


Enhancing Bioactive Components of Euryale ferox with Lactobacillus curvatus to Reduce H2O2-Induced Oxidative Stress in Human Skin Fibroblasts

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Microorganism Inoculum
2.3. Fermentation with Lactobacillus Curvatus
2.4. Euryale ferox Bioactive Component Assay
2.5. Determination of Molecular Mass of Polypeptides
2.6. Determination of Molecular Mass of Polysaccharides
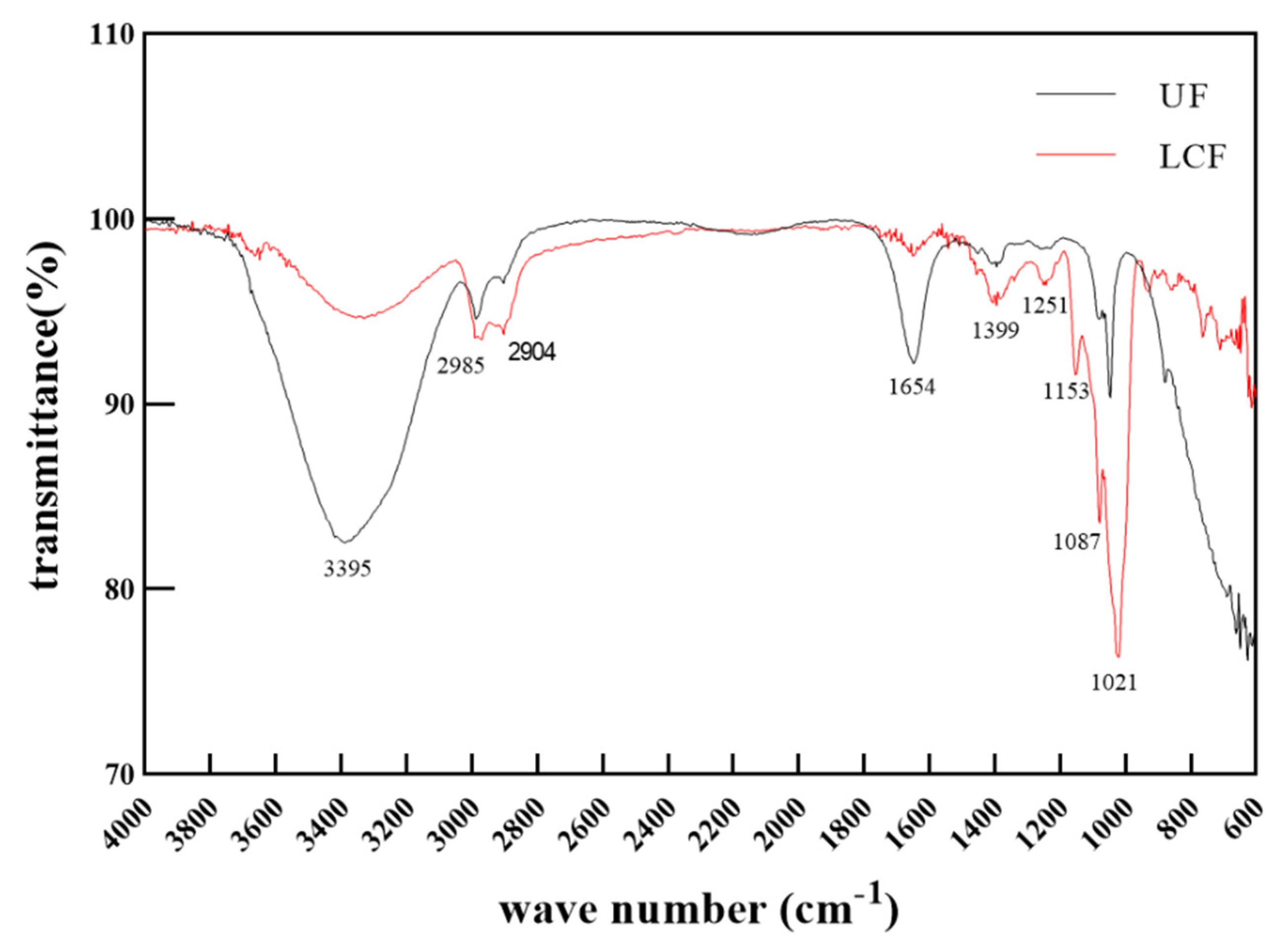
2.7. FT-IR Analysis of Polysaccharides
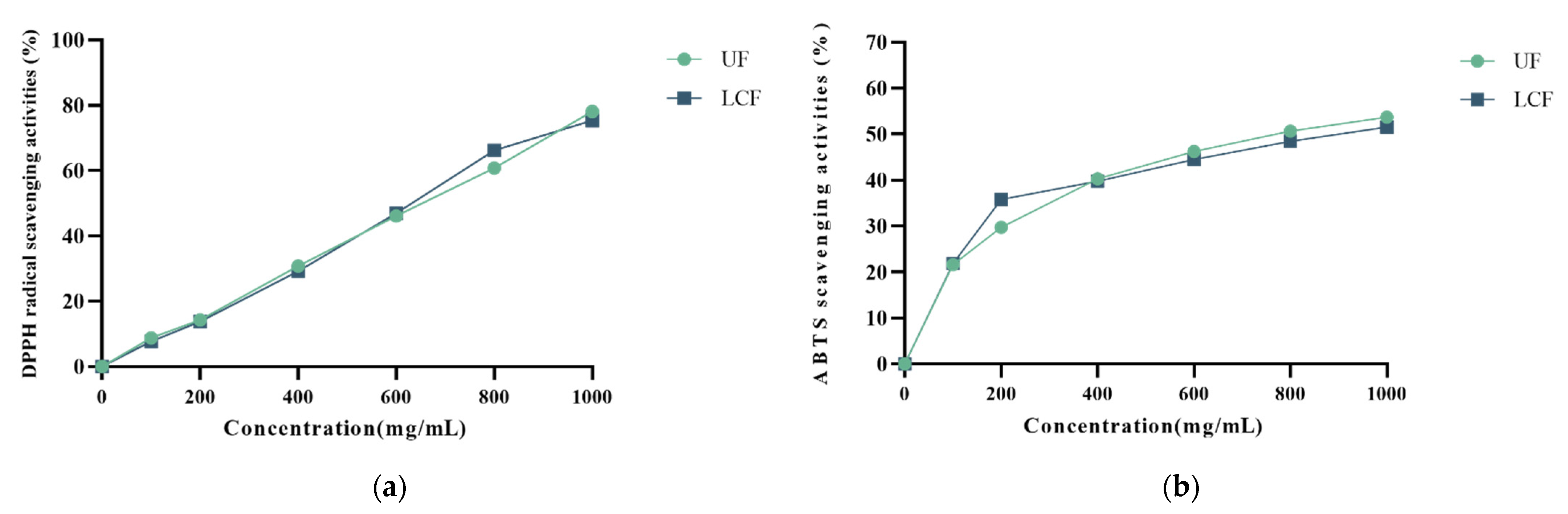
2.8. DPPH and ABTS Scavenging Activity
2.9. Establishment of Cell Culture and H2O2 Damage Model
2.9.1. Cell Culture
2.9.2. H2O2 Damage Model
2.10. Cell Viability Determination
2.11. Determination of Total Antioxidant Capacity and ROS
2.12. Antioxidant Enzymes and ELISA
2.13. Statistical Analysis
3. Results
3.1. Changes in Content of Bioactive Components in Euryale ferox
3.2. Spectroscopy Analysis of Euryale ferox
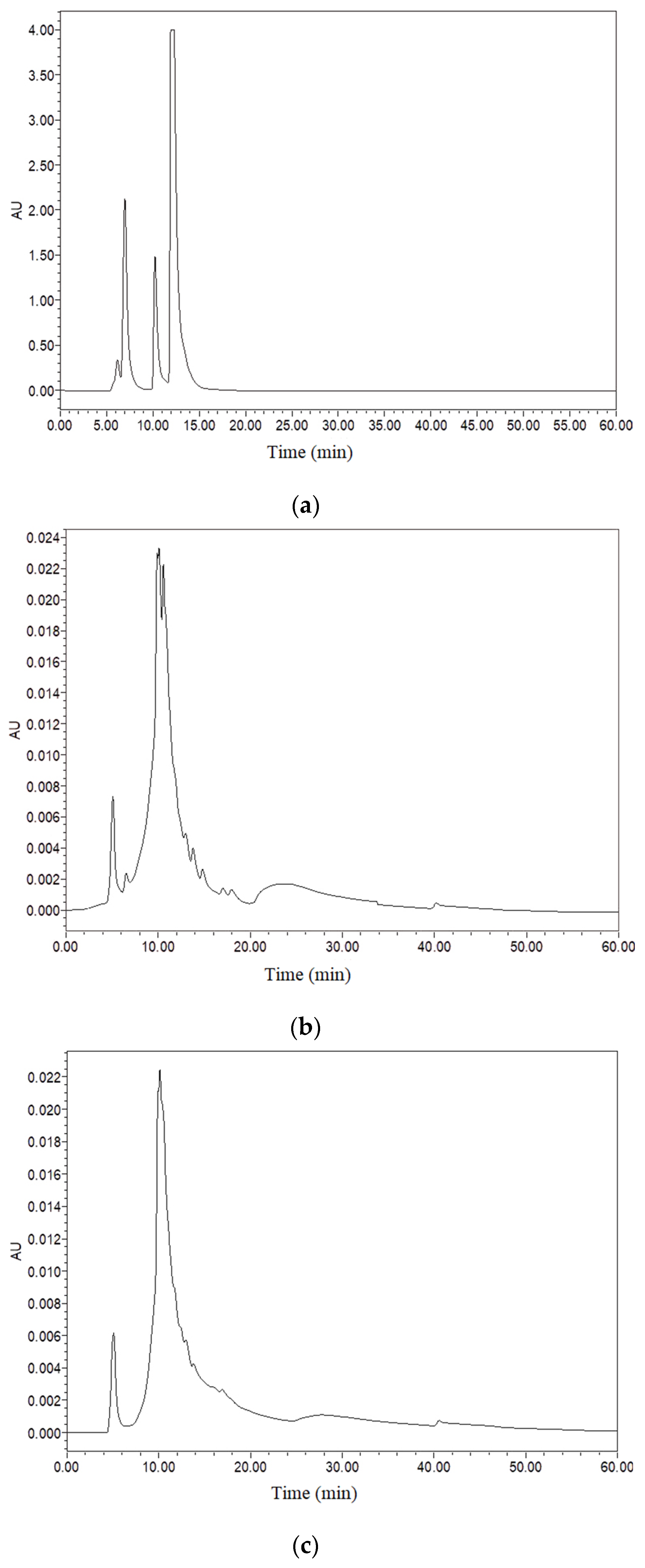
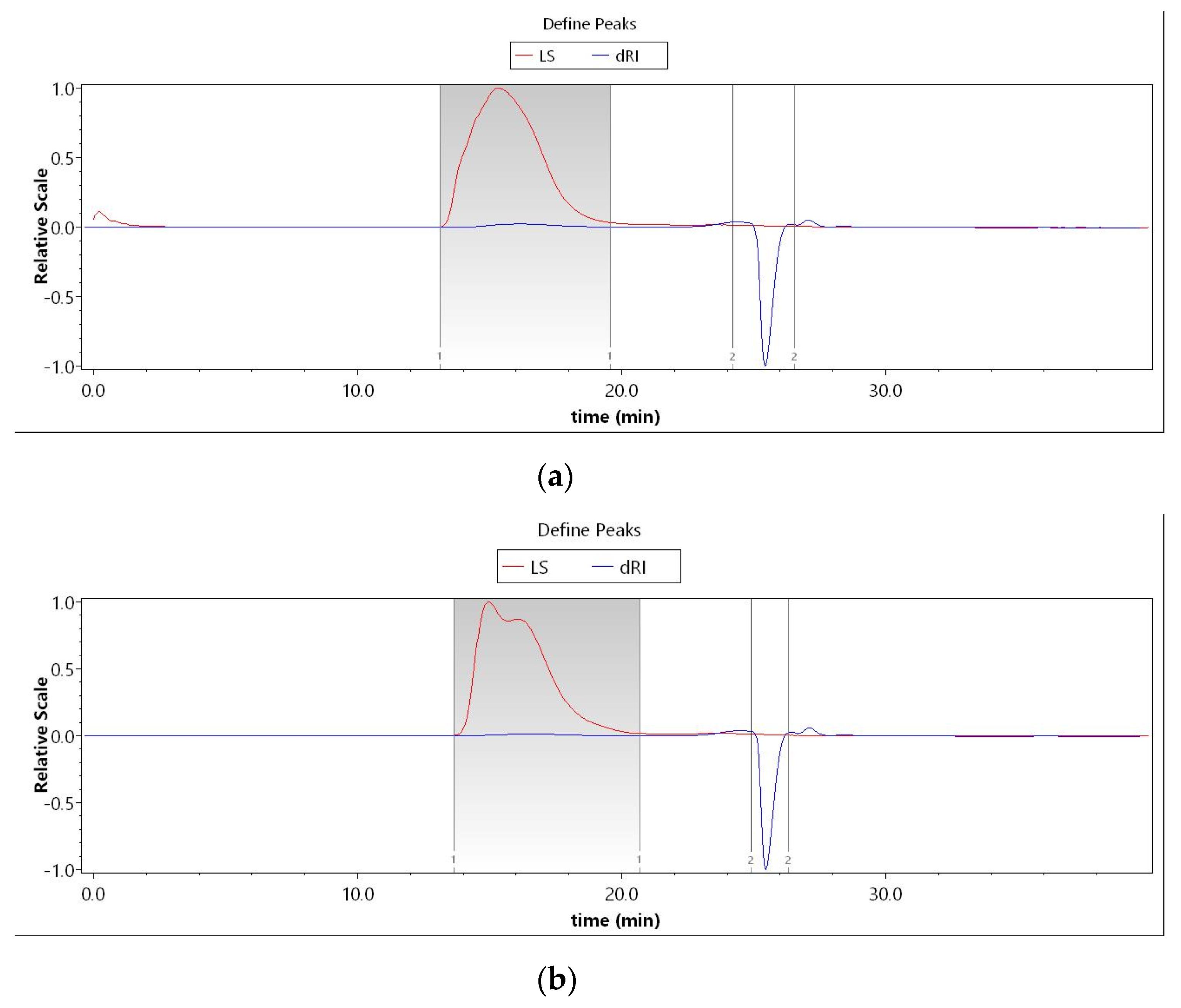
3.2.1. Polypeptide and Polysaccharide Molecular Weights
3.2.2. Characterization of Molecular Structures
3.3. Scavenging Free Radicals
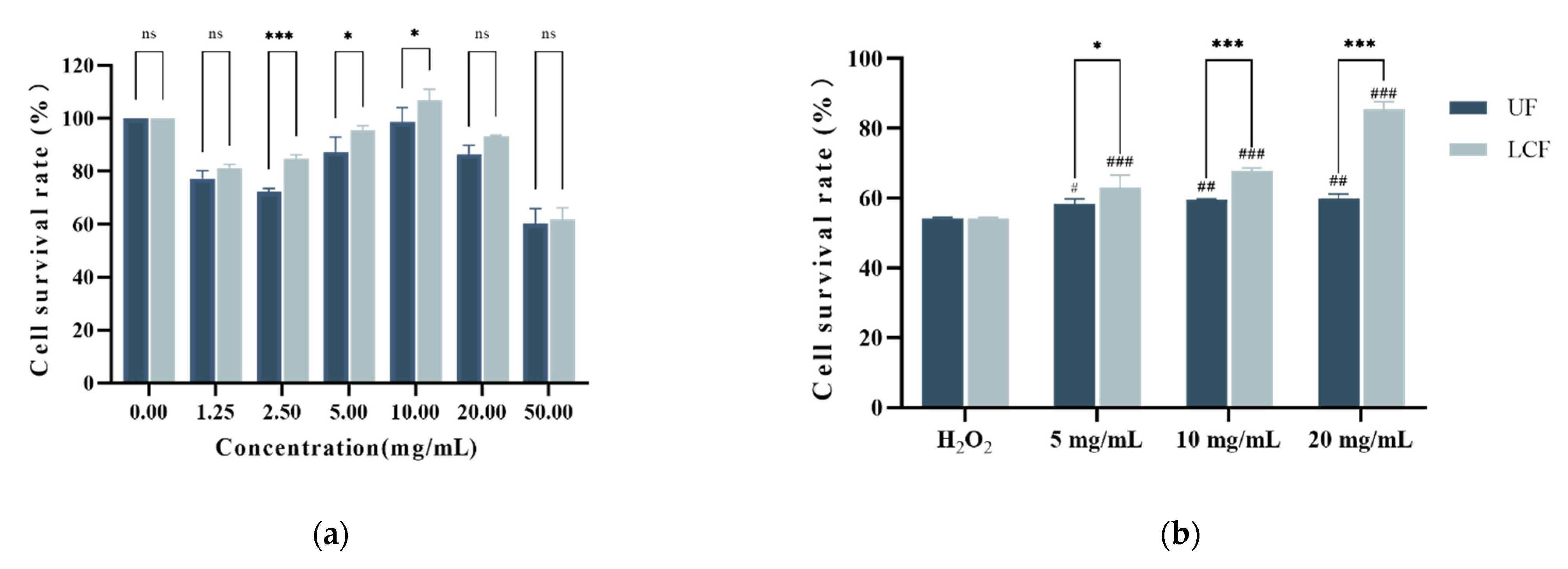
