
Glucose Starvation-Caused Oxidative Stress Induces Inflammation and Autophagy in Human Gingival Fibroblasts

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Cell Culture
2.3. Detection of Cell Proliferation
2.4. Cell Survival Assay
2.5. Immunofluorescence Staining
2.6. ROS Detection and Staining
2.7. Real-Time Polymerize Chain Reaction
2.8. Human HO-1 and SOD ELISA
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
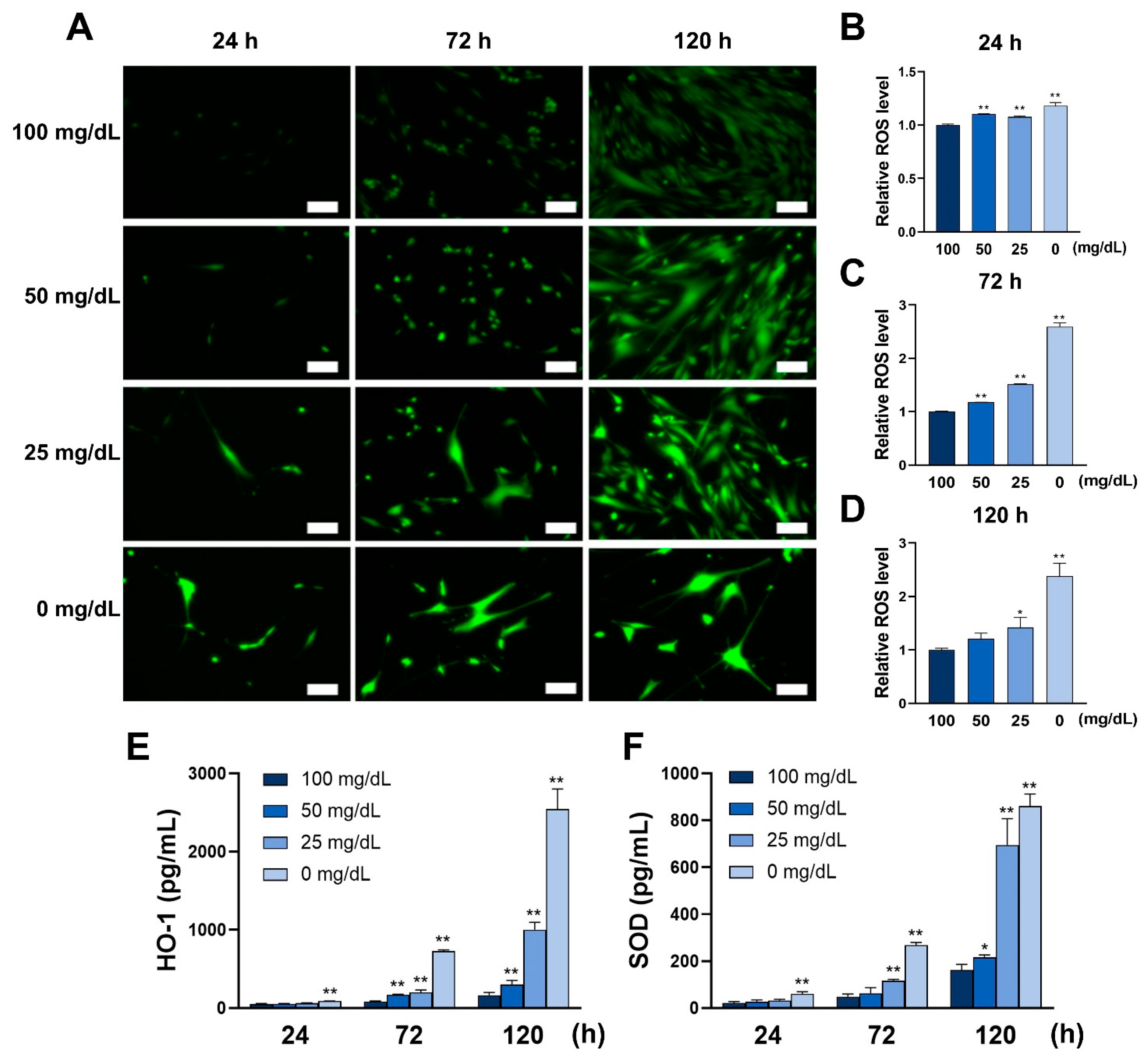
3.1. Low Glucose Conditions Increase ROS Levels
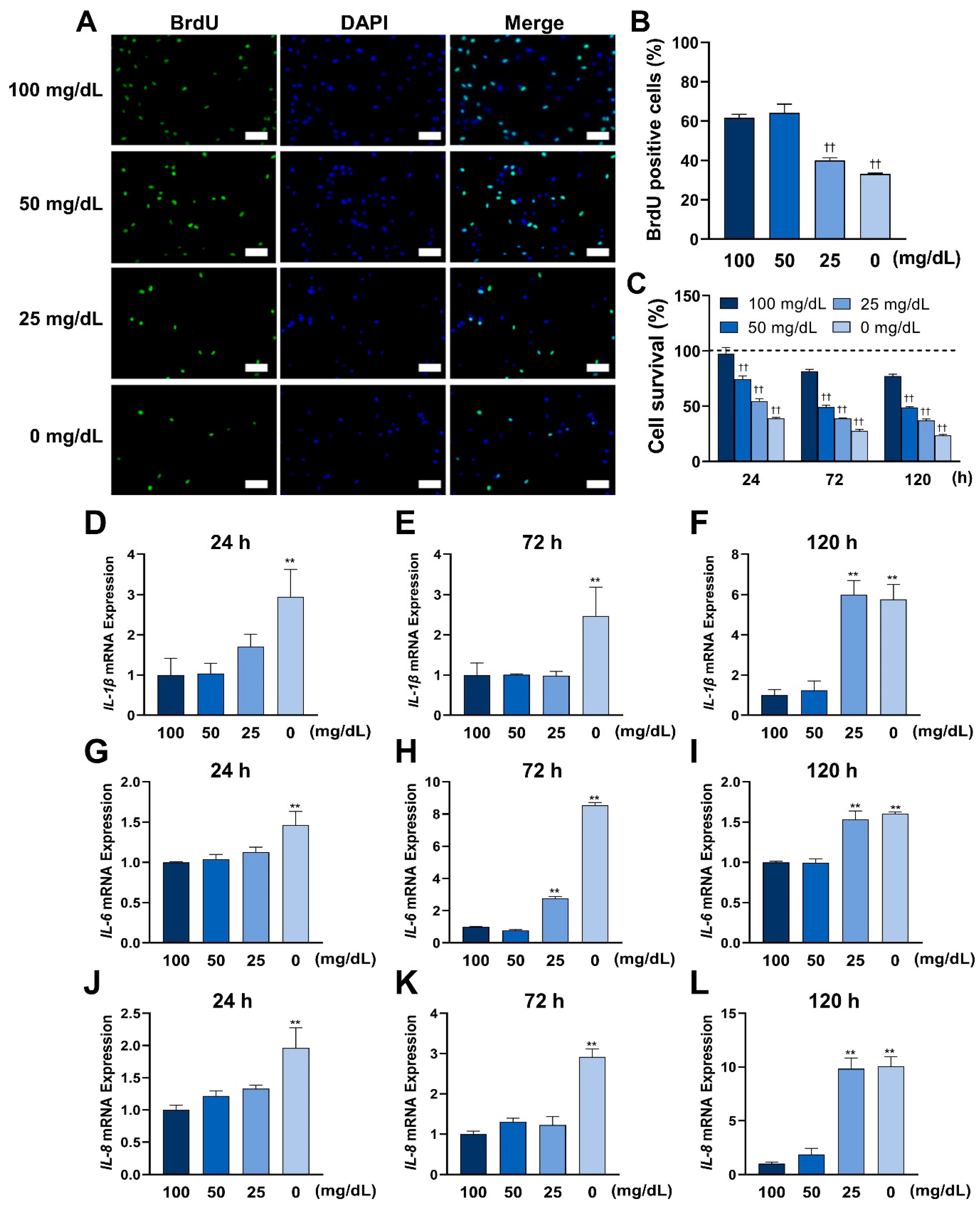
3.2. Low Glucose Conditions Impair Cellular Viability and Promote the Expression of Inflammatory Cytokines
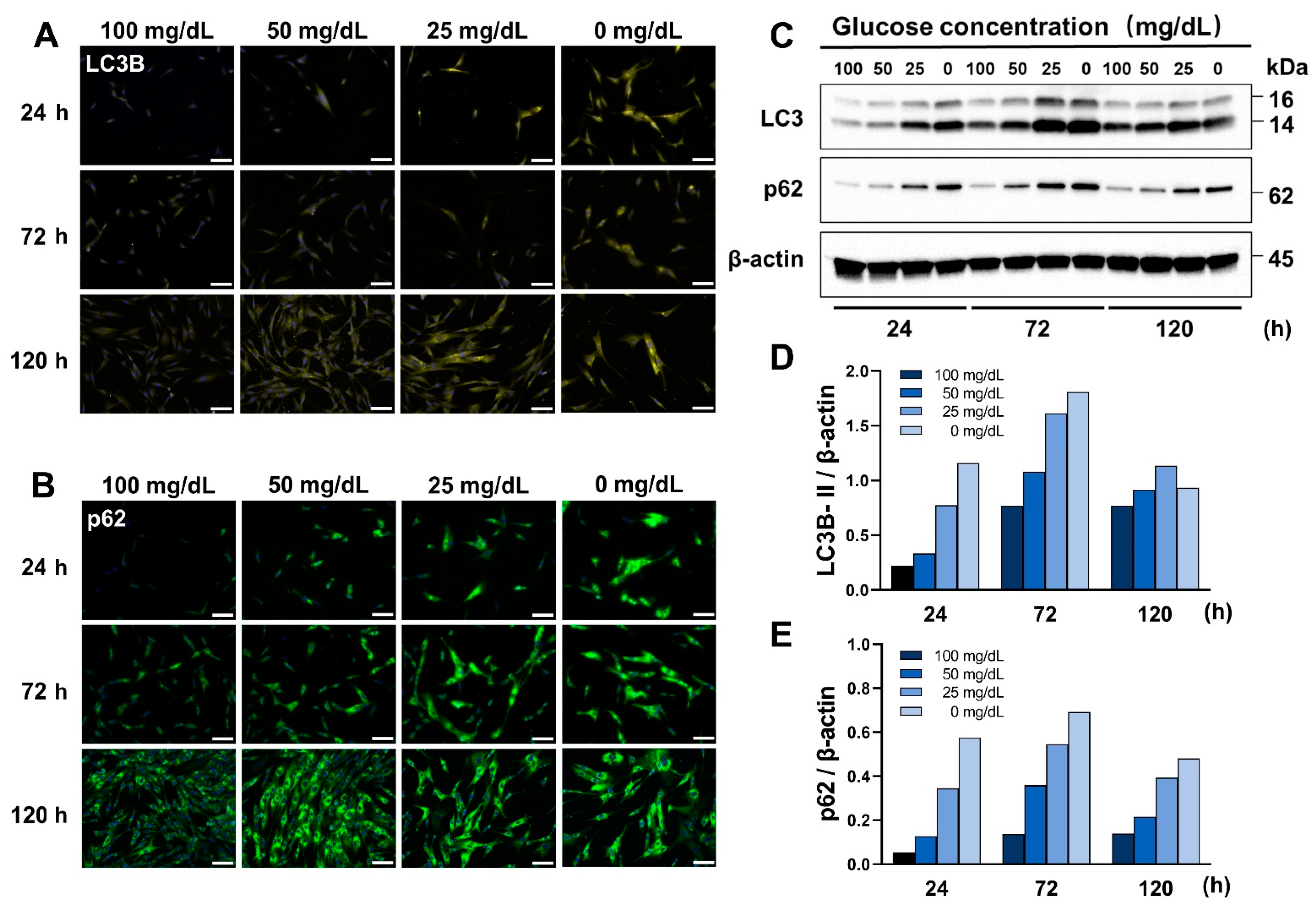
3.3. Low Glucose Conditions Induce Autophagy
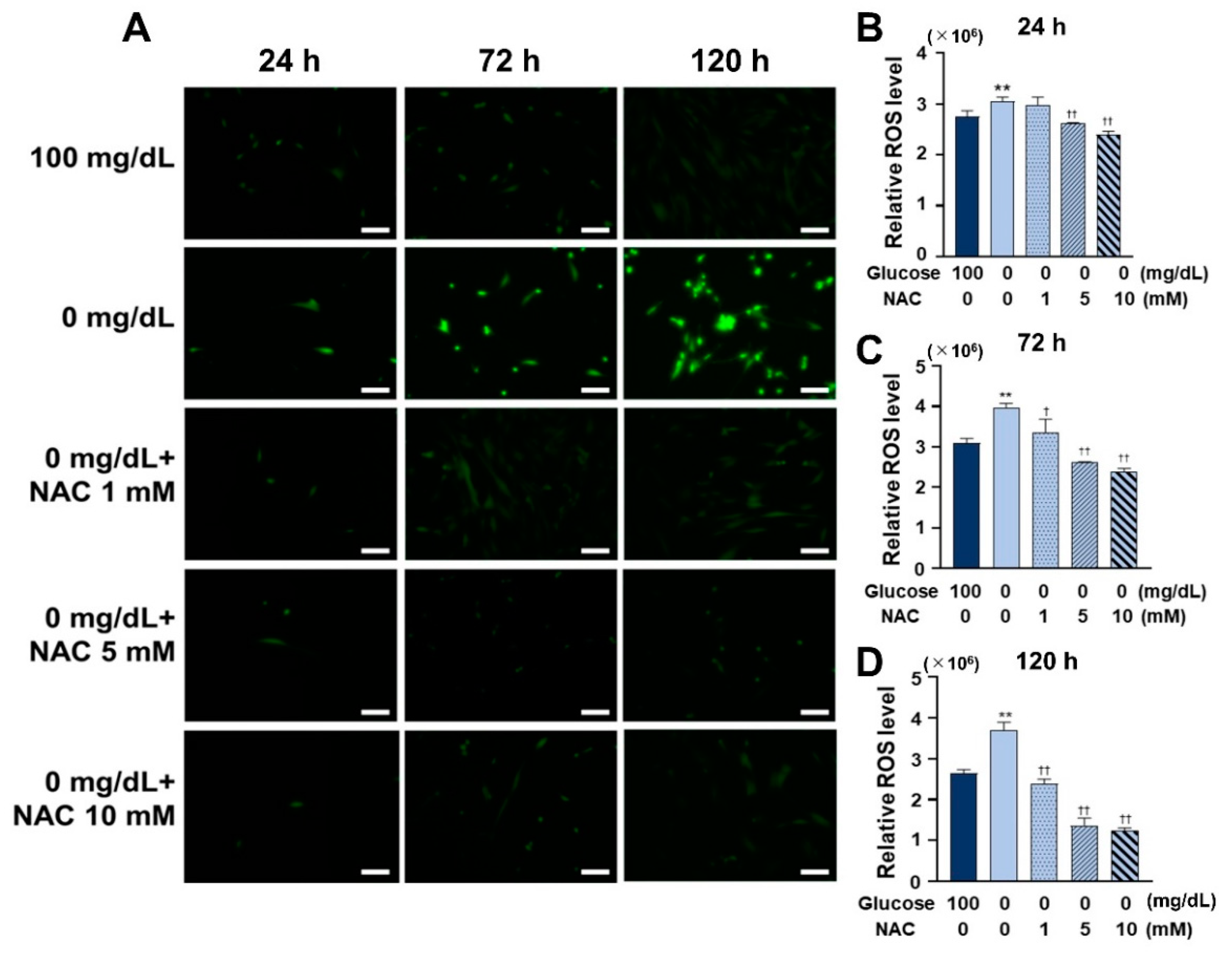
3.4. NAC Administration Inhibited ROS Formation
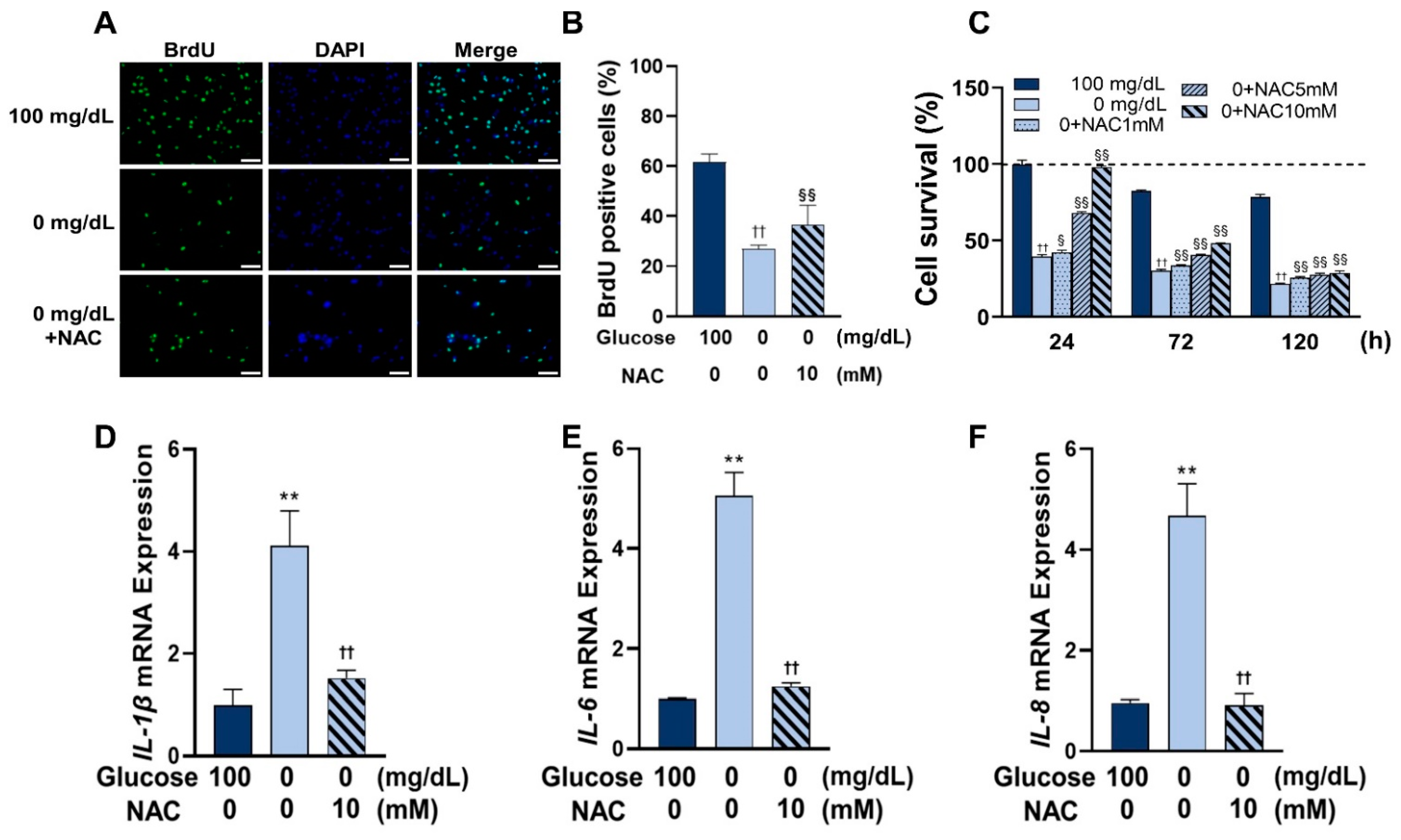
3.5. Administration of NAC Rescued Cell Viability and Inhibited the Expression of Inflammatