
Silicon Improves the Redox Homeostasis to Alleviate Glyphosate Toxicity in Tomato Plants—Are Nanomaterials Relevant?

 ,
,  ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Artificial Substrate
2.2. Plant Material and Growth Conditions
2.3. Experimental Design
- CTL—control plants grown in OECD substrate (negative control);
- Si—plants grown in OECD substrate and treated once a week with 1 mM Si by foliar spraying;
- Nano-SiO2—plants grown in OECD substrate and treated once a week with 1 mM nano-SiO2 by foliar spraying;
- GLY—plants grown in OECD substrate contaminated by 10 mg kg−1 GLY (positive control);
- GLY + Si—plants grown in OECD substrate contaminated by 10 mg kg−1 GLY and treated once a week with 1 mM Si by foliar spraying;
- GLY + nano-SiO2—plants grown in OECD substrate contaminated by 10 mg kg−1 GLY and treated once a week with 1 mM nano-SiO2 by foliar spraying
2.4. Biometric Determinations
2.5. Assessment of Lipid Peroxidation (LP)
2.6. Determination of ROS Levels—Superoxide Anion (O2•−) and Hydrogen Peroxide (H2O2)
2.7. Quantification of Non-Enzymatic Antioxidants—Proline (Pro), Glutathione (GSH) and Ascorbate (AsA)
2.8. Extraction and Quantification of Antioxidant Enzymes—Superoxide Dismutase (SOD; EC 1.15.1.1), Catalase (CAT; EC 1.11.1.6), Ascorbate Peroxidase (APX; EC 1.11.1.11), Glutathione Reductase (GR; EC 1.8.1.7), and Dehydroascorbate Reductase (DHAR; EC 1.8.5.1)
2.9. Quantification of GLY and Aminomethylphosphonic Acid (AMPA) Accumulation in Plant Tissues
2.10. Statistical Analyses
