
Oxidative Stress and Antioxidant Pathway in Allergic Rhinitis



Abstract
:1. Introduction
2. Oxidative Stress Pathways
2.1. Overview of Oxidative Stress
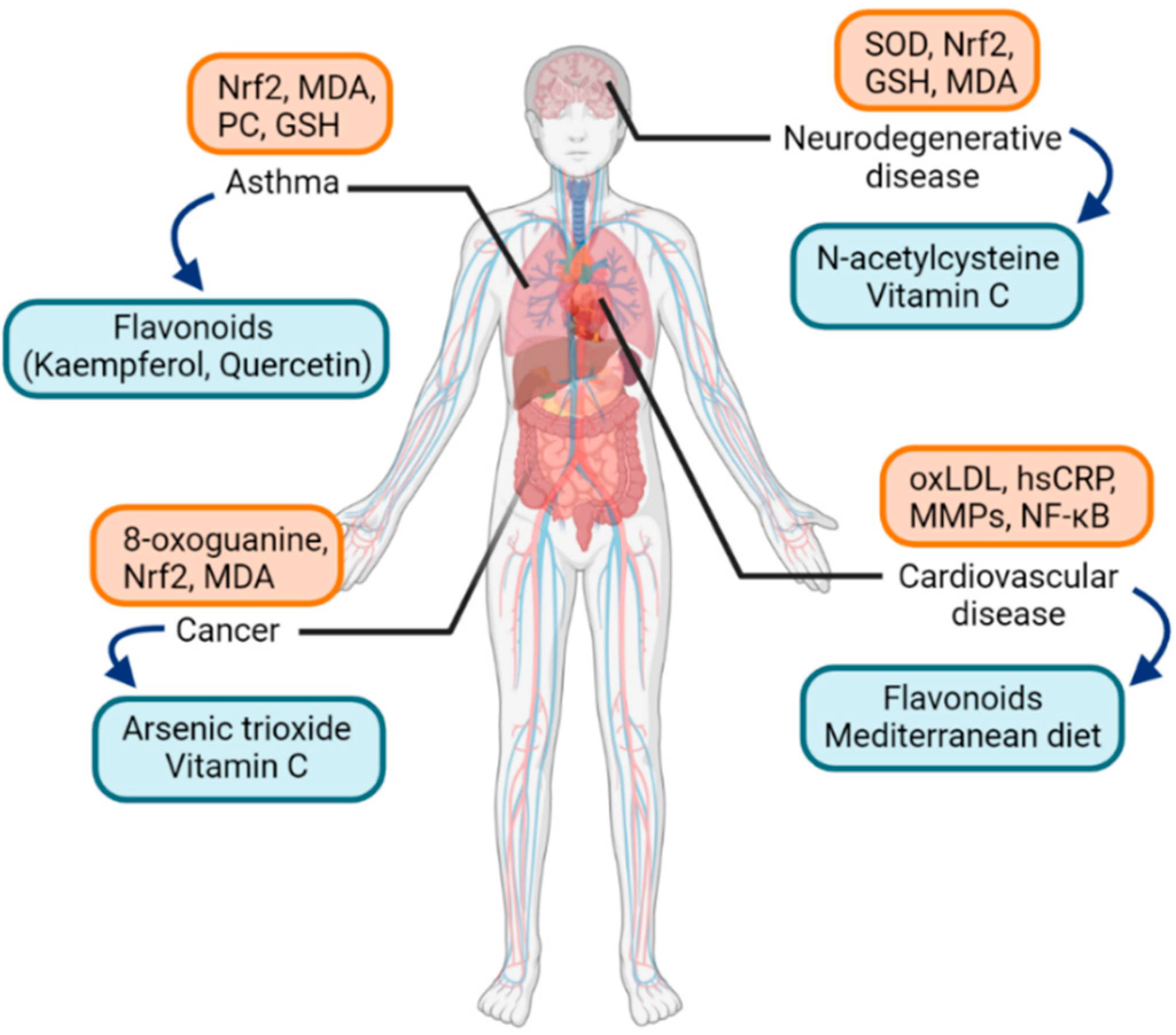
2.2. Oxidative Stress and Possible Therapeutic Antioxidants in Diseases
3. Allergic Rhinitis and Oxidative Stress
3.1. Pathophysiology of Allergic Rhinitis
3.2. Oxidative Stress and Allergic Rhinitis
3.3. Therapeutic Antioxidants in Allergic Rhinitis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical relevance of biomarkers of oxidative stress. Antioxid. Redox Signal. 2015, 23, 1144–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayer, S.B.; Maghzal, G.; Stocker, R.; Hampton, M.B.; Winterbourn, C.C. Neutrophil-mediated oxidation of erythrocyte peroxiredoxin 2 as a potential marker of oxidative stress in inflammation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2013, 27, 3315–3322. [Google Scholar]
- Samitas, K.; Carter, A.; Kariyawasam, H.H.; Xanthou, G. Upper and lower airway remodelling mechanisms in asthma, allergic rhinitis and chronic rhinosinusitis: The one airway concept revisited. Allergy 2018, 73, 993–1002. [Google Scholar] [CrossRef] [Green Version]
- Shamssain, M.H.; Shamsian, N. Prevalence and severity of asthma, rhinitis, and atopic eczema in 13- to 14-year-old schoolchildren from the northeast of england. Ann. Allergy Asthma Immunol. Off. Publ. Am. Coll. Allergy Asthma Immunol. 2001, 86, 428–432. [Google Scholar] [CrossRef]
- Bousquet, J.; Annesi-Maesano, I.; Carat, F.; Léger, D.; Rugina, M.; Pribil, C.; El Hasnaoui, A.; Chanal, I. Characteristics of intermittent and persistent allergic rhinitis: Dreams study group. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2005, 35, 728–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linneberg, A.; Nielsen, N.H.; Madsen, F.; Frølund, L.; Dirksen, A.; Jørgensen, T. Increasing prevalence of specific ige to aeroallergens in an adult population: Two cross-sectional surveys 8 years apart: The copenhagen allergy study. J. Allergy Clin. Immunol. 2000, 106, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, J.; Khaltaev, N.; Cruz, A.A.; Denburg, J.; Fokkens, W.J.; Togias, A.; Zuberbier, T.; Baena-Cagnani, C.E.; Canonica, G.W.; van Weel, C.; et al. Allergic rhinitis and its impact on asthma (aria) 2008 update (in collaboration with the world health organization, ga(2)len and allergen). Allergy 2008, 63 (Suppl. S86), 8–160. [Google Scholar] [CrossRef]
- Asher, M.I.; Montefort, S.; Björkstén, B.; Lai, C.K.; Strachan, D.P.; Weiland, S.K.; Williams, H. Worldwide time trends in the prevalence of symptoms of asthma, allergic rhinoconjunctivitis, and eczema in childhood: Isaac phases one and three repeat multicountry cross-sectional surveys. Lancet 2006, 368, 733–743. [Google Scholar] [CrossRef]
- Lee, S.L.; Wong, W.; Lau, Y.L. Increasing prevalence of allergic rhinitis but not asthma among children in hong kong from 1995 to 2001 (phase 3 international study of asthma and allergies in childhood). Pediatric Allergy Immunol. Off. Publ. Eur. Soc. Pediatric Allergy Immunol. 2004, 15, 72–78. [Google Scholar] [CrossRef]