3.4. Effects of UF and LCF on HSF Cell Viability
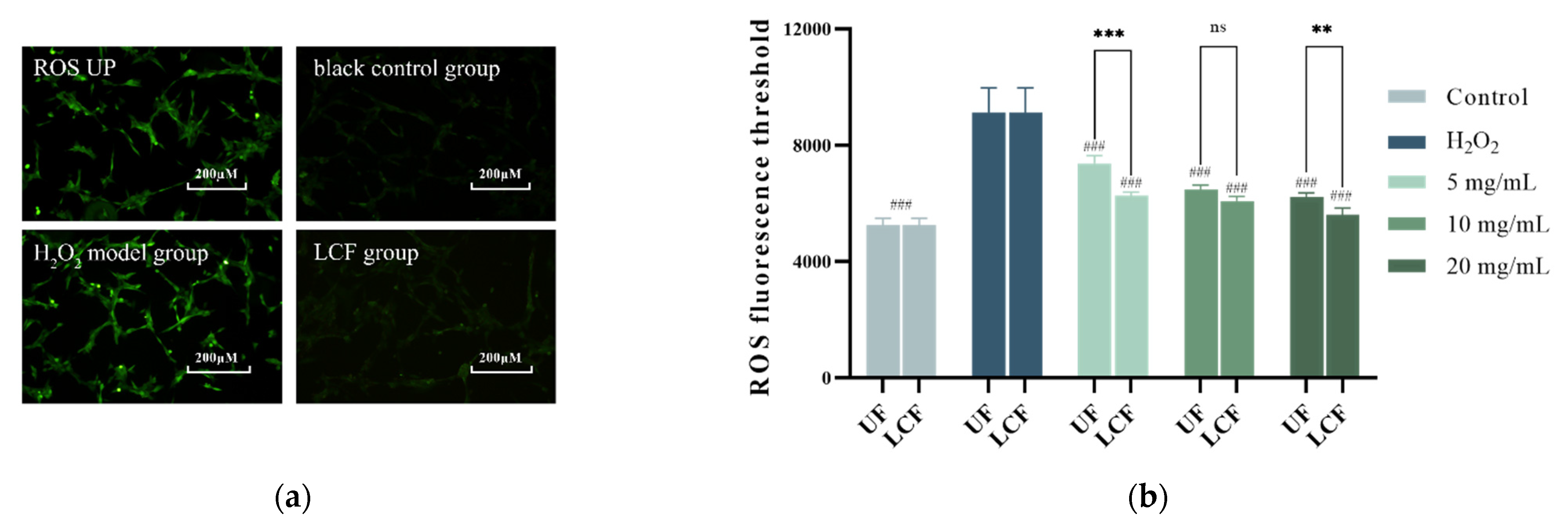
3.5. Effects of UF and LCF on ROS Level
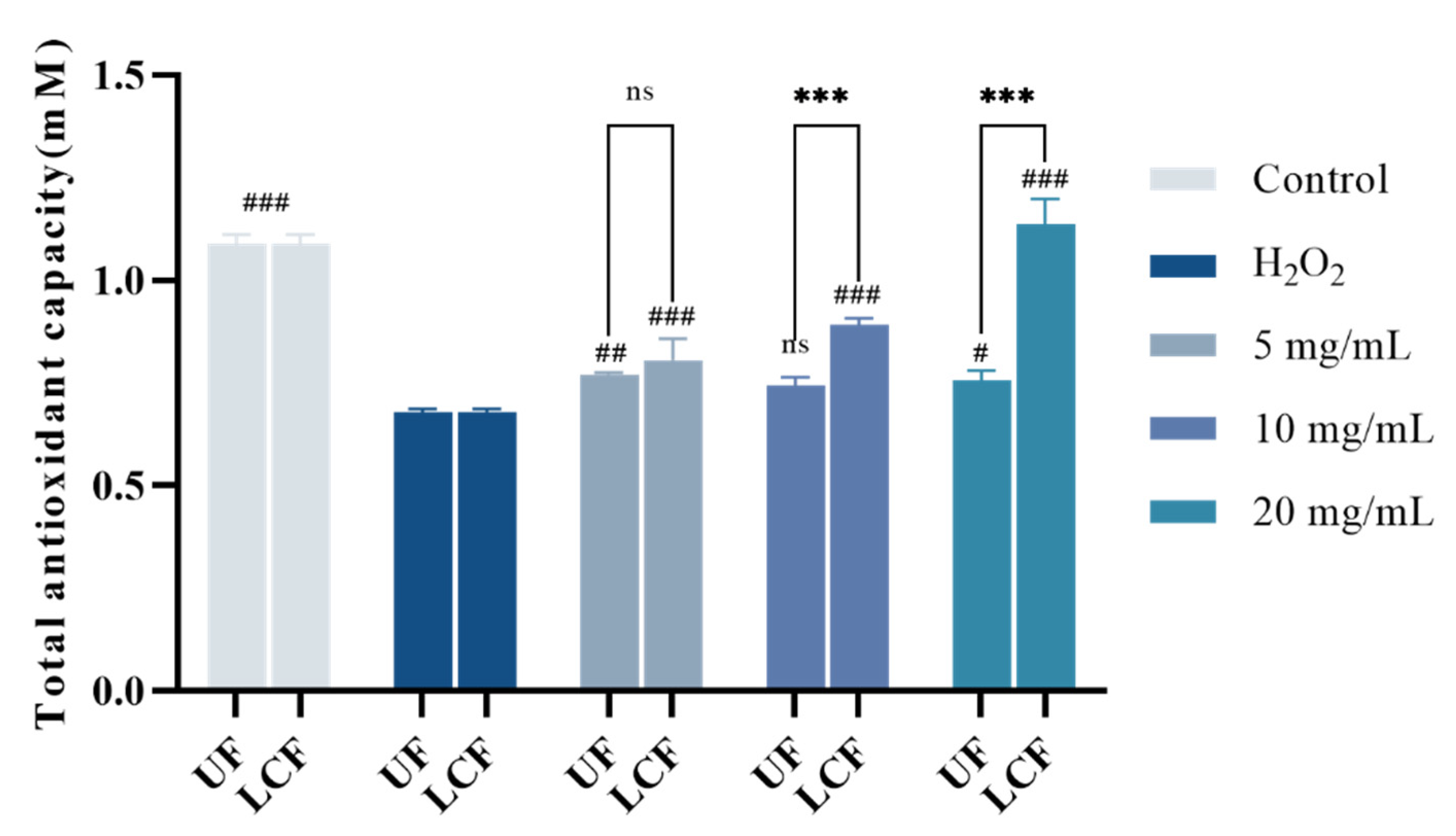
3.6. Effects of UF and LCF on Cellular Antioxidant Capacity
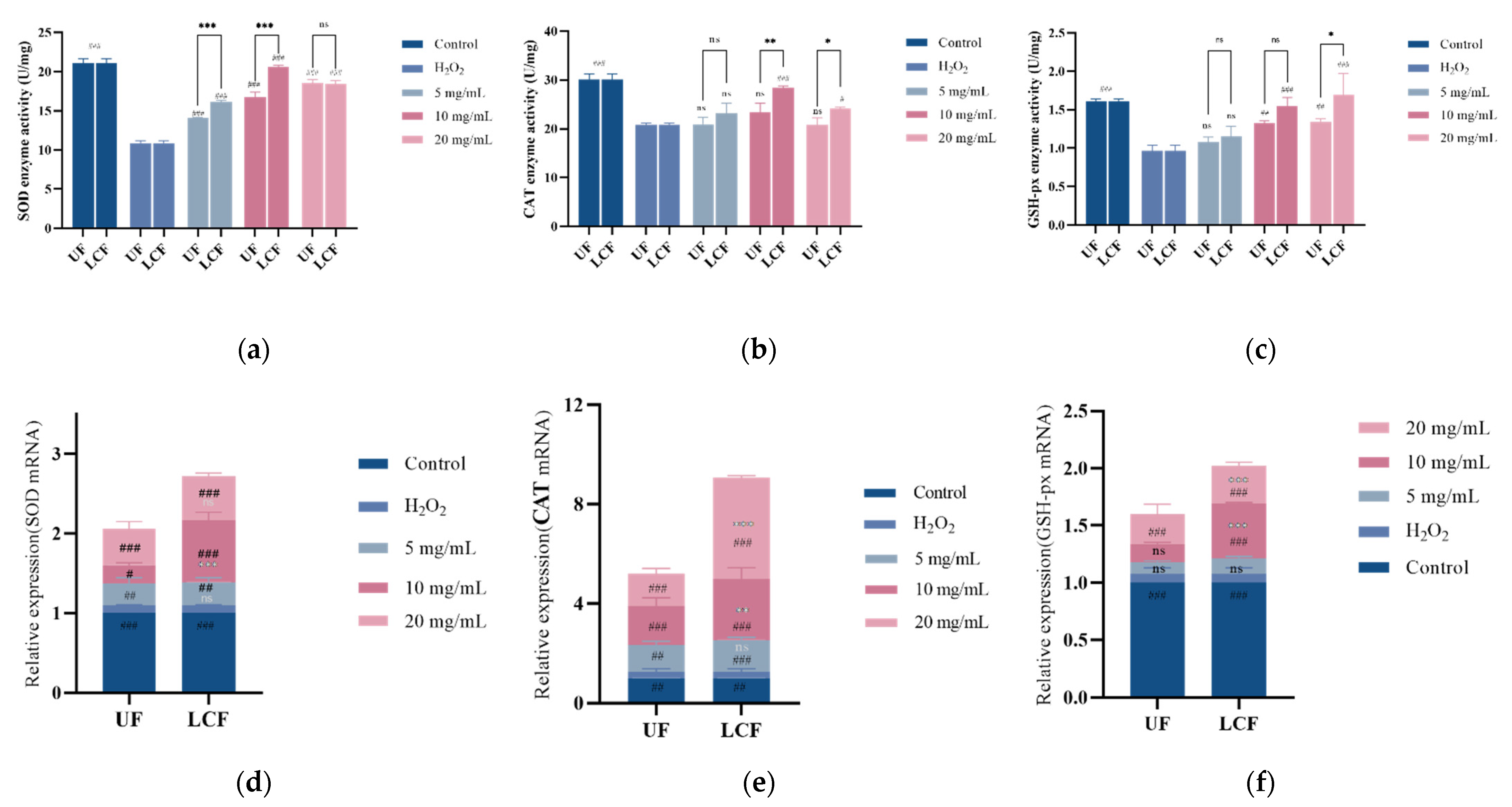
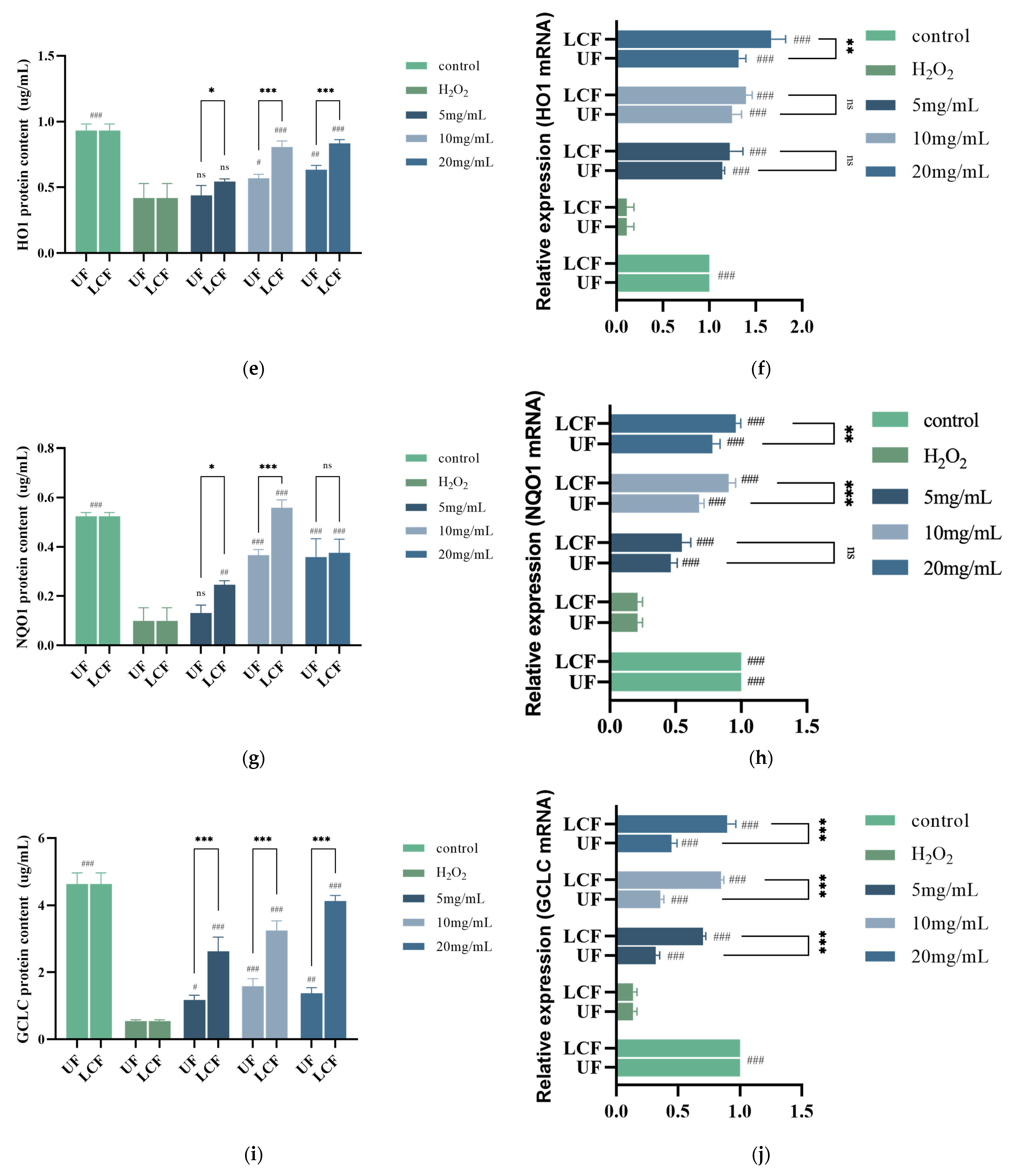
3.7. Effects of UF and LCF on Antioxidant Enzyme Content and mRNA Expression
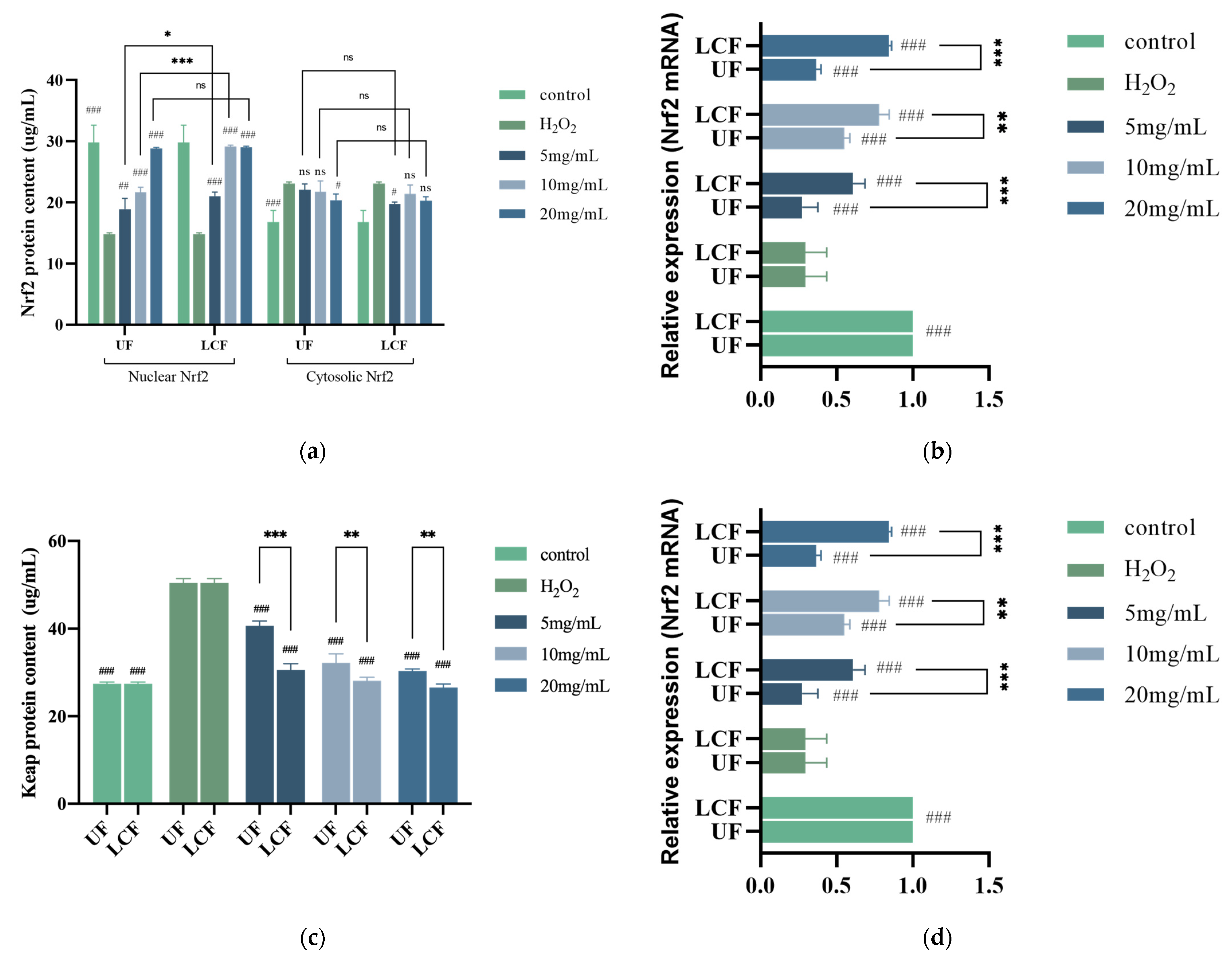
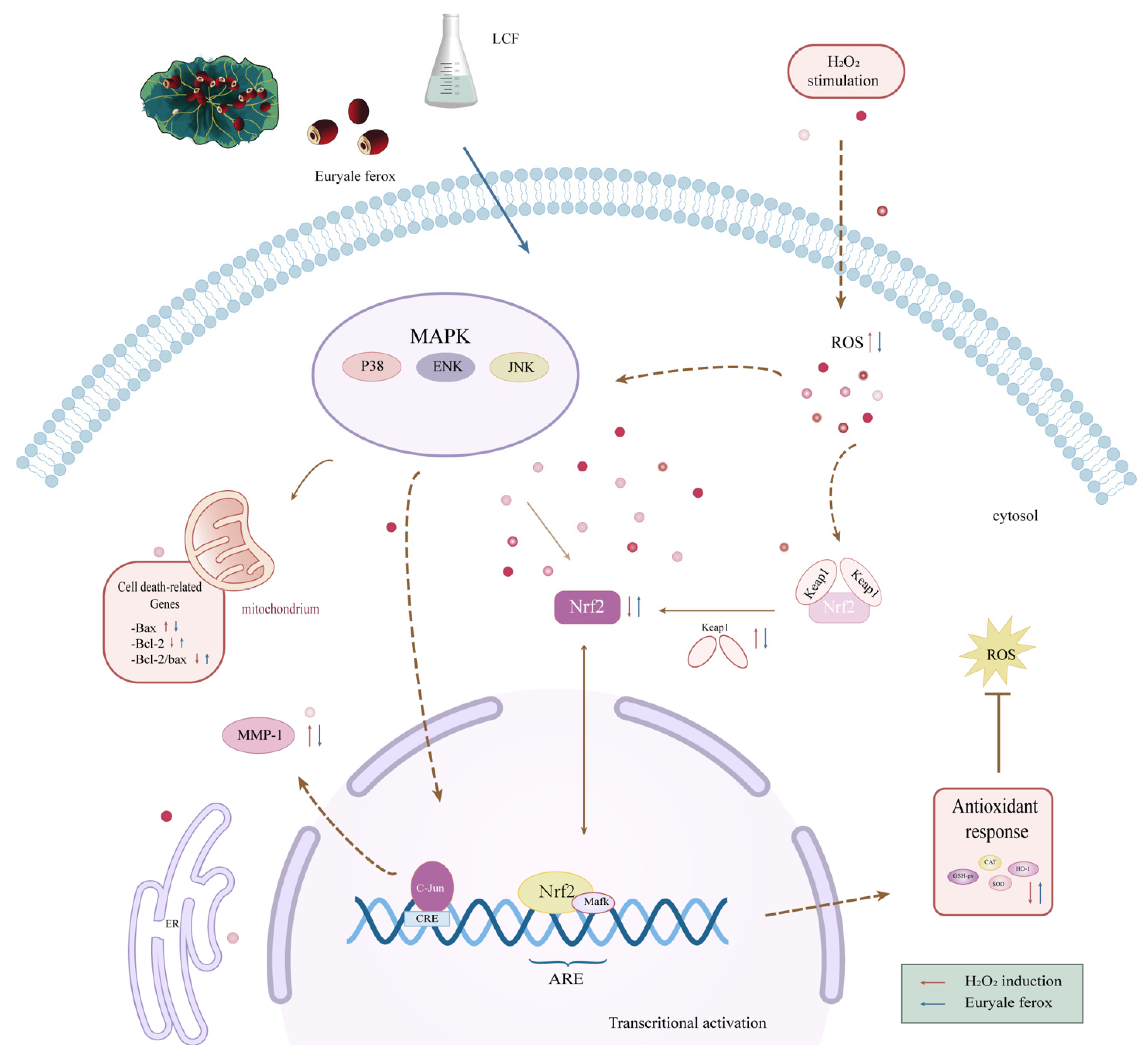
3.8. Effects of UF and LCF on Nrf2/Keap1 Signaling Pathway
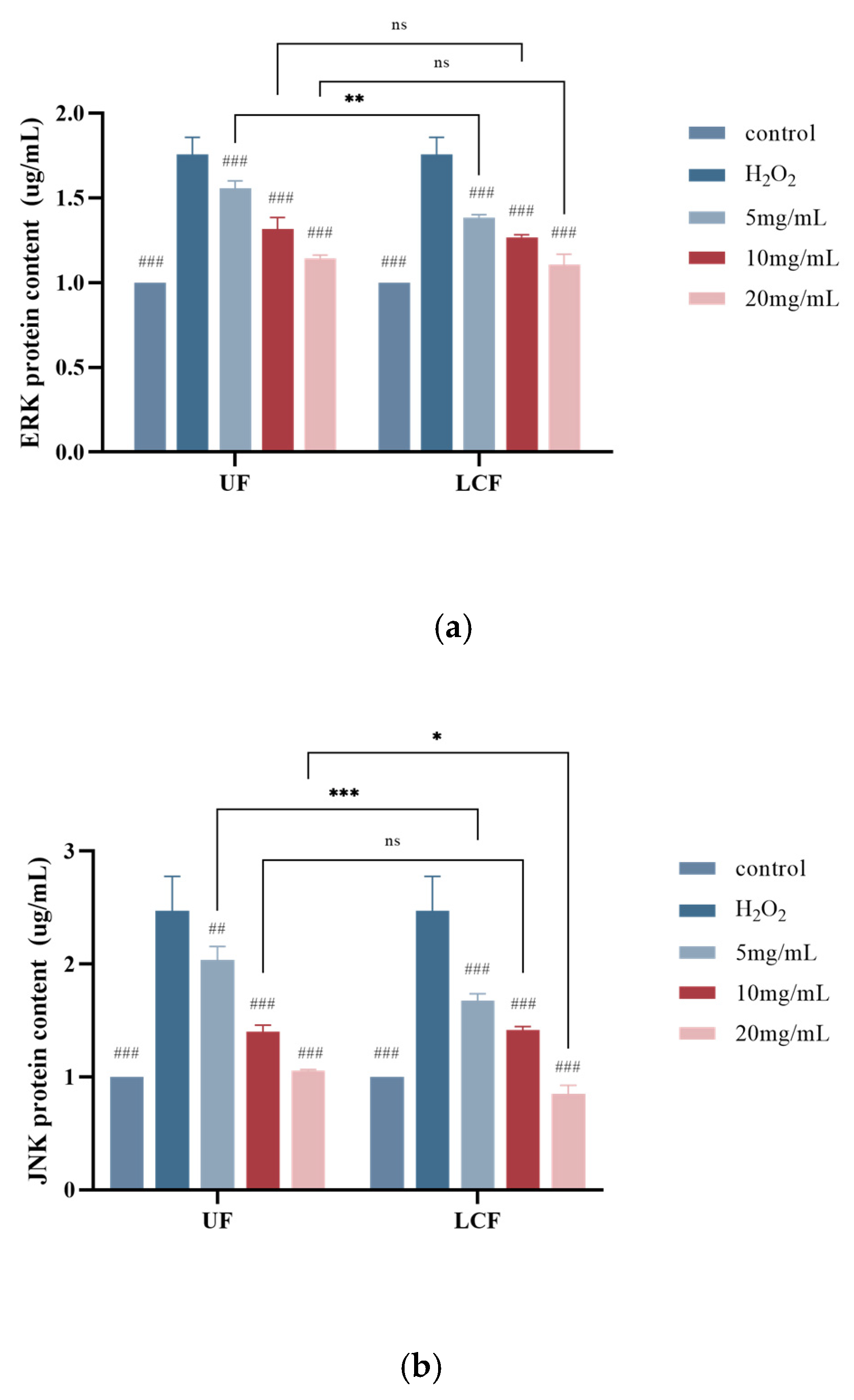
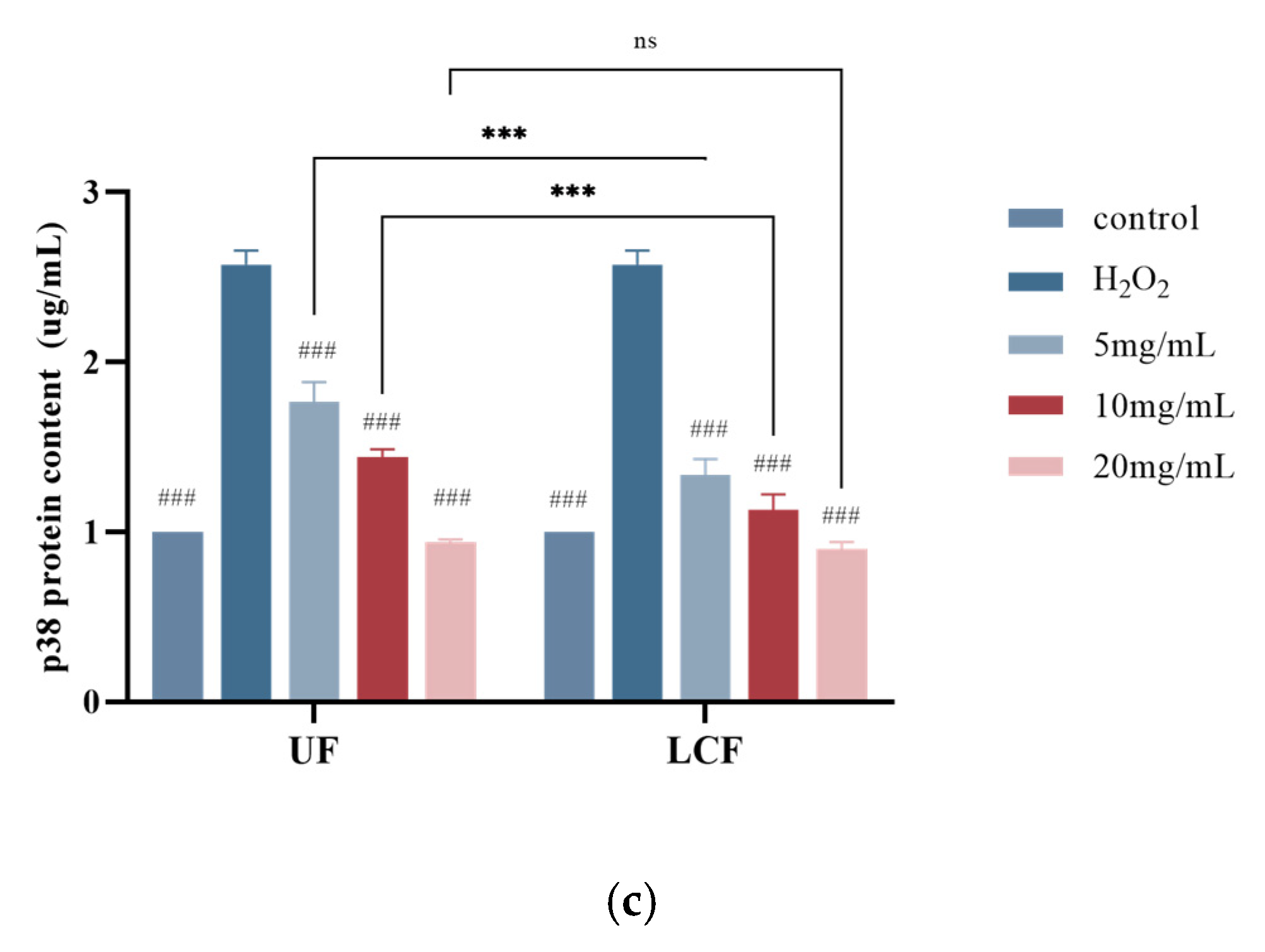
3.9. Effects of UF and LCF on MAPK Signaling Pathway
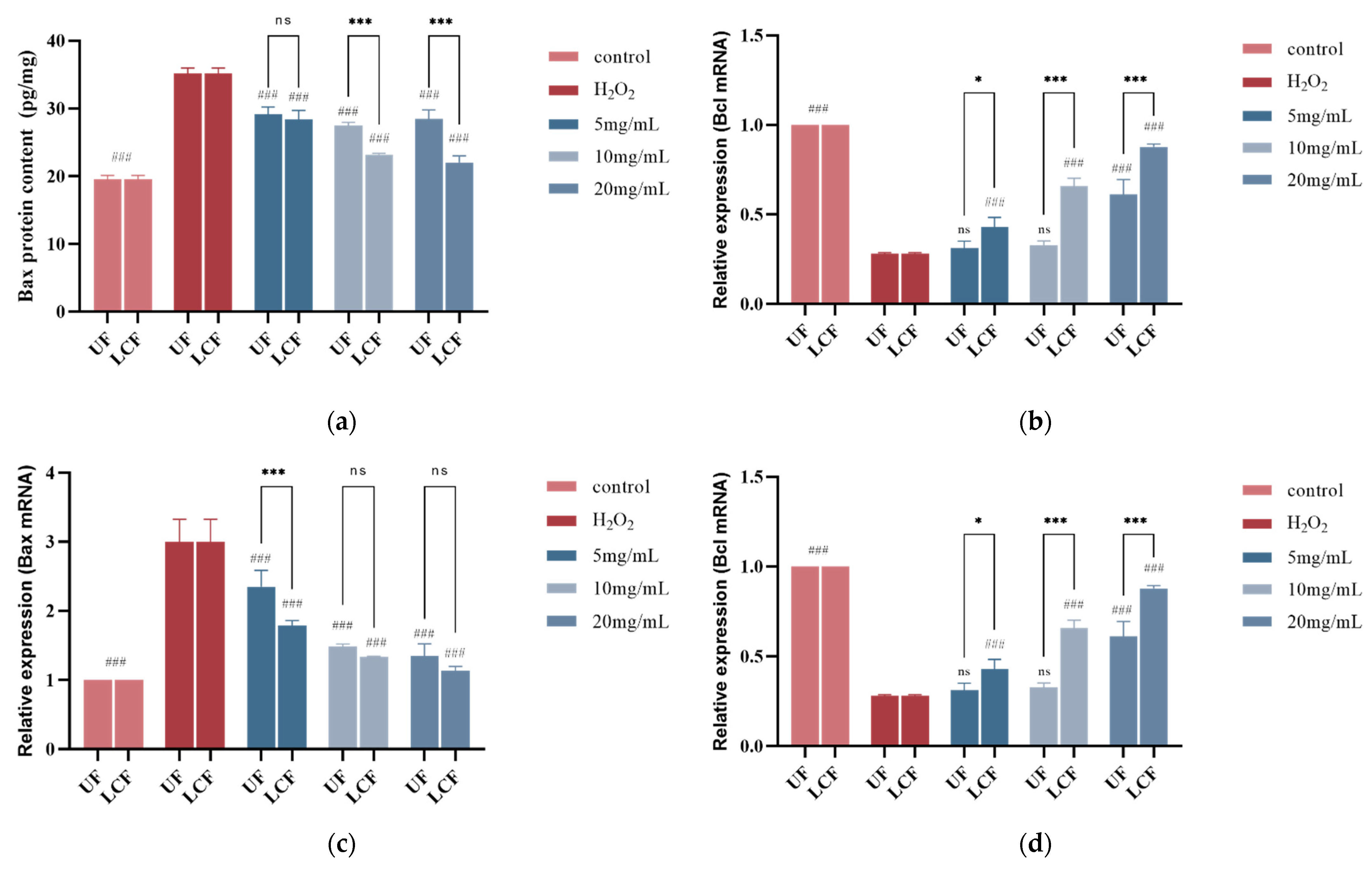
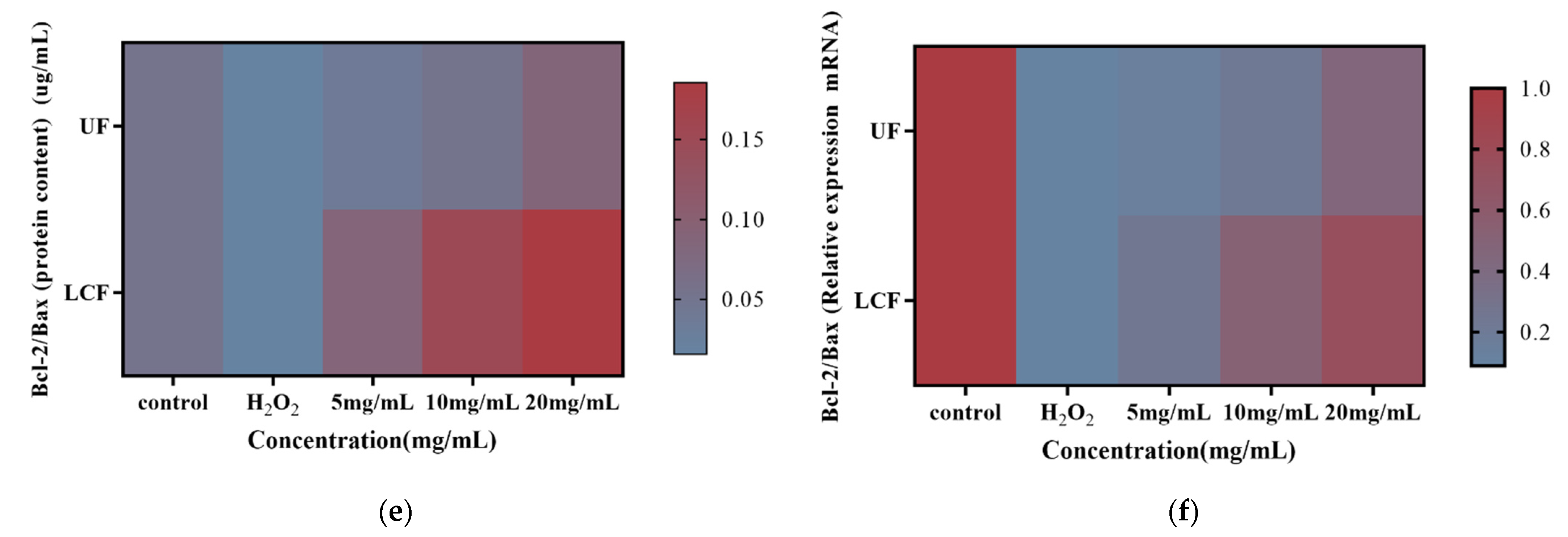
3.10. Effects of UF and LCF on Bax and Bcl-2 Protein Content
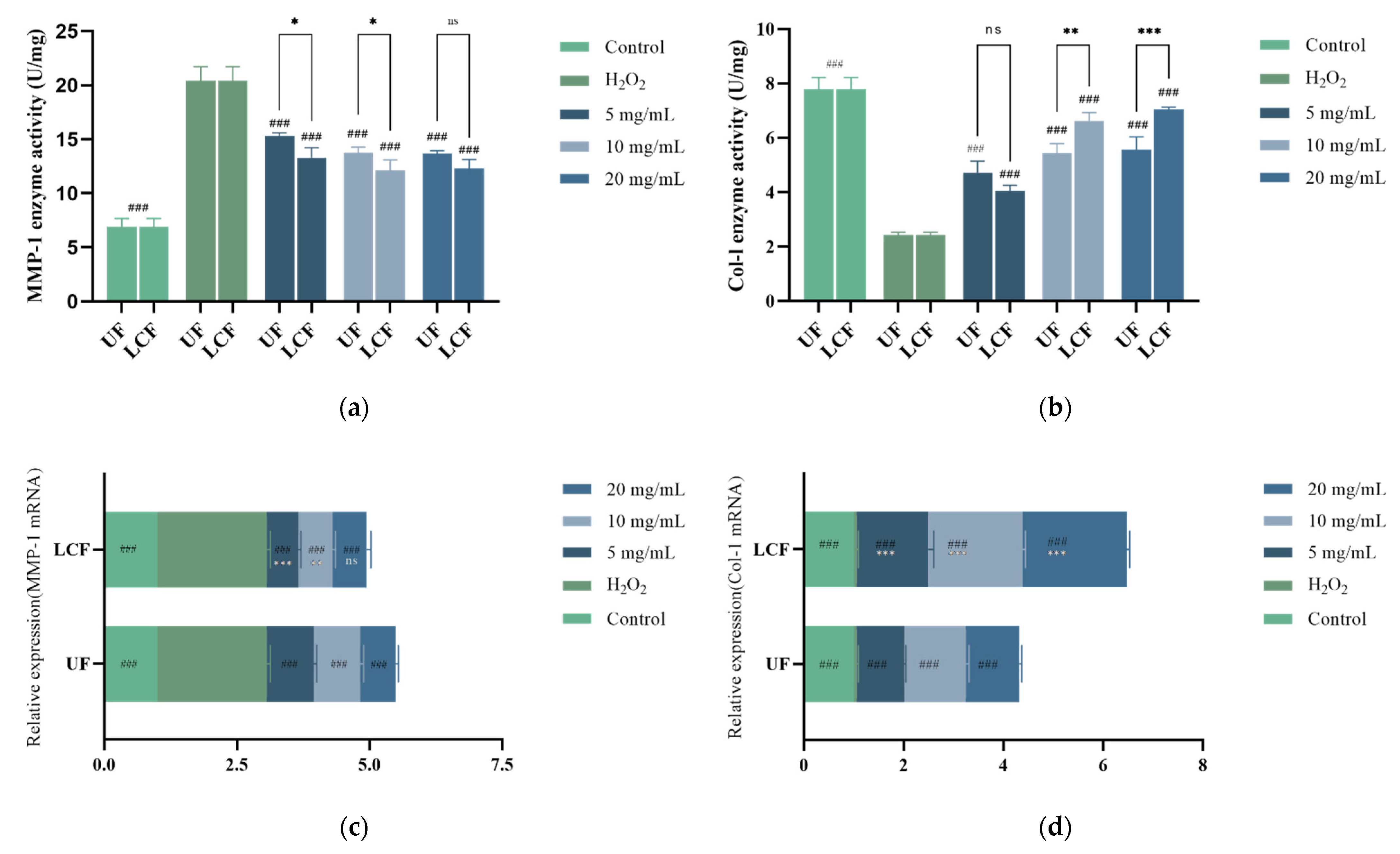
3.11. Effects of UF and LCF on Content and Expression Level of MMP-1 and COL-1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Averina, O.V.; Poluektova, E.U.; Marsova, M.V.; Danilenko, V.N. Biomarkers and Utility of the Antioxidant Potential of Probiotic Lactobacilli and Bifidobacteria as Representatives of the Human Gut Microbiota. Biomedicines 2021, 9, 1340. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Chen, R.; Wang, X.S.; Shen, B.; Yue, W.; Wu, Q. Antioxidant and anti-fatigue activities of phenolic extract from the seed coat of Euryale ferox Salisb. and identification of three phenolic compounds by LC-ESI-MS/MS. Molecules 2013, 18, 11003–11021. [Google Scholar] [CrossRef] [PubMed]
- Düz, M.; Doğan, Y.N.; Doğan, İ. Antioxidant activitiy of Lactobacillus plantarum, Lactobacillus sake and Lactobacillus curvatus strains isolated from fermented turkish sucuk. An. Acad. Bras. Cienc. 2020, 92, e20200105. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Ju, E.M.; Kim, J.H. Antioxidant activity of extracts from Euryale ferox seed. Exp. Mol. Med. 2002, 34, 100–106. [Google Scholar] [CrossRef]
- He, S.; Wang, D.; Zhang, Y.; Yang, S.; Li, X.; Wei, D.; Zhang, M.; Qin, J. Chemical Components and Biological Activities of the Essential Oil from Traditional Medicinal Food, Euryale ferox Salisb., Seeds. J. Essent. Oil-Bear. Plants 2019, 22, 73–81. [Google Scholar] [CrossRef]
- Song, C.W.; Wang, S.M.; Zhou, L.L.; Hou, F.F.; Wang, K.J.; Han, Q.B.; Li, N.; Cheng, Y.X. Isolation and identification of compounds responsible for antioxidant capacity of euryale ferox seeds. J. Agric. Food Chem. 2011, 59, 1199–1204. [Google Scholar] [CrossRef]