3. Results
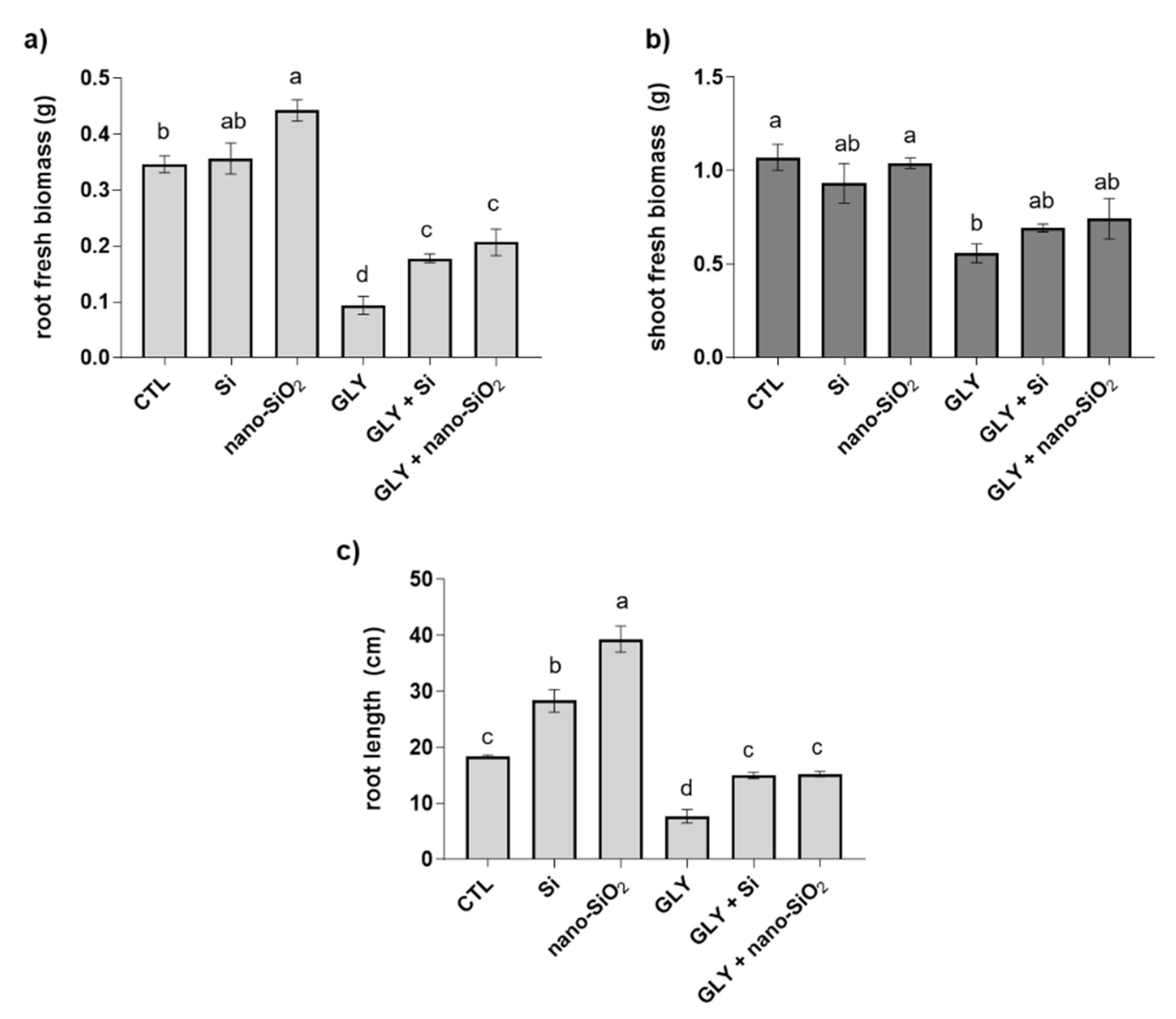
3.1. Biometric and Growth-Related Parameters
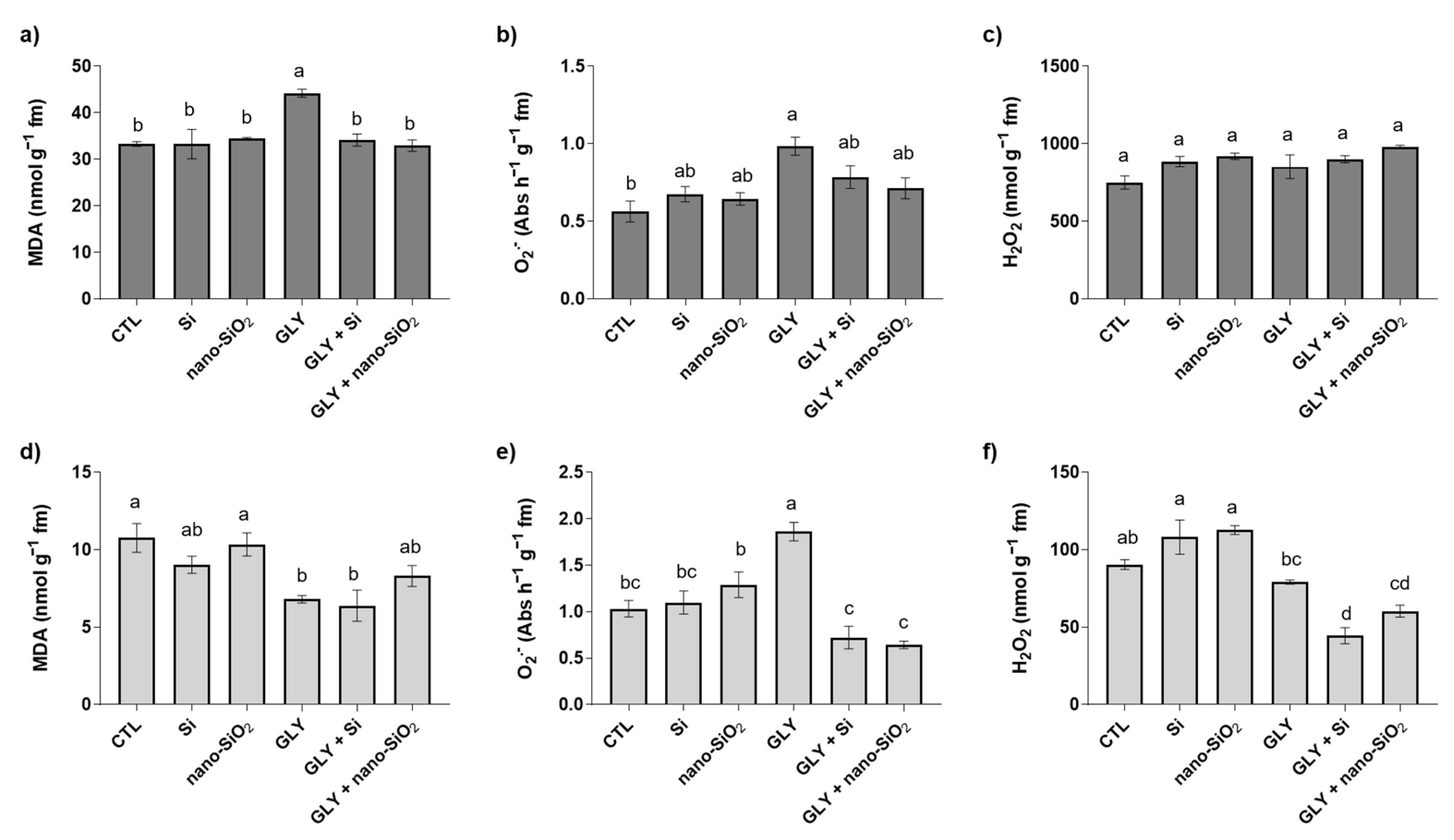
3.2. Lipid Peroxidation (LP)—MDA content
3.3. ROS Homeostasis—O2•− and H2O2 Content
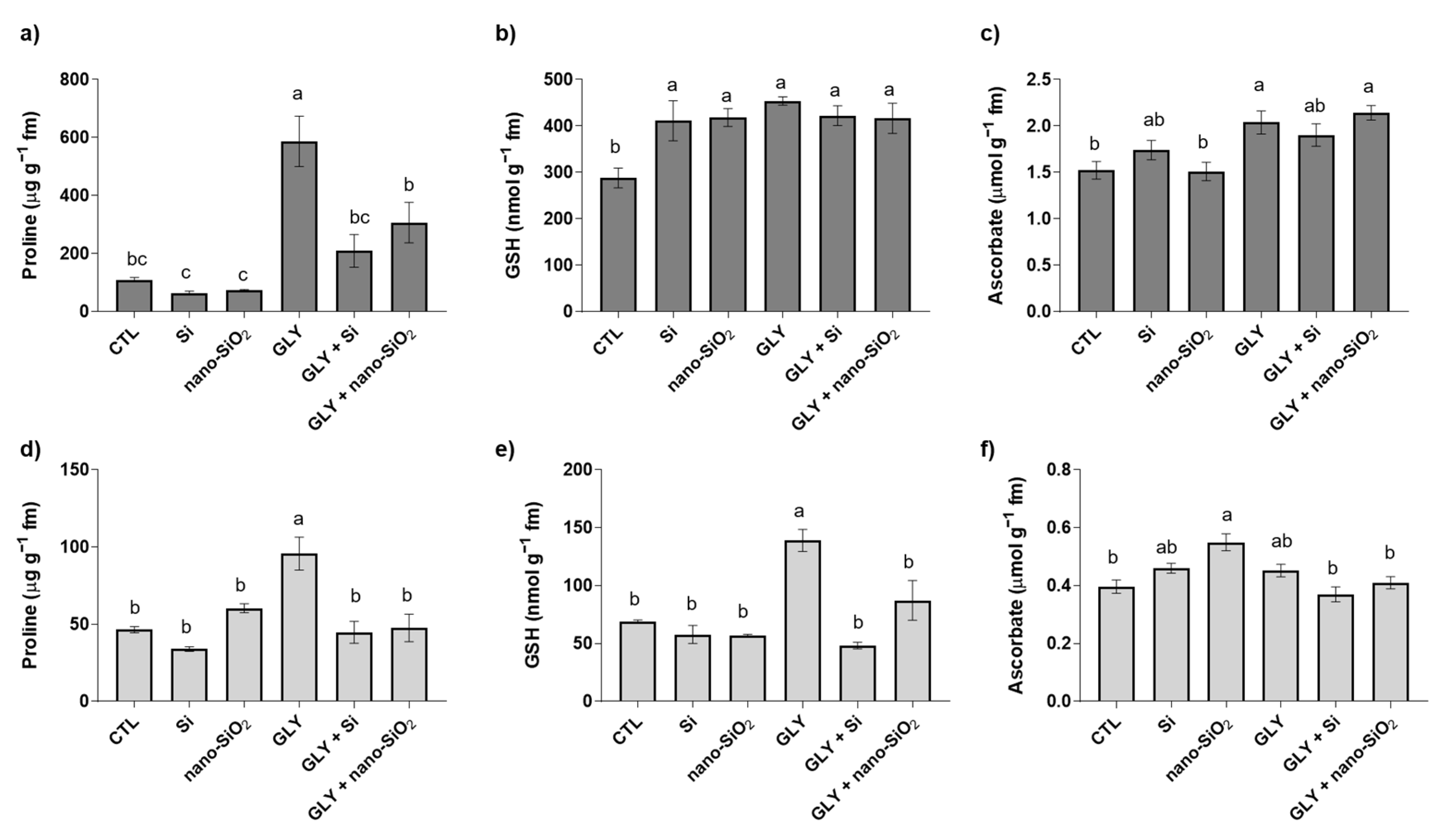
3.4. Non-Enzymatic Antioxidants—Pro, GSH and AsA
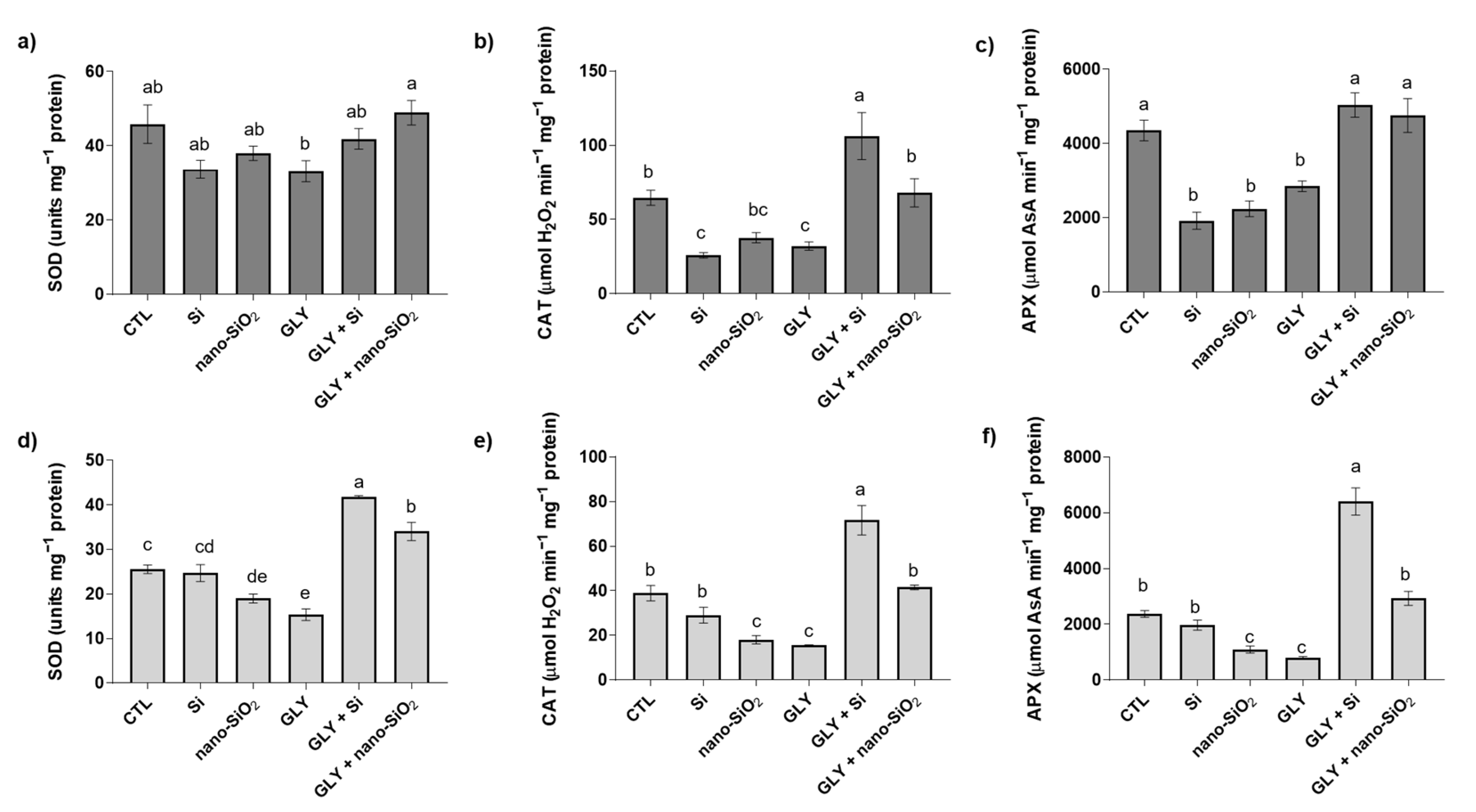
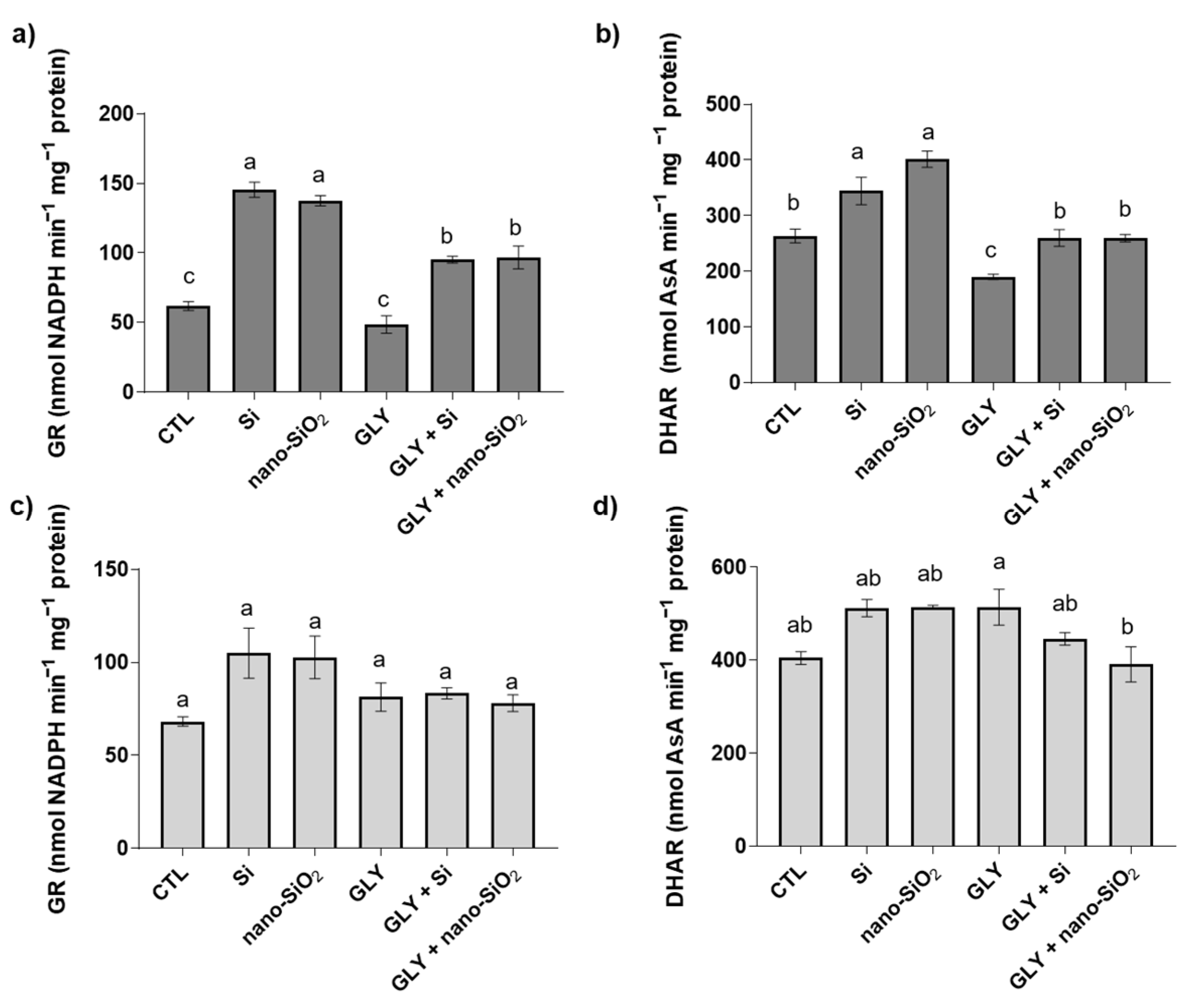
3.5. Enzymatic Antioxidants—Activity of SOD, CAT, APX, GR, and DHAR
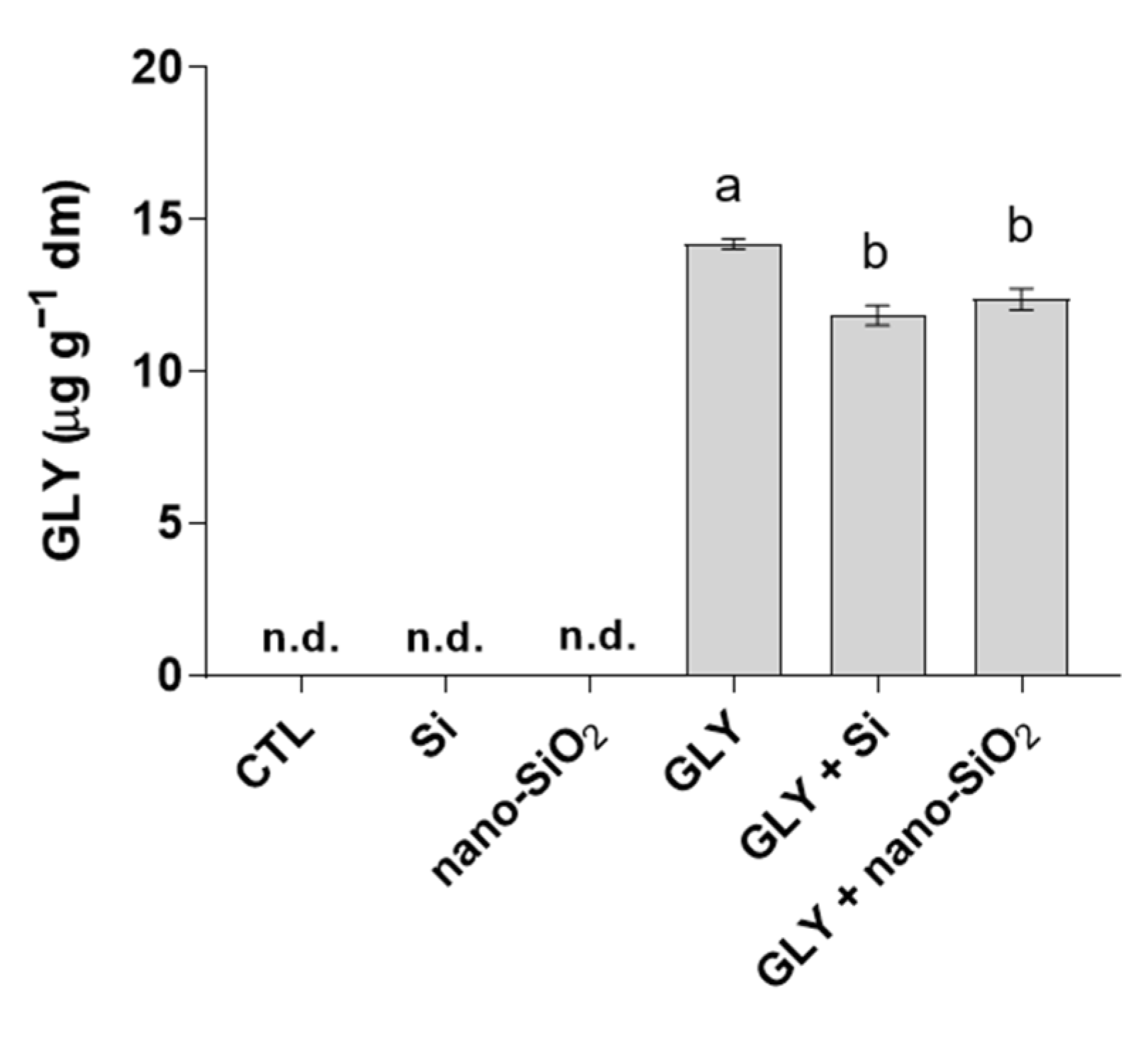
3.6. Bioaccumulation of GLY in Shoots and Roots
4. Discussions
4.1. GLY-Mediated Inhibition of Plant Growth Is Efficiently Counteracted by the Foliar Application of Si or Nano-SiO2
4.2. The Foliar Application of Si or Nano-SiO2 Reduces GLY-Induced Oxidative Stress, Particularly Stimulating the Enzymes of the Antioxidant Defense System
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The Future of Food and Agriculture—Trends and Challenges; FAO: Rome, Italy, 2017. [Google Scholar]
- Nishimoto, R. Global trends in the crop protection industry. J. Pestic. Sci. 2019, 44, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Mandal, A.; Sarkar, B.; Mandal, S.; Vithanage, M.; Patra, A.K.; Manna, M.C. Impact of agrochemicals on soil health. In Agrochemicals Detection, Treatment and Remediation; Elsevier: Amsterdam, The Netherlands, 2020; pp. 161–187. [Google Scholar]
- Atwood, D.; Paisley-Jones, C. Pesticides Industry Sales and Usage. 2008–2012 Market Estimates; U.S. Environmental Protection Agency (EPA): Washington, DC, USA, 2017.