- Reed, S.D.; Lee, T.A.; McCrory, D.C. The economic burden of allergic rhinitis: A critical evaluation of the literature. Pharmacoeconomics 2004, 22, 345–361. [Google Scholar] [CrossRef]
- Greiner, A.N.; Hellings, P.W.; Rotiroti, G.; Scadding, G.K. Allergic rhinitis. Lancet 2011, 378, 2112–2122. [Google Scholar] [CrossRef]
- Meltzer, E.O. Allergic rhinitis: Burden of illness, quality of life, comorbidities, and control. Immunol. Allergy Clin. North Am. 2016, 36, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Leynaert, B.; Neukirch, C.; Liard, R.; Bousquet, J.; Neukirch, F. Quality of life in allergic rhinitis and asthma. A population-based study of young adults. Am. J. Respir. Crit. Care Med. 2000, 162, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Meltzer, E.O.; Blaiss, M.S.; Derebery, M.J.; Mahr, T.A.; Gordon, B.R.; Sheth, K.K.; Simmons, A.L.; Wingertzahn, M.A.; Boyle, J.M. Burden of allergic rhinitis: Results from the pediatric allergies in america survey. J. Allergy Clin. Immunol. 2009, 124, S43–S70. [Google Scholar] [CrossRef]
- White, P.; Smith, H.; Baker, N.; Davis, W.; Frew, A. Symptom control in patients with hay fever in uk general practice: How well are we doing and is there a need for allergen immunotherapy? Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 1998, 28, 266–270. [Google Scholar] [CrossRef]
- Durham, S.R.; Penagos, M. Sublingual or subcutaneous immunotherapy for allergic rhinitis? J. Allergy Clin. Immunol. 2016, 137, 339–349.e10. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: Concept and some practical aspects. Antioxidants 2020, 9, 852. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.J. Positive oxidative stress in aging and aging-related disease tolerance. Redox Biol. 2014, 2, 165–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sies, H. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ros) homeostasis and redox regulation in cellular signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Church, D.F.; Pryor, W.A. Free-radical chemistry of cigarette smoke and its toxicological implications. Environ. Health Perspect. 1985, 64, 111–126. [Google Scholar] [CrossRef]
- Freudenthal, B.D.; Beard, W.A.; Perera, L.; Shock, D.D.; Kim, T.; Schlick, T.; Wilson, S.H. Uncovering the polymerase-induced cytotoxicity of an oxidized nucleotide. Nature 2015, 517, 635–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leon, J.; Sakumi, K.; Castillo, E.; Sheng, Z.; Oka, S.; Nakabeppu, Y. 8-oxoguanine accumulation in mitochondrial DNA causes mitochondrial dysfunction and impairs neuritogenesis in cultured adult mouse cortical neurons under oxidative conditions. Sci. Rep. 2016, 6, 22086. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, F.J.; Mudway, I.S. Protein oxidation at the air-lung interface. Amino Acids 2003, 25, 375–396. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R. Metal ion-catalyzed oxidation of proteins: Biochemical mechanism and biological consequences. Free Radic. Biol. Med. 1990, 9, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Ghezzi, P. Environmental risk factors and their footprints in vivo—A proposal for the classification of oxidative stress biomarkers. Redox Biol. 2020, 34, 101442. [Google Scholar] [CrossRef]