- Chen, G.; Chen, B.; Song, D. Co-microbiological regulation of phenolic release through solid-state fermentation of corn kernels (Zea mays L.) to improve their antioxidant activity. LWT 2021, 142, 111003. [Google Scholar] [CrossRef]
- Dordević, T.M.; Šiler-Marinković, S.S.; Dimitrijević-Branković, S.I. Effect of fermentation on antioxidant properties of some cereals and pseudo cereals. Food Chem. 2010, 119, 957–963. [Google Scholar] [CrossRef]
- Degli, U.; Di, S. Lactic Acid Fermentation: A Traditional Process for New Applications. Ph.D. Thesis, University of Parma, Parma, Italy, 2018. [Google Scholar]
- Lee, D.E.; Huh, C.S.; Ra, J.; Choi, I.D.; Jeong, J.W.; Kim, S.H.; Ryu, J.H.; Seo, Y.K.; Koh, J.S.; Lee, J.H.; et al. Clinical evidence of effects of Lactobacillus plantarum HY7714 on skin aging: A randomized, double blind, placebo-controlled study. J. Microbiol. Biotechnol. 2015, 25, 2160–2168. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, W.; Wei, Z.; Yin, B.; Man, C.; Jiang, Y. Enhancement of functional characteristics of blueberry juice fermented by Lactobacillus plantarum. LWT 2021, 139, 110590. [Google Scholar] [CrossRef]