- Van Bruggen, A.H.C.; He, M.M.; Shin, K.; Mai, V.; Jeong, K.C.; Finckh, M.R.; Morris, J.G. Environmental and health effects of the herbicide glyphosate. Sci. Total Environ. 2018, 616, 255–268. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Datta, S.; Wani, A.B.; Dhanjal, D.S.; Romero, R.; Singh, J. Glyphosate Uptake, Translocation, Resistance Emergence in Crops, Analytical Monitoring, Toxicity and Degradation: A Review; Springer International Publishing: Berlin/Heidelberg, Germany, 2020; Volume 18, ISBN 1031102000. [Google Scholar]
- Gomes, M.P.; Smedbol, E.; Chalifour, A.; Hénault-Ethier, L.; Labrecque, M.; Lepage, L.; Lucotte, M.; Juneau, P. Alteration of plant physiology by glyphosate and its by-product aminomethylphosphonic acid: An overview. J. Exp. Bot. 2014, 65, 4691–4703. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, K.M.; Weaver, L.M. The shikimate pathway. Annu. Rev. Plant Biol. 1999, 50, 473–503. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O. Glyphosate: Environmental fate and impact. Weed Sci. 2020, 68, 201–207. [Google Scholar] [CrossRef]
- Soares, C.; Pereira, R.; Spormann, S.; Fidalgo, F. Is soil contamination by a glyphosate commercial formulation truly harmless to non-target plants?—Evaluation of oxidative damage and antioxidant responses in tomato. Environ. Pollut. 2019, 247, 256–265. [Google Scholar] [CrossRef]
- Spormann, S.; Soares, C.; Fidalgo, F. Salicylic acid alleviates glyphosate-induced oxidative stress in Hordeum vulgare L. J. Environ. Manag. 2019, 241, 226–234. [Google Scholar] [CrossRef]
- Gomes, M.P.; Le Manac’h, S.G.; Hénault-Ethier, L.; Labrecque, M.; Lucotte, M.; Juneau, P. Glyphosate-Dependent Inhibition of Photosynthesis in Willow. Front. Plant Sci. 2017, 8, 207. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Singh, N.B.; Singh, A.; Hussain, I.; Yadav, V. Physiological and biochemical roles of nitric oxide against toxicity produced by glyphosate herbicide in Pisum sativum. Russ. J. Plant Physiol. 2017, 64, 518–524. [Google Scholar] [CrossRef]
- Epstein, E. Silicon: Its manifold roles in plants. Ann. Appl. Biol. 2009, 155, 155–160. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, W.; Zhu, Y.-G.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Epstein, E. Silicon. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Rahman, M.M.; Ansary, M.M.U.; Keya, S.S.; Abdelrahman, M.; Miah, M.G.; Phan Tran, L.S. Silicon in mitigation of abiotic stress-induced oxidative damage in plants. Crit. Rev. Biotechnol. 2021, 41, 918–934. [Google Scholar] [CrossRef] [PubMed]
- Guntzer, F.; Keller, C.; Meunier, J.D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Luyckx, M.; Hausman, J.F.; Lutts, S.; Guerriero, G. Silicon and plants: Current knowledge and technological perspectives. Front. Plant Sci. 2017, 8, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, A.; Tripathi, D.K.; Yadav, S.; Chauhan, D.K.; Živčák, M.; Ghorbanpour, M.; El-Sheery, N.I.; Brestic, M. Application of silicon nanoparticles in agriculture. 3 Biotech 2019, 9, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, C.; Branco-Neves, S.; de Sousa, A.; Azenha, M.; Cunha, A.; Pereira, R.; Fidalgo, F. SiO2 nanomaterial as a tool to improve Hordeum vulgare L. tolerance to nano-NiO stress. Sci. Total Environ. 2018, 622, 517–525. [Google Scholar] [CrossRef] [PubMed]
- OECD. Test No. 208: Terrestrial Plant Test: Seedling Emergence and Seedling Growth Test; OECD Publishing: Paris, France, 2006. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2015. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Khan, M.I.R.; Fujita, M. Silicon-mediated regulation of antioxidant defense and glyoxalase systems confers drought stress tolerance in Brassica napus L. S. Afr. J. Bot. 2018, 115, 50–57. [Google Scholar] [CrossRef]
- Wang, S.; Wang, F.; Gao, S. Foliar application with nano-silicon alleviates Cd toxicity in rice seedlings. Environ. Sci. Pollut. Res. 2015, 22, 2837–2845. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Torabian, S. Nano-silicon alters antioxidant activities of soybean seedlings under salt toxicity. Protoplasma 2018, 255, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Khaliq, A.; Ali, S.; Hameed, A.; Farooq, M.A.; Farid, M.; Shakoor, M.B.; Mahmood, K.; Ishaque, W.; Rizwan, M. Silicon alleviates nickel toxicity in cotton seedlings through enhancing growth, photosynthesis, and suppressing Ni uptake and oxidative stress. Arch. Agron. Soil Sci. 2016, 62, 633–647. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Gajewska, E.; Skłodowska, M. Effect of nickel on ROS content and antioxidative enzyme activities in wheat leaves. BioMetals 2007, 20, 27–36. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant. Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Gillespie, K.M.; Ainsworth, E.A. Measurement of reduced, oxidized and total ascorbate content in plants. Nat. Protoc. 2007, 2, 871–874. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. rapid and sensitive method for quantitation of microgram quantities of protein utilizing principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Donahue, J.L.; Okpodu, C.M.; Cramer, C.L.; Grabau, E.A.; Alscher, R.G. Responses of antioxidants to paraquat in pea leaves (relationships to resistance). Plant Physiol. 1997, 113, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Aebi, H. [13] Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen-peroxide is scavenged by ascorbate-specific peroxidase in spinach-chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Murshed, R.; Lopez-Lauri, F.; Sallanon, H. Microplate quantification of enzymes of the plant ascorbate-glutathione cycle. Anal. Biochem. 2008, 383, 320–322. [Google Scholar] [CrossRef] [PubMed]