- Siu, G.M.; Draper, H.H. Metabolism of malonaldehyde in vivo and in vitro. Lipids 1982, 17, 349–355. [Google Scholar] [CrossRef]
- Karadogan, B.; Beyaz, S.; Gelincik, A.; Buyukozturk, S.; Arda, N. Evaluation of oxidative stress biomarkers and antioxidant parameters in allergic asthma patients with different level of asthma control. J. Asthma Off. J. Assoc. Care Asthma 2021. [Google Scholar] [CrossRef]
- Hoffmann, F.; Ender, F.; Schmudde, I.; Lewkowich, I.P.; Köhl, J.; König, P.; Laumonnier, Y. Origin, localization, and immunoregulatory properties of pulmonary phagocytes in allergic asthma. Front. Immunol. 2016, 7, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, V.; Banga, J.; Silveyra, P. Oxidative stress and cellular pathways of asthma and inflammation: Therapeutic strategies and pharmacological targets. Pharmacol. Ther. 2018, 181, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Mortaz, E.; Adcock, I.M.; Ito, K.; Kraneveld, A.D.; Nijkamp, F.P.; Folkerts, G. Cigarette smoke induces cxcl8 production by human neutrophils via activation of tlr9 receptor. Eur. Respir. J. 2010, 36, 1143–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosio, M.G.; Saetta, M.; Agusti, A. Immunologic aspects of chronic obstructive pulmonary disease. N. Engl. J. Med. 2009, 360, 2445–2454. [Google Scholar] [CrossRef]
- Li, Q.; Engelhardt, J.F. Interleukin-1beta induction of nfkappab is partially regulated by h2o2-mediated activation of nfkappab-inducing kinase. J. Biol. Chem. 2006, 281, 1495–1505. [Google Scholar] [CrossRef] [Green Version]
- Rahman, I.; MacNee, W. Role of transcription factors in inflammatory lung diseases. Thorax 1998, 53, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Mizumura, K.; Maruoka, S.; Shimizu, T.; Gon, Y. Role of nrf2 in the pathogenesis of respiratory diseases. Respir. Investig. 2020, 58, 28–35. [Google Scholar] [CrossRef]
- Shintani, Y.; Maruoka, S.; Gon, Y.; Koyama, D.; Yoshida, A.; Kozu, Y.; Kuroda, K.; Takeshita, I.; Tsuboi, E.; Soda, K.; et al. Nuclear factor erythroid 2-related factor 2 (nrf2) regulates airway epithelial barrier integrity. Allergol. Int. Off. J. Jpn. Soc. Allergol. 2015, 64, S54–S63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayers, I.; Hall, I.P. Pharmacogenetic approaches in the treatment of asthma. Curr. Allergy Asthma Rep. 2005, 5, 101–108. [Google Scholar] [CrossRef]
- Barnes, P.J. New anti-inflammatory targets for chronic obstructive pulmonary disease. Nat. Rev. Drug Discov. 2013, 12, 543–559. [Google Scholar] [CrossRef]
- Belvisi, M.G.; Hele, D.J.; Birrell, M.A. New anti-inflammatory therapies and targets for asthma and chronic obstructive pulmonary disease. Expert Opin. Targets 2004, 8, 265–285. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.H.; Shin, D.; Han, S.Y.; Park, S.H.; Kang, M.K.; Kim, J.L.; Kang, Y.H. Blockade of airway inflammation by kaempferol via disturbing tyk-stat signaling in airway epithelial cells and in asthmatic mice. Evid. Based Complementary Altern. Med. Ecam 2013, 2013, 250725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, S.; Pishnamazi, S.; Sadrzadeh, S.M.; Farid, F.; Farid, R.; Watson, R.R. Pycnogenol((r)) in the management of asthma. J. Med. Food 2001, 4, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.H.; Riesen, S.K.; Truong, K.P.; Lau, E.W.; Rohdewald, P.; Barreta, R.A. Pycnogenol as an adjunct in the management of childhood asthma. J. Asthma Off. J. Assoc. Care Asthma 2004, 41, 825–832. [Google Scholar] [CrossRef]
- Sbodio, J.I.; Snyder, S.H.; Paul, B.D. Redox mechanisms in neurodegeneration: From disease outcomes to therapeutic opportunities. Antioxid. Redox Signal. 2019, 30, 1450–1499. [Google Scholar] [CrossRef] [PubMed]
- Bradley-Whitman, M.A.; Timmons, M.D.; Beckett, T.L.; Murphy, M.P.; Lynn, B.C.; Lovell, M.A. Nucleic acid oxidation: An early feature of alzheimer’s disease. J. Neurochem. 2014, 128, 294–304. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, A.; Gu, F.; Essa, M.M.; Wegiel, J.; Kaur, K.; Brown, W.T.; Chauhan, V. Brain region-specific deficit in mitochondrial electron transport chain complexes in children with autism. J. Neurochem. 2011, 117, 209–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardan, A.Y.; Fung, L.K.; Libove, R.A.; Obukhanych, T.V.; Nair, S.; Herzenberg, L.A.; Frazier, T.W.; Tirouvanziam, R. A randomized controlled pilot trial of oral n-acetylcysteine in children with autism. Biol. Psychiatry 2012, 71, 956–961. [Google Scholar] [CrossRef] [Green Version]
- Dolske, M.C.; Spollen, J.; McKay, S.; Lancashire, E.; Tolbert, L. A preliminary trial of ascorbic acid as supplemental therapy for autism. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1993, 17, 765–774. [Google Scholar] [CrossRef]
- Jones, D.P.; Sies, H. The redox code. Antioxid. Redox Signal. 2015, 23, 734–746. [Google Scholar] [CrossRef] [Green Version]
- Kattoor, A.J.; Pothineni, N.V.K.; Palagiri, D.; Mehta, J.L. Oxidative stress in atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 42. [Google Scholar] [CrossRef]
- Senoner, T.; Dichtl, W. Oxidative stress in cardiovascular diseases: Still a therapeutic target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samadi, A.; Sabuncuoglu, S.; Samadi, M.; Isikhan, S.Y.; Chirumbolo, S.; Peana, M.; Lay, I.; Yalcinkaya, A.; Bjørklund, G. A comprehensive review on oxysterols and related diseases. Curr. Med. Chem. 2021, 28, 110–136. [Google Scholar] [CrossRef] [PubMed]
- Marchio, P.; Guerra-Ojeda, S.; Vila, J.M.; Aldasoro, M.; Victor, V.M.; Mauricio, M.D. Targeting early atherosclerosis: A focus on oxidative stress and inflammation. Oxidative Med. Cell. Longev. 2019, 2019, 8563845. [Google Scholar] [CrossRef]
- Chen, X.Q.; Hu, T.; Han, Y.; Huang, W.; Yuan, H.B.; Zhang, Y.T.; Du, Y.; Jiang, Y.W. Preventive effects of catechins on cardiovascular disease. Molecules 2016, 21, 1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCullough, M.L.; Peterson, J.J.; Patel, R.; Jacques, P.F.; Shah, R.; Dwyer, J.T. Flavonoid intake and cardiovascular disease mortality in a prospective cohort of us adults. Am. J. Clin. Nutr. 2012, 95, 454–464. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Hoffmann, G. Mediterranean dietary pattern, inflammation and endothelial function: A systematic review and meta-analysis of intervention trials. Nutr. Metab. Cardiovasc. Dis. NMCD 2014, 24, 929–939. [Google Scholar] [CrossRef]