- Bhat, R.; Suryanarayana, L.C.; Chandrashekara, K.A.; Krishnan, P.; Kush, A.; Ravikumar, P. Lactobacillus plantarum mediated fermentation of Psidium guajava L. fruit extract. J. Biosci. Bioeng. 2015, 119, 430–432. [Google Scholar] [CrossRef] [PubMed]
- Hur, S.J.; Lee, S.Y.; Kim, Y.C.; Choi, I.; Kim, G.B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, L.; Qiao, N.; Xiao, Y.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W.; Zhai, Q. Latilactobacillus curvatus: A candidate probiotic with excellent fermentation properties and health benefits. Foods 2020, 9, 1366. [Google Scholar] [CrossRef] [PubMed]
- Kaprasob, R.; Kerdchoechuen, O.; Laohakunjit, N.; Somboonpanyakul, P. B vitamins and prebiotic fructooligosaccharides of cashew apple fermented with probiotic strains Lactobacillus spp., Leuconostoc mesenteroides and Bifidobacterium longum. Process Biochem. 2018, 70, 9–19. [Google Scholar] [CrossRef]
- Chen, G.; Liu, Y.; Zeng, J.; Tian, X.; Bei, Q.; Wu, Z. Enhancing three phenolic fractions of oats (Avena sativa L.) and their antioxidant activities by solid-state fermentation with Monascus anka and Bacillus subtilis. J. Cereal Sci. 2020, 93, 102940. [Google Scholar] [CrossRef]