- Pinto, E.; Soares, A.G.; Ferreira, I.M.P.L.V.O. Quantitative analysis of glyphosate, glufosinate and AMPA in irrigation water by: In situ derivatization-dispersive liquid-liquid microextraction combined with UPLC-MS/MS. Anal. Methods 2018, 10, 554–561. [Google Scholar] [CrossRef]
- Gomes, M.P.; Le Manac’h, S.G.; Maccario, S.; Labrecque, M.; Lucotte, M.; Juneau, P. Differential effects of glyphosate and aminomethylphosphonic acid (AMPA) on photosynthesis and chlorophyll metabolism in willow plants. Pestic. Biochem. Physiol. 2016, 130, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Zhong, G.; Wu, Z.; Yin, J.; Chai, L. Responses of Hydrilla verticillata (L.f.) Royle and Vallisneria natans (Lour.) Hara to glyphosate exposure. Chemosphere 2018, 193, 385–393. [Google Scholar] [CrossRef]
- Singh, H.; Singh, N.B.; Singh, A.; Hussain, I. Exogenous Application of Salicylic Acid to Alleviate Glyphosate Stress in Solanum lycopersicum. Int. J. Veg. Sci. 2017, 23, 552–566. [Google Scholar] [CrossRef]
- Soares, C.; Spormann, S.; Fidalgo, F. Salicylic acid improves the performance of the enzymatic antioxidant system of barley exposed to glyphosate. Free Radic. Biol. Med. 2018, 120, S157. [Google Scholar] [CrossRef]
- Fernandes, B.; Soares, C.; Braga, C.; Rebotim, A.; Ferreira, R.; Ferreira, J.; Fidalgo, F.; Pereira, R.; Cachada, A. Ecotoxicological assessment of a glyphosate-based herbicide in cover plants: Medicago sativa L. as a model species. Appl. Sci. 2020, 10, 5098. [Google Scholar] [CrossRef]
- Soares, C.; Pereira, R.; Martins, M.; Tamagnini, P.; Serôdio, J.; Moutinho-Pereira, J.; Cunha, A.; Fidalgo, F. Glyphosate-dependent effects on photosynthesis of Solanum lycopersicum L.—An ecophysiological, ultrastructural and molecular approach. J. Hazard. Mater. 2020, 398, 122871. [Google Scholar] [CrossRef]
- Gratão, P.L.; Polle, A.; Lea, P.J.; Azevedo, R.A. Making the life of heavy metal-stressed plants a little easier. Funct. Plant Biol. 2005, 32, 481. [Google Scholar] [CrossRef]
- Zhang, C.; Feng, Y.; Liu, Y.W.; Chang, H.Q.; Li, Z.J.; Xue, J.M. Uptake and translocation of organic pollutants in plants: A review. J. Integr. Agric. 2017, 16, 1659–1668. [Google Scholar] [CrossRef] [Green Version]
- Ahsan, N.; Lee, D.G.; Lee, K.W.; Alam, I.; Lee, S.H.; Bahk, J.D.; Lee, B.H. Glyphosate-induced oxidative stress in rice leaves revealed by proteomic approach. Plant Physiol. Biochem. 2008, 46, 1062–1070. [Google Scholar] [CrossRef] [PubMed]
- Mertens, M.; Höss, S.; Neumann, G.; Afzal, J.; Reichenbecher, W. Glyphosate, a chelating agent—relevant for ecological risk assessment? Environ. Sci. Pollut. Res. 2018, 25, 5298–5317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cakmak, I.; Yazici, A.; Tutus, Y.; Ozturk, L. Glyphosate reduced seed and leaf concentrations of calcium, manganese, magnesium, and iron in non-glyphosate resistant soybean. Eur. J. Agron. 2009, 31, 114–119. [Google Scholar] [CrossRef] [Green Version]
- Bellaloui, N.; Reddy, K.N.; Zablotowicz, R.M.; Abbas, H.K.; Abel, C.A. Effects of glyphosate application on seed iron and root ferric (III) reductase in soybean cultivars. J. Agric. Food Chem. 2009, 57, 9569–9574. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.S.; Ozturk, L.; Cakmak, I.; Budak, H. Turfgrass species response exposed to increasing rates of glyphosate application. Eur. J. Agron. 2009, 31, 120–125. [Google Scholar]
- Zobiole, L.; Kremer, R.J.; Oliveira, R.S., Jr.; Constantin, J. Glyphosate affects chlorophyll, nodulation and nutrient accumulation of “second generation” glyphosate-resistant soybean (Glycine max L.). Pestic. Biochem. Physiol. 2011, 99, 53–60. [Google Scholar] [CrossRef]
- Zobiole, L.; Kremer, R.J.; Oliveira, R.S., Jr.; Constantin, J. Glyphosate affects micro-organisms in rhizospheres of glyphosate-resistant soybeans. J. Appl. Microbiol. 2011, 110, 118–127. [Google Scholar] [CrossRef]
- Tzin, V.; Galili, G. New Insights into the shikimate and aromatic amino acids biosynthesis pathways in plants. Mol. Plant 2010, 3, 956–972. [Google Scholar] [CrossRef]
- della-Cioppa, G.; Bauer, S.C.; Taylor, M.L.; Rochester, D.E.; Klein, B.K.; Shah, D.M.; Fraley, R.T.; Kishore, G.M. Targeting a Herbicide–Resistant Enzyme from Escherichia coli to Chloroplasts of Higher Plants. Bio/Technology 1987, 5, 579–584. [Google Scholar] [CrossRef]
- Meharg, C.; Meharg, A.A. Silicon, the silver bullet for mitigating biotic and abiotic stress, and improving grain quality, in rice? Environ. Exp. Bot. 2015, 120, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Imtiaz, M.; Rizwan, M.S.; Mushtaq, M.A.; Ashraf, M.; Shahzad, S.M.; Yousaf, B.; Saeed, D.A.; Rizwan, M.; Nawaz, M.A.; Mehmood, S.; et al. Silicon occurrence, uptake, transport and mechanisms of heavy metals, minerals and salinity enhanced tolerance in plants with future prospects: A review. J. Environ. Manag. 2016, 183, 521–529. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.K.; Varma, R.K.; Singh, S.; Sachan, M.; Guerriero, G.; Kushwaha, B.K.; Bhardwaj, S.; Ramawat, N.; Sharma, S.; Singh, V.P.; et al. Silicon tackles butachlor toxicity in rice seedlings by regulating anatomical characteristics, ascorbate-glutathione cycle, proline metabolism and levels of nutrients. Sci. Rep. 2020, 10, 14078. [Google Scholar] [CrossRef]