- Ditano-Vázquez, P.; Torres-Peña, J.D.; Galeano-Valle, F.; Pérez-Caballero, A.I.; Demelo-Rodríguez, P.; Lopez-Miranda, J.; Katsiki, N.; Delgado-Lista, J.; Alvarez-Sala-Walther, L.A. The fluid aspect of the mediterranean diet in the prevention and management of cardiovascular disease and diabetes: The role of polyphenol content in moderate consumption of wine and olive oil. Nutrients 2019, 11, 2833. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative stress in cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Sosa, V.; Moliné, T.; Somoza, R.; Paciucci, R.; Kondoh, H.; ME, L.L. Oxidative stress and cancer: An overview. Ageing Res. Rev. 2013, 12, 376–390. [Google Scholar] [CrossRef]
- Rao, Y.; Li, R.; Zhang, D. A drug from poison: How the therapeutic effect of arsenic trioxide on acute promyelocytic leukemia was discovered. Sci. China. Life Sci. 2013, 56, 495–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reczek, C.R.; Chandel, N.S. Revisiting vitamin c and cancer. Science 2015, 350, 1317–1318. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, D.L.; Powis, G. Clinically evaluated cancer drugs inhibiting redox signaling. Antioxid. Redox Signal. 2017, 26, 262–273. [Google Scholar] [CrossRef] [Green Version]
- Sin, B.; Togias, A. Pathophysiology of allergic and nonallergic rhinitis. Proc. Am. Thorac. Soc. 2011, 8, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, L.M.; Togias, A. Clinical practice. Allergic rhinitis. N. Engl. J. Med. 2015, 372, 456–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Oord, R.A.; Sheikh, A. Filaggrin gene defects and risk of developing allergic sensitisation and allergic disorders: Systematic review and meta-analysis. BMJ 2009, 339, b2433. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.G.; Zhu, J.H.; Cheng, W.H.; Bao, Y.; Ho, Y.S.; Reddi, A.R.; Holmgren, A.; Arnér, E.S. Paradoxical roles of antioxidant enzymes: Basic mechanisms and health implications. Physiol. Rev. 2016, 96, 307–364. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, G.M.; Roman, M.G.; Flores, L.C.; Hubbard, G.B.; Salmon, A.B.; Zhang, Y.; Gelfond, J.; Ikeno, Y. The paradoxical role of thioredoxin on oxidative stress and aging. Arch. Biochem. Biophys. 2015, 576, 32–38. [Google Scholar] [CrossRef]
- Miller, D.M.; Buettner, G.R.; Aust, S.D. Transition metals as catalysts of “autoxidation” reactions. Free Radic. Biol. Med. 1990, 8, 95–108. [Google Scholar] [CrossRef]
- Henzler, T.; Steudle, E. Transport and metabolic degradation of hydrogen peroxide in chara corallina: Model calculations and measurements with the pressure probe suggest transport of H(2)O(2) across water channels. J. Exp. Bot. 2000, 51, 2053–2066. [Google Scholar] [CrossRef] [Green Version]