- Dulf, F.V.; Vodnar, D.C.; Socaciu, C. Effects of solid-state fermentation with two filamentous fungi on the total phenolic contents, flavonoids, antioxidant activities and lipid fractions of plum fruit (Prunus domestica L.) by-products. Food Chem. 2016, 209, 27–36. [Google Scholar] [CrossRef]
- Zhang, Y.; You, S.; Wang, D.; Zhao, D.; Zhang, J.; An, Q.; Li, M.; Wang, C. Fermented Dendrobium officinale polysaccharides protect UVA-induced photoaging of human skin fibroblasts. Food Sci. Nutr. 2022, 10, 1275–1288. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, Y.; Huang, G. Preparation and antioxidant activities of important traditional plant polysaccharides. Int. J. Biol. Macromol. 2018, 111, 780–786. [Google Scholar] [CrossRef]
- Picton, L.; Bataille, I.; Muller, G. Analysis of a complex polysaccharide (gum arabic) by multi-angle laser light scattering coupled on-line to size exclusion chromatography and flow field flow fractionation. Carbohydr. Polym. 2000, 42, 23–31. [Google Scholar] [CrossRef]
- Buranov, A.U.; Mazza, G. Extraction and characterization of hemicelluloses from flax shives by different methods. Carbohydr. Polym. 2010, 79, 17–25. [Google Scholar] [CrossRef]
- Liu, J.; Meng, C.G.; Yan, Y.H.; Shan, Y.N.; Kan, J.; Jin, C.H. Structure, physical property and antioxidant activity of catechin grafted Tremella fuciformis polysaccharide. Int. J. Biol. Macromol. 2016, 82, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.H.; Xie, M.Y.; Nie, S.P.; Shen, M.Y.; Wang, Y.X.; Li, C. Isolation, chemical composition and antioxidant activities of a water-soluble polysaccharide from Cyclocarya paliurus (Batal.) Iljinskaja. Food Chem. 2010, 119, 1626–1632. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, R.; Zhang, Y.; Yang, Y.; Sun, X.; Zhang, Q.; Yang, N. Biotransformation of phenolics and metabolites and the change in antioxidant activity in kiwifruit induced by Lactobacillus plantarum fermentation. J. Sci. Food Agric. 2020, 100, 3283–3290. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, D.; Fu, H.; Zhao, D.; Zhang, J.; Li, M.; Wang, C. Protective effects of extracellular proteins of Saccharomycopsis fibuligera on UVA-damaged human skin fibroblasts. J. Funct. Foods 2022, 88, 104897. [Google Scholar] [CrossRef]
- Dong, L.I.; Yun, W.U. Research progress on the main components of Euryale ferox and predictive analysis on Q—Marker. Grain Oil 2021. Available online: https://www.cnki.com.cn/Article/CJFDTotal-LSYY202109008.htm (accessed on 25 March 2022).
- Kim, Y.; Goodner, K.L.; Park, J.D.; Choi, J.; Talcott, S.T. Changes in antioxidant phytochemicals and volatile composition of Camellia sinensis by oxidation during tea fermentation. Food Chem. 2011, 129, 1331–1342. [Google Scholar] [CrossRef]
- Michlmayr, H.; Kneifel, W. β-Glucosidase activities of lactic acid bacteria: Mechanisms, impact on fermented food and human health. FEMS Microbiol. Lett. 2014, 352, 1–10. [Google Scholar] [CrossRef]
- Lee, J.; Koo, N.; Min, D.B. Reactive Oxygen Species, Aging, and Antioxidative Nutraceuticals. Compr. Rev. Food Sci. Food Saf. 2004, 3, 21–33. [Google Scholar] [CrossRef]
- Ajibola, C.F.; Fashakin, J.B.; Fagbemi, T.N.; Aluko, R.E. Effect of peptide size on antioxidant properties of African yam bean seed (Sphenostylis stenocarpa) protein hydrolysate fractions. Int. J. Mol. Sci. 2011, 12, 6685–6702. [Google Scholar] [CrossRef]
- Zhu, K.; Zhou, H.; Qian, H. Antioxidant and free radical-scavenging activities of wheat germ protein hydrolysates (WGPH) prepared with alcalase. Process Biochem. 2006, 41, 1296–1302. [Google Scholar] [CrossRef]
- Girgih, A.T.; Udenigwe, C.C.; Aluko, R.E. In Vitro Antioxidant Properties of Hemp Seed (Cannabis sativa L.) Protein Hydrolysate Fractions. J. Am. Oil Chem. Soc. 2010 883 2010, 88, 381–389. [Google Scholar] [CrossRef]
- Wang, C. Euryale ferox 芡实 (Qianshi, Gordon Euryale Seed). In Dietary Chinese Herbs; Springer: Berlin/Heidelberg, Germany, 2015; pp. 371–377. [Google Scholar] [CrossRef]
- Guo, G.; Kong, Y.; Su, J.; Wang, G.; Zhang, M.; Wang, S.; Song, Z. Immunomodulatory activity of aqueous extract from Crassostrea sikamea in the splenocytes of Sprague-Dawley rats. Food Sci. Nutr. 2022, 10, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Qi, J.; Du, D.; Liu, Y.; Jiang, X. Current advances of Dendrobium officinale polysaccharides in dermatology: A literature review. Pharm. Biol. 2020, 58, 664–673. [Google Scholar] [CrossRef]
- Guo, R.; Guo, S.; Gao, X.; Wang, H.; Hu, W.; Duan, R.; Dong, T.T.X.; Tsim, K.W.K. Fermentation of Danggui Buxue Tang, an ancient Chinese herbal mixture, together with Lactobacillus plantarum enhances the anti-diabetic functions of herbal product. Chinese Med. 2020, 15, 98. [Google Scholar] [CrossRef]
- Cai, L.; Kang, Y.J. Oxidative stress and diabetic cardiomyopathy. Cardiovasc. Toxicol. 2001, 1, 181–193. [Google Scholar] [CrossRef]
- Shahbazi, R.; Sharifzad, F.; Bagheri, R.; Alsadi, N.; Yasavoli-Sharahi, H.; Matar, C. Anti-inflammatory and immunomodulatory properties of fermented plant foods. Nutrients 2021, 13, 1516. [Google Scholar] [CrossRef]
- Kongpetch, S.; Kukongviriyapan, V.; Prawan, A.; Senggunprai, L.; Kukongviriyapan, U.; Buranrat, B. Crucial Role of Heme Oxygenase-1 on the Sensitivity of Cholangiocarcinoma Cells to Chemotherapeutic Agents. PLoS ONE 2012, 7, e34994. [Google Scholar] [CrossRef]
- Espinosa-Díez, C.; Miguel, V.; Vallejo, S.; Sánchez, F.J.; Sandoval, E.; Blanco, E.; Cannata, P.; Peiró, C.; Sánchez-Ferrer, C.F.; Lamas, S. Role of glutathione biosynthesis in endothelial dysfunction and fibrosis. Redox Biol. 2018, 14, 88–99. [Google Scholar] [CrossRef]
- Kwok, H.H.; Ng, W.Y.; Yang, M.S.M.; Mak, N.K.; Wong, R.N.S.; Yue, P.Y.K. The ginsenoside protopanaxatriol protects endothelial cells from hydrogen peroxide-induced cell injury and cell death by modulating intracellular redox status. Free Radic. Biol. Med. 2010, 48, 437–445. [Google Scholar] [CrossRef]
- Toutfaire, M.; Bauwens, E.; Debacq-Chainiaux, F. The impact of cellular senescence in skin ageing: A notion of mosaic and therapeutic strategies. Biochem. Pharmacol. 2017, 142, 1–12. [Google Scholar] [CrossRef]
- Watson, R.E.B.; Gibbs, N.K.; Griffiths, C.E.M.; Sherratt, M.J. Damage to Skin Extracellular Matrix Induced by UV Exposure. Antioxid. Redox Signal. 2014, 21, 1063–1077. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M. Plant-Derived Antioxidants: Significance in Skin Health and the Ageing Process. Int. J. Mol. Sci. 2022, 23, 585. [Google Scholar] [CrossRef]
- Mizutani, T.; Masaki, H. Anti-photoaging capability of antioxidant extract from Camellia japonica leaf. Exp. Dermatol. 2014, 23, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lu, Y.R.; Lin, I.F.; Kang, W.; Chen, H.B.; Lu, H.F.; Wang, H.M.D. Reversing UVB-induced photoaging with Hibiscus sabdariffa calyx aqueous extract. J. Sci. Food Agric. 2020, 100, 672–681. [Google Scholar] [CrossRef]
- Jiang, T.; Huang, Z.; Lin, Y.; Zhang, Z.; Fang, D.; Zhang, D.D. The Protective Role of Nrf2 in Streptozotocin-Induced Diabetic Nephropathy. Diabetes 2010, 59, 850–860. [Google Scholar] [CrossRef]
- Taguchi, K.; Motohashi, H.; Yamamoto, M. Molecular mechanisms of the Keap1–Nrf2 pathway in stress response and cancer evolution. Genes Cells 2011, 16, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Eizirik, D.L.; Flodström, M.; Karlsen, A.E.; Welsh, N. The harmony of the spheres: Inducible nitric oxide synthase and related genes in pancreatic beta cells. Diabetologia 1996, 39, 875–890. [Google Scholar] [CrossRef]
- Zhu, H.; Jia, Z.; Misra, B.R.; Zhang, L.; Cao, Z.; Yamamoto, M.; Trush, M.A.; Misra, H.P.; Li, Y. Nuclear Factor E2-Related Factor 2-Dependent Myocardiac Cytoprotection Against Oxidative and Electrophilic Stress. Cardiovasc. Toxicol. 2008, 8, 71–85. [Google Scholar] [CrossRef]
- Dayalan Naidu, S.; Kostov, R.V.; Dinkova-Kostova, A.T. Transcription factors Hsf1 and Nrf2 engage in crosstalk for cytoprotection. Trends Pharmacol. Sci. 2015, 36, 6–14. [Google Scholar] [CrossRef]
- Sakai, N.; Wada, T.; Furuichi, K.; Iwata, Y.; Yoshimoto, K.; Kitagawa, K.; Kokubo, S.; Kobayashi, M.; Hara, A.; Yamahana, J.; et al. Involvement of extracellular signal-regulated kinase and p38 in human diabetic nephropathy. Am. J. Kidney Dis. 2005, 45, 54–65. [Google Scholar] [CrossRef]
- Gupta, J.; Nebreda, A.R. Roles of p38α mitogen-activated protein kinase in mouse models of inflammatory diseases and cancer. FEBS J. 2015, 282, 1841–1857. [Google Scholar] [CrossRef]
- Bonni, A.; Brunet, A.; West, A.E.; Datta, S.R.; Takasu, M.A.; Greenberg, M.E. Cell Survival Promoted by the Ras-MAPK Signaling Pathway by Transcription-Dependent and -Independent Mechanisms. Science 1999, 286, 1358–1362. [Google Scholar] [CrossRef]
- Ma, L.; Wu, F.; Shao, Q.; Chen, G.; Xu, L.; Lu, F. Baicalin Alleviates Oxidative Stress and Inflammation in Diabetic Nephropathy via Nrf2 and MAPK Signaling Pathway. Drug Des. Devel. Ther. 2021, 15, 3207. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioactive Components | Fermentation System (mg/mL) | |
|---|---|---|
| UF | LCF | |
| Total phenolic content | 0.041 ± 0.000 a | 0.052 ± 0.015 b |
| Total protein content | 4.320 ± 0.700 a | 7.463 ± 0.261 a |
| Total flavonoid content | 1.358 ± 0.167 a | 1.402 ± 0.000 b |
| Total polypeptide content | 0.399 ± 0.009 a | 0.627 ± 0.012 a |
| Total polysaccharide content | 1.657 ± 0.088 a | 2.101 ± 0.000 b |
| Sample Name | Peak Limits (min) | Area (%) | Molecular Weight (Da) |
|---|---|---|---|
| UF | 5.10 | 7.21 | 350,018.87 |
| 10.13 | 92.79 | 2539.43 | |
| LCF | 5.10 | 10.24 | 349,156.22 |
| 9.92 | 30.37 | 3123.51 | |
| 10.10 10.59 | 23.17 36.23 | 2608.02 1625.89 |
| UF | LCF | |||
|---|---|---|---|---|
| Peak Name | Peak 1 | Peak 2 | Peak 1 | Peak 2 |
| Peak Limits (min) | 13.131–19.579 | 24.210–26.526 | 13.629–20.683 | 24.887–26.312 |
| Mw | 1.617 × 106 (±1.946%) | 5.099 × 104 (±20.538%) | 1.088 × 106 (±1.088%) | 4.306 × 103 (±13.482%) |
| Mz | 1.139 × 107 (±8.631%) | 1.612 × 105 (±47.918%) | 4.140 × 106 (±2.591%) | 3.382 × 104 (±40.654%) |
| Mw/Mn | 1.809 (±2.285%) | 2.259 (±26.791%) | 1.631 (±1.391%) | 1.851 (±17.937%) |
| Mz/Mn | 12.741 (±8.714%) | 7.143 (±50.912%) | 6.204 (±2.732%) | 14.536 (±42.340%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; You, S.; Zhang, Y.; Zhao, J.; Wang, D.; Zhao, D.; Li, M.; Wang, C. Enhancing Bioactive Components of Euryale ferox with Lactobacillus curvatus to Reduce H2O2-Induced Oxidative Stress in Human Skin Fibroblasts. Antioxidants 2022, 11, 1881. https://doi.org/10.3390/antiox11101881
Jiang Y, You S, Zhang Y, Zhao J, Wang D, Zhao D, Li M, Wang C. Enhancing Bioactive Components of Euryale ferox with Lactobacillus curvatus to Reduce H2O2-Induced Oxidative Stress in Human Skin Fibroblasts. Antioxidants. 2022; 11(10):1881. https://doi.org/10.3390/antiox11101881
Chicago/Turabian StyleJiang, Yanbing, Shiquan You, Yongtao Zhang, Jingsha Zhao, Dongdong Wang, Dan Zhao, Meng Li, and Changtao Wang. 2022. "Enhancing Bioactive Components of Euryale ferox with Lactobacillus curvatus to Reduce H2O2-Induced Oxidative Stress in Human Skin Fibroblasts" Antioxidants 11, no. 10: 1881. https://doi.org/10.3390/antiox11101881
APA StyleJiang, Y., You, S., Zhang, Y., Zhao, J., Wang, D., Zhao, D., Li, M., & Wang, C. (2022). Enhancing Bioactive Components of Euryale ferox with Lactobacillus curvatus to Reduce H2O2-Induced Oxidative Stress in Human Skin Fibroblasts. Antioxidants, 11(10), 1881. https://doi.org/10.3390/antiox11101881