- Malhotra, C.; Kapoor, R.T. Silicon: A sustainable tool in abiotic stress tolerance in plants. In Plant Abiotic Stress Tolerance: Agronomic, Molecular and Biotechnological Approaches; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 333–356. ISBN 9783030061180. [Google Scholar]
- Souri, Z.; Khanna, K.; Karimi, N.; Ahmad, P. Silicon and Plants: Current Knowledge and Future Prospects. J. Plant Growth Regul. 2020, 40, 906–925. [Google Scholar] [CrossRef]
- Sonobe, K.; Hattori, T.; An, P.; Tsuji, W.; Eneji, A.E.; Kobayashi, S.; Kawamura, Y.; Tanaka, K.; Inanaga, S. Effect of silicon application on sorghum root responses to water stress. J. Plant Nutr. 2011, 34, 71–82. [Google Scholar] [CrossRef]
- Liu, P.; Yin, L.; Deng, X.; Wang, S.; Tanaka, K.; Zhang, S. Aquaporin-mediated increase in root hydraulic conductance is involved in silicon-induced improved root water uptake under osmotic stress in Sorghum bicolor L. J. Exp. Bot. 2014, 65, 4747–4756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon alleviates drought stress of rice plants by improving plant water status, photosynthesis and mineral nutrient absorption. Biol. Trace Elem. Res. 2011, 142, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Schröder, P.; Lyubenova, L.; Huber, C. Do heavy metals and metalloids influence the detoxification of organic xenobiotics in plants? Environ. Sci. Pollut. Res. 2009, 16, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Guo, J.; Hu, Y.; Gong, H. Distinct physiological responses of tomato and cucumber plants in silicon-mediated alleviation of cadmium stress. Front. Plant Sci. 2015, 6, 453. [Google Scholar] [CrossRef] [Green Version]
- Alzahrani, Y.; Kuşvuran, A.; Alharby, H.F.; Kuşvuran, S.; Rady, M.M. The defensive role of silicon in wheat against stress conditions induced by drought, salinity or cadmium. Ecotoxicol. Environ. Saf. 2018, 154, 187–196. [Google Scholar] [CrossRef]
- Hussain, A.; Rizwan, M.; Ali, Q.; Ali, S. Seed priming with silicon nanoparticles improved the biomass and yield while reduced the oxidative stress and cadmium concentration in wheat grains. Environ. Sci. Pollut. Res. 2019, 26, 7579–7588. [Google Scholar] [CrossRef]
- Ashfaque, F.; Inam, A.; Inam, A.; Iqbal, S.; Sahay, S. Response of silicon on metal accumulation, photosynthetic inhibition and oxidative stress in chromium-induced mustard (Brassica juncea L.). S. Afr. J. Bot. 2017, 111, 153–160. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K. Silicon nanoparticles (SiNp) alleviate chromium (VI) phytotoxicity in Pisum sativum (L.) seedlings. Plant Physiol. Biochem. 2015, 96, 189–198. [Google Scholar] [CrossRef]
- Pontigo, S.; Godoy, K.; Jiménez, H.; Gutiérrez-Moraga, A.; Mora, M.D.L.L.; Cartes, P. Silicon-Mediated Alleviation of Aluminum Toxicity by Modulation of Al/Si Uptake and Antioxidant Performance in Ryegrass Plants. Front. Plant Sci. 2017, 8, 642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sousa, A.; Saleh, A.M.; Habeeb, T.H.; Hassan, Y.M.; Zrieq, R.; Wadaan, M.A.M.; Hozzein, W.N.; Selim, S.; Matos, M.; AbdElgawad, H. Silicon dioxide nanoparticles ameliorate the phytotoxic hazards of aluminum in maize grown on acidic soil. Sci. Total Environ. 2019, 693, 133636. [Google Scholar] [CrossRef]
- Liu, J.; Cai, H.; Mei, C.; Wang, M. Effects of nano-silicon and common silicon on lead uptake and translocation in two rice cultivars. Front. Environ. Sci. Eng. 2015, 9, 905–911. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. Silicon nanoparticles more effectively alleviated UV-B stress than silicon in wheat (Triticum aestivum) seedlings. Plant Physiol. Biochem. 2017, 110, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Haghighi, M.; Pessarakli, M. Influence of silicon and nano-silicon on salinity tolerance of cherry tomatoes (Solanum lycopersicum L.) at early growth stage. Sci. Hortic. 2013, 161, 111–117. [Google Scholar] [CrossRef]
- Caverzan, A.; Piasecki, C.; Chavarria, G.; Stewart, C.N.; Vargas, L. Defenses against ROS in crops and weeds: The effects of interference and herbicides. Int. J. Mol. Sci. 2019, 20, 1086. [Google Scholar] [CrossRef] [Green Version]
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Khan, A.L.; Waqas, M.; Lee, I.-J. Silicon Regulates Antioxidant Activities of Crop Plants under Abiotic-Induced Oxidative Stress: A Review. Front. Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, C.; Carvalho, M.E.A.; Azevedo, R.A.; Fidalgo, F. Plants facing oxidative challenges—A little help from the antioxidant networks. Environ. Exp. Bot. 2019, 161, 4–25. [Google Scholar] [CrossRef]
- Poustini, K.; Siosemardeh, A.; Ranjbar, M. Proline accumulation as a response to salt stress in 30 wheat (Triticum aestivum L.) cultivars differing in salt tolerance. Genet. Resour. Crop Evol. 2007, 54, 925–934. [Google Scholar] [CrossRef]