- Marinho, H.S.; Real, C.; Cyrne, L.; Soares, H.; Antunes, F. Hydrogen peroxide sensing, signaling and regulation of transcription factors. Redox Biol. 2014, 2, 535–562. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I.; et al. An nrf2/small maf heterodimer mediates the induction of phase ii detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Cebula, M.; Schmidt, E.E.; Arnér, E.S. Trxr1 as a potent regulator of the nrf2-keap1 response system. Antioxid. Redox Signal. 2015, 23, 823–853. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Tang, R.; Ba, G.; Li, M.; Lin, H. Anti-allergic and anti-inflammatory effects of resveratrol via inhibiting txnip-oxidative stress pathway in a mouse model of allergic rhinitis. World Allergy Organ. J. 2020, 13, 100473. [Google Scholar] [CrossRef] [PubMed]
- Schreck, R.; Rieber, P.; Baeuerle, P.A. Reactive oxygen intermediates as apparently widely used messengers in the activation of the nf-kappa b transcription factor and hiv-1. EMBO J. 1991, 10, 2247–2258. [Google Scholar] [CrossRef]
- Perkins, N.D. Integrating cell-signalling pathways with nf-kappab and ikk function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef]
- Pande, V.; Ramos, M.J. Molecular recognition of 15-deoxy-delta(12,14)-prostaglandin j2 by nuclear factor-kappa b and other cellular proteins. Bioorganic Med. Chem. Lett. 2005, 15, 4057–4063. [Google Scholar] [CrossRef] [PubMed]
- Seyedsadjadi, N.; Grant, R. The potential benefit of monitoring oxidative stress and inflammation in the prevention of non-communicable diseases (ncds). Antioxidants 2020, 10, 15. [Google Scholar] [CrossRef]
- Piao, C.H.; Fan, Y.J.; Nguyen, T.V.; Song, C.H.; Chai, O.H. Mangiferin alleviates ovalbumin-induced allergic rhinitis via nrf2/ho-1/nf-κb signaling pathways. Int. J. Mol. Sci. 2020, 21, 3415. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cui, C.; Sun, H. Anti-inflammatory effect of mangiferin on an experimental model of allergic rhinitis through the inhibition of nf-κb signaling pathways. J. Environ. Pathol. Toxicol. Oncol. Off. Organ Int. Soc. Environ. Toxicol. Cancer 2020, 39, 357–364. [Google Scholar] [CrossRef]
- Mattila, P.; Joenväärä, S.; Renkonen, J.; Toppila-Salmi, S.; Renkonen, R. Allergy as an epithelial barrier disease. Clin. Transl. Allergy 2011, 1, 5. [Google Scholar] [CrossRef] [Green Version]
- Van Gerven, L.; Boeckxstaens, G.; Hellings, P. Up-date on neuro-immune mechanisms involved in allergic and non-allergic rhinitis. Rhinology 2012, 50, 227–235. [Google Scholar] [CrossRef]
- Siti Sarah, C.O.; Md Shukri, N.; Mohd Ashari, N.S.; Wong, K.K. Zonula occludens and nasal epithelial barrier integrity in allergic rhinitis. PeerJ 2020, 8, e9834. [Google Scholar] [CrossRef] [PubMed]
- Steelant, B.; Farré, R.; Wawrzyniak, P.; Belmans, J.; Dekimpe, E.; Vanheel, H.; Van Gerven, L.; Kortekaas Krohn, I.; Bullens, D.M.A.; Ceuppens, J.L.; et al. Impaired barrier function in patients with house dust mite-induced allergic rhinitis is accompanied by decreased occludin and zonula occludens-1 expression. J. Allergy Clin. Immunol. 2016, 137, 1043–1053.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beutel, O.; Maraspini, R.; Pombo-García, K.; Martin-Lemaitre, C.; Honigmann, A. Phase separation of zonula occludens proteins drives formation of tight junctions. Cell 2019, 179, 923–936.e11. [Google Scholar] [CrossRef]
- London, N.R., Jr.; Tharakan, A.; Lane, A.P.; Biswal, S.; Ramanathan, M., Jr. Nuclear erythroid 2-related factor 2 activation inhibits house dust mite-induced sinonasal epithelial cell barrier dysfunction. Int. Forum Allergy Rhinol. 2017, 7, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.T.; Fan, Y.; Piao, C.H.; Nguyen, T.V.; Shin, D.U.; Jung, S.Y.; Hyeon, E.; Song, C.H.; Lee, S.Y.; Shin, H.S.; et al. Piper nigrum extract improves ova-induced nasal epithelial barrier dysfunction via activating nrf2/ho-1 signaling. Cell. Immunol. 2020, 351, 104035. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, P.J. Oxidative stress-based therapeutics in copd. Redox Biol. 2020, 33, 101544. [Google Scholar] [CrossRef] [PubMed]
- Fão, L.; Mota, S.I.; Rego, A.C. Shaping the nrf2-are-related pathways in alzheimer’s and parkinson’s diseases. Ageing Res. Rev. 2019, 54, 100942. [Google Scholar] [CrossRef] [PubMed]
- Yusin, J.; Wang, V.; Henning, S.M.; Yang, J.; Tseng, C.H.; Thames, G.; Arnold, I.; Heber, D.; Lee, R.P.; Sanavio, L.; et al. The effect of broccoli sprout extract on seasonal grass pollen-induced allergic rhinitis. Nutrients 2021, 13, 1337. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Zhang, Y.; Shen, L. Preliminary clinical effect evaluation of resveratrol in adults with allergic rhinitis. Int. Arch. Allergy Immunol. 2018, 175, 231–236. [Google Scholar] [CrossRef]
- Bui, T.T.; Piao, C.H.; Hyeon, E.; Fan, Y.; Choi, D.W.; Jung, S.Y.; Jang, B.H.; Shin, H.S.; Song, C.H.; Chai, O.H. Preventive effect of bupleurum chinense on nasal inflammation via suppressing t helper type 2, eosinophil and mast cell activation. Am. J. Chin. Med. 2019, 47, 405–421. [Google Scholar] [CrossRef]
- Edo, Y.; Otaki, A.; Asano, K. Quercetin enhances the thioredoxin production of nasal epithelial cells in vitro and in vivo. Medicines 2018, 5, 124. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Lu, Y.; Li, F.; Wu, W.; Xie, D.; Feng, Y. In vitro and in vivo antiallergic effects of taurine on allergic rhinitis. Int. Arch. Allergy Immunol. 2020, 181, 404–416. [Google Scholar] [CrossRef]
- Miraglia Del Giudice, M.; Maiello, N.; Capristo, C.; Alterio, E.; Capasso, M.; Perrone, L.; Ciprandi, G. Resveratrol plus carboxymethyl-β-glucan reduces nasal symptoms in children with pollen-induced allergic rhinitis. Curr. Med Res. Opin. 2014, 30, 1931–1935. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Exogenous Antioxidants | Diet Source | Authors (Year) | Study Designs | Therapeutic Antioxidant Effects |
|---|---|---|---|---|
| Sulforaphane | Broccoli, cabbage | Yusin, J. et al. (2021) [94] | Clinical trial (double-blind, randomized, placebo-controlled) | Clinical measurements (TNSS, PNIF) of AR patients improved after sulforaphane supplementation. In nasal mucus fluid, T2 cytokines such as IL-4, IL-5, and IL-13 were decreased, but there was no statistical significance. |
| London, N.R., Jr. et al. (2017) [89] | Human study (tissue-specific) | Human SNECs were harvested and stimulated with HDM with/without Nrf2 activation with sulforaphane. Epithelial cell junction protein ZO-1 was disrupted with HDM stimulation but increased when treated with sulforaphane before stimulation with HDM. Similar beneficial effect was found with transepithelial electrical resistance. | ||
