.; Borowiak-Sobkowiak, B. Mild abiotic stress affects development and stimulates hormesis of hemp aphid Phorodon cannabis. Insects 2021, 12, 420. [Google Scholar] [CrossRef]
- Durak, R.; Borowiak-Sobkowiak, B. Influence of temperature on the biological parameters of the anholocyclic species Cinara tujafilina (Hemiptera: Aphidoidea). Cent. Eur. J. Biol. 2013, 8, 570–577. [Google Scholar] [CrossRef]
- Zhao, F.; Hoffmann, A.A.; Xing, K.; Ma, C. Life stages of an aphid living under similar thermal conditions differ in thermal performance. J. Insect Physiol. 2017, 99, 1–7. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| SOD | CAT | GST | β-Glucosidase | POD | PPO | |
|---|---|---|---|---|---|---|
| Aphis pomi | ||||||
| T | F(2,72) = 4.99 *** | F(2,72) = 72.42 *** | F(2,72) = 20.61 *** | F(2,72) = 534.30 *** | F(2,72) = 31.95 *** | F(2,72) = 64.17 *** |
| t | F(5,72) = 9.96 *** | F(5,72) = 114.14 *** | F(5,72) = 6.42 *** | F(5,72) = 474.83 *** | F(5,72) = 26.70 *** | F(5,72) = 64.61 *** |
| m | F(1,72) = 2937.76 *** | F(1,72) = 677.21 *** | F(1,72) = 555,58 *** | F(1,72) = 14.82 *** | F(1,72) = 970.45 *** | F(1,72) = 13.02 *** |
| T × m | F(2,72) = 3.63 *** | F(2,72) = 32.87 *** | F(2,72) = 4.31 * | F(2,72) = 0.18 | F(2,72) = 2.01 | F(2,72) = 11.56 *** |
| T × t | F(10,72) = 1.72 | F(10,72) = 20.21 *** | F(10,72) = 1.85 | F(10,72) = 54.52 *** | F(10,72) = 3.58 *** | F(10,72) = 5.62 *** |
| t × m | F(5,72) = 2.87 ** | F(5,72) = 20.51 *** | F(5,72) = 2.83 * | F(5,72) = 4.33 *** | F(5,72) = 5.28 *** | F(5,72) = 14.41 *** |
| T × t × m | F(10,72) = 1.90 * | F(10,72) = 6.29 *** | F(10,72) = 1.07 | F(10,72) = 4.30 *** | F(10,72) = 1.59 | F(10,72) = 26.65 *** |
| Macrosiphum rosae | ||||||
| T | F(2,72) = 63.86 *** | F(2,72) = 2.33 ** | F(2,72) = 5.39 ** | F(2,72) = 7.66 *** | F(2,72) = 6.25 ** | F(2,72) = 15.98 *** |
| t | F(5,72) = 43.43 *** | F(5,72) = 32.83 *** | F(5,72) = 2.86 * | F(5,72) = 1.39 | F(5,72) = 19.10 *** | F(5,72) = 3.98 ** |
| m | F(1,72) = 976.55 *** | F(1,72) = 37.78 *** | F(1,72) = 7884.39 *** | F(1,72) = 20.58 *** | F(1,72) = 2351.88 *** | F(1,72) = 108.12*** |
| T × m | F(2,72) = 84.61 *** | F(2,72) = 0.43 ** | F(2,72) = 0.10 | F(2,72) = 1.17 | F(2,72) = 4.24 * | F(2,72) = 0.14 |
| T × t | F(10,72) = 15.83 *** | F(10,72) = 2.18 ** | F(10,72) = 2.71 ** | F(10,72) = 1.62 | F(10,72) = 2.21 * | F(10,72) = 2.43 * |
| t × m | F(5,72) = 34.01 *** | F(5,72) = 10.28 *** | F(5,72) = 2.09 | F(5,72) = 3.22 * | F(5,72) = 17.58 *** | F(5,72) = 3.55 ** |
| T × t × m | F(10,72) = 14.17 *** | F(10,72) = 3.81 *** | F(10,72) = 2.68 ** | F(10,72) = 1.27 | F(10,72) = 2.33 * | F(10,72) = 3.03 ** |
| Cinara cupressi | ||||||
| T | F(2,72) = 118.67 *** | F(2,72) = 335.74 *** | F(2,72) = 154.28 *** | F(2,72) = 13.74 *** | F(2,72) = 114.34 *** | F(2,72) = 60.89 *** |
| t | F(5,72) = 17.43 *** | F(5,72) = 529.52 *** | F(5,72) = 85.47 *** | F(5,72) = 6.00 *** | F(5,72) = 60.27 *** | F(5,72) = 12.05 *** |
| m | F(1,72) = 1729.37 *** | F(1,72) = 378.08 *** | F(1,72) = 8183.41 *** | F(1,72) = 2467.09 *** | F(1,72) = 2661.945 *** | F(1,72) = 2297.45 *** |
| T × m | F(2,72) = 24.44 *** | F(2,72) = 123.77 *** | F(2,72) = 21.18 *** | F(2,72) = 6.37 ** | F(2,72) = 124.26 *** | F(2,72) = 37.06 *** |
| T × t | F(10,72) = 7.57 *** | F(10,72) = 114.84 *** | F(10,72) = 9.08 *** | F(10,72) = 4.07 *** | F(10,72) = 15.27 *** | F(10,72) = 19.35 *** |
| t × m | F(5,72) = 7.67 *** | F(5,72) = 173.21 *** | F(5,72) = 48.55 *** | F(5,72) = 3.16 * | F(5,72) = 53.85 *** | F(5,72) = 17.96 *** |
| T × t × m | F(10,72) = 3.17 ** | F(10,72) = 53.65 *** | F(10,72) = 3.46 *** | F(10,72) = 1.72 | F(10,72) = 13.71 *** | F(10,72) = 11.26 *** |
| Test Score (F) | |
|---|---|
| PC1 | |
| Temperature (T) | 38.31 *** |
| Morph (m) | 4635.07 *** |
| T × m | 3.31 * |
| PC2 | |
| Temperature (T) | 96.06 *** |
| Morph (m) | 4.32 * |
| T × m | 0.85 |
| Spearman’s rho Correlation Coefficients | ||
|---|---|---|
| PC1 | PC2 | |
| Enzymes | ||
| SOD | −0.27 ** | 0.64 *** |
| CAT | −0.55 *** | 0.37 *** |
| β-glucosidase | 0.37 *** | 0.88 *** |
| GST | 0.88 *** | 0.24 * |
| POD | 0.90 *** | 0.12 |
| PPO | 0.95 *** | 0.30 ** |
| Groups of enzymes | ||
| G1 (antioxidant enzymes) | −0.37 *** | 0.65 *** |
| G2 (detoxifying enzymes) | 0.90 *** | 0.35 *** |
| G3 (oxidoreductive enzymes) | 0.97 *** | 0.30 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durak, R.; Dampc, J.; Kula-Maximenko, M.; Mołoń, M.; Durak, T. Changes in Antioxidative, Oxidoreductive and Detoxification Enzymes during Development of Aphids and Temperature Increase. Antioxidants 2021, 10, 1181. https://doi.org/10.3390/antiox10081181
Durak R, Dampc J, Kula-Maximenko M, Mołoń M, Durak T. Changes in Antioxidative, Oxidoreductive and Detoxification Enzymes during Development of Aphids and Temperature Increase. Antioxidants. 2021; 10(8):1181. https://doi.org/10.3390/antiox10081181
Chicago/Turabian StyleDurak, Roma, Jan Dampc, Monika Kula-Maximenko, Mateusz Mołoń, and Tomasz Durak. 2021. "Changes in Antioxidative, Oxidoreductive and Detoxification Enzymes during Development of Aphids and Temperature Increase" Antioxidants 10, no. 8: 1181. https://doi.org/10.3390/antiox10081181
APA StyleDurak, R., Dampc, J., Kula-Maximenko, M., Mołoń, M., & Durak, T. (2021). Changes in Antioxidative, Oxidoreductive and Detoxification Enzymes during Development of Aphids and Temperature Increase. Antioxidants, 10(8), 1181. https://doi.org/10.3390/antiox10081181