- Hare, P.D.; Cress, W.A. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul. 1997, 21, 79–102. [Google Scholar] [CrossRef]
- Abdel-Haliem, M.E.F.; Hegazy, H.S.; Hassan, N.S.; Naguib, D.M. Effect of silica ions and nano silica on rice plants under salinity stress. Ecol. Eng. 2017, 99, 282–289. [Google Scholar] [CrossRef]
- Sousa, B.; Soares, C.; Oliveira, F.; Martins, M.; Branco-Neves, S.; Barbosa, B.; Ataíde, I.; Teixeira, J.; Azenha, M.; Azevedo, R.A.; et al. Response of Solanum lycopersicum L. to diclofenac—Impacts on the plant’s antioxidant mechanisms. Environ. Pollut. 2020, 258, 113762. [Google Scholar] [CrossRef]
- Kapoor, D.; Singh, S.; Kumar, V.; Romero, R.; Prasad, R.; Singh, J. Antioxidant enzymes regulation in plants in reference to reactive oxygen species (ROS) and reactive nitrogen species (RNS). Plant Gene 2019, 19, 100182. [Google Scholar] [CrossRef]
- Soares, C.; Branco-Neves, S.; de Sousa, A.; Teixeira, J.; Pereira, R.; Fidalgo, F. Can nano-SiO2 reduce the phytotoxicity of acetaminophen?—A physiological, biochemical and molecular approach. Environ. Pollut. 2018, 241, 900–911. [Google Scholar] [CrossRef]
- Backer, B.; Zhong, G.; Wu, Z.; Liu, N.; Yin, J. Phosphate alleviation of glyphosate-induced toxicity in Hydrocharis dubia. Aquat. Toxicol. 2018, 201, 91–98. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Basantani, M.; Srivastava, A. Plant glutathione transferases—A decade falls short. Can. J. Bot. 2007, 85, 443–456. [Google Scholar] [CrossRef]
- Singh, A.; Prasad, S.M.; Singh, R.P. Plant Responses to Xenobiotics; Springer: Singapore, 2016; pp. 1–346. [Google Scholar] [CrossRef]
- Ramel, F.; Sulmon, C.; Serra, A.-A.; Gouesbet, G.; Couée, I. Xenobiotic sensing and signalling in higher plants. J. Exp. Bot. 2012, 63, 3999–4014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K. Silicon nanoparticles more efficiently alleviate arsenate toxicity than silicon in maize cultiver and hybrid differing in arsenate tolerance. Front. Environ. Sci. 2016, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- Andreani, T.; Nogueira, V.; Gavina, A.; Fernandes, S.; Rodrigues, J.L.; Pinto, V.V.; Ferreira, M.J.; Silva, A.M.; Pereira, C.M.; Pereira, R. Ecotoxicity to freshwater organisms and cytotoxicity of nanomaterials: Are we generating sufficient data for their risk assessment? Nanomaterials 2021, 11, 66. [Google Scholar] [CrossRef] [PubMed]
- Andreani, T.; Fernandes, P.M.V.; Nogueira, V.; Pinto, V.V.; Ferreira, M.J.; Rasteiro, M.G.; Pereira, R.; Pereira, C.M. The critical role of the dispersant agents in the preparation and ecotoxicity of nanomaterial suspensions. Environ. Sci. Pollut. Res. 2020, 27, 19845–19857. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Precursor Ion (m/z) | Product Ion (m/z) | Cone Voltage (V) | Collision Energy (V) |
|---|---|---|---|---|
| GLY-FMOC | 392.2 | Q:88.0 | 20 | 20 |
| q:170.0 | 20 | 10 | ||
| 1,2-13C2, 15N GLY-FMOC | 395.2 | 91.0 | 20 | 20 |
| AMPA-FMOC | 334.0 | Q:112.1 | 20 | 15 |
| q:179.1 | 20 | 20 | ||
| 13C,15N-AMPA | 336.0 | 114.1 | 20 | 15 |
| Organ | Treatment | Total AsA | AsA/Total AsA | DHA/Total AsA | AsA/DHA |
|---|---|---|---|---|---|
| Shoots | CTL | 1.52 ± 0.09 b | 0.72 ± 0.05 a | 0.28 ± 0.05 a | 3.44 ± 0.66 a |
| Si | 1.74 ± 0.10 ab | 0.59 ± 0.06 a | 0.41 ± 0.06 a | 1.55 ± 0.40 b | |
| Nano-SiO2 | 1.51 ± 0.10 b | 0.56 ± 0.05 a | 0.44 ± 0.05 a | 1.31 ± 0.27 b | |
| GLY | 2.04 ± 0.12 a | 0.56 ± 0.03 a | 0.44 ± 0.03 a | 1.47 ± 0.22 b | |
| GLY + Si | 1.90 ± 0.12 ab | 0.60 ± 0.05 a | 0.40 ± 0.05 a | 1.48 ± 0.30 b | |
| GLY + nano-SiO2 | 2.14 ± 0.08 a | 0.63 ± 0.02 a | 0.37 ± 0.02 a | 1.65 ± 0.19 b | |
| Roots | CTL | 0.40 ± 0.02 b | 0.41 ± 0.03 bc | 0.59 ± 0.03 ab | 0.77 ± 0.05 b |
| Si | 0.46 ± 0.02 ab | 0.33 ± 0.04 c | 0.67 ± 0.04 a | 0.42 ± 0.06 c | |
| Nano-SiO2 | 0.55 ± 0.03 a | 0.33 ± 0.03 bc | 0.67 ± 0.03 a | 0.38 ± 0.04 c | |
| GLY | 0.45 ± 0.02 ab | 0.50 ± 0.02 a | 0.50 ± 0.02 bc | 1.02 ± 0.04 a | |
| GLY + Si | 0.37 ± 0.03 b | 0.45 ± 0.01 ab | 0.49 ± 0.02 c | 0.79 ± 0.03 b | |
| GLY + nano-SiO2 | 0.41 ± 0.02 b | 0.44 ± 0.01 ab | 0.56 ± 0.01 abc | 0.80 ± 0.05 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, C.; Nadais, P.; Sousa, B.; Pinto, E.; Ferreira, I.M.P.L.V.O.; Pereira, R.; Fidalgo, F. Silicon Improves the Redox Homeostasis to Alleviate Glyphosate Toxicity in Tomato Plants—Are Nanomaterials Relevant? Antioxidants 2021, 10, 1320. https://doi.org/10.3390/antiox10081320
Soares C, Nadais P, Sousa B, Pinto E, Ferreira IMPLVO, Pereira R, Fidalgo F. Silicon Improves the Redox Homeostasis to Alleviate Glyphosate Toxicity in Tomato Plants—Are Nanomaterials Relevant? Antioxidants. 2021; 10(8):1320. https://doi.org/10.3390/antiox10081320
Chicago/Turabian StyleSoares, Cristiano, Pedro Nadais, Bruno Sousa, Edgar Pinto, Isabel M. P. L. V. O. Ferreira, Ruth Pereira, and Fernanda Fidalgo. 2021. "Silicon Improves the Redox Homeostasis to Alleviate Glyphosate Toxicity in Tomato Plants—Are Nanomaterials Relevant?" Antioxidants 10, no. 8: 1320. https://doi.org/10.3390/antiox10081320
APA StyleSoares, C., Nadais, P., Sousa, B., Pinto, E., Ferreira, I. M. P. L. V. O., Pereira, R., & Fidalgo, F. (2021). Silicon Improves the Redox Homeostasis to Alleviate Glyphosate Toxicity in Tomato Plants—Are Nanomaterials Relevant? Antioxidants, 10(8), 1320. https://doi.org/10.3390/antiox10081320