| Resveratrol | Grapes, berries, peanuts | Zhang, W. et al. (2020) [77] | Animal study (OVA-induced murine AR model) | After resveratrol treatment, TXNIP, MDA, SOD, inflammatory cytokines, eosinophil numbers, and nasal symptoms were significantly altered compared to untreated AR mice. |
| Lv, C. et al. (2018) [95] | Clinical trial (double-blind, randomized, placebo-controlled) | AR patients treated with resveratrol showed reduction in nasal symptoms, serum IgE, IL-4, TNF-α, and eosinophil levels. | ||
| Mangiferin | Mango | Piao, C.H. et al. (2020) [82] | Animal study (OVA-induced murine AR model) | Mangiferin treatment led to reduction of nasal symptoms, nasal mucosa inflammation, inflammatory cell infiltration, and epithelial disruption in histopathology. In NALF, MDA level reduced, SOD activity increased, and Nrf2/HO-1 expression was upregulated, while expression of NF-κB was decreased. |
| Wang, Y. et al. (2020) [83] | Animal study (OVA-induced murine AR model) | After administration of mangiferin, MDA level was decreased, and NF-κB pathway was prevented, which led to downregulation of TNF-α and IL-1β. In histopathology, ciliary loss and eosinophil infiltration were decreased. | ||
| Piper nigrum extract | Black pepper | Bui, T.T. et al. (2020) [90] | Animal study (OVA-induced murine AR model) | After Piper nigrum extract treatment, mast cells histamine release, nasal symptoms in early phase reaction, and eosinophil accumulation in nasal lavage fluid and nasal tissue were decreased. |
| Quercetin | Onions, red wine, tea | Edo, Y. et al. (2018) [97] | Human study (tissue-specific) Animal study (OVA-induced murine AR model) | Human SNECs showed increased TRX production in ELISA when treated with quercetin. In animal model, quercetin was orally administered, and the nasal symptoms were inhibited. In NALF, TRX levels were increased. |
| Taurine | Scallops, tuna, octopus | Zhou, J. et al. (2020) [98] | Human study (serum marker) Animal study (OVA-induced murine AR model) | In AR patients compared to healthy controls, after treatment of taurine, serum SOD3 level was decreased. In animal model, AR symptoms, inflammatory cytokines (TNF- α, IL-4, and IL-6), and eosinophil and mast cell infiltration in nasal mucosa were decreased. SOD3 production was increased. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, M.; Lee, D.; Lee, S.H.; Kim, T.H. Oxidative Stress and Antioxidant Pathway in Allergic Rhinitis. Antioxidants 2021, 10, 1266. https://doi.org/10.3390/antiox10081266
Han M, Lee D, Lee SH, Kim TH. Oxidative Stress and Antioxidant Pathway in Allergic Rhinitis. Antioxidants. 2021; 10(8):1266. https://doi.org/10.3390/antiox10081266
Chicago/Turabian StyleHan, Munsoo, Dabin Lee, Sang Hag Lee, and Tae Hoon Kim. 2021. "Oxidative Stress and Antioxidant Pathway in Allergic Rhinitis" Antioxidants 10, no. 8: 1266. https://doi.org/10.3390/antiox10081266
APA StyleHan, M., Lee, D., Lee, S. H., & Kim, T. H. (2021). Oxidative Stress and Antioxidant Pathway in Allergic Rhinitis. Antioxidants, 10(8), 1266. https://doi.org/10.3390/antiox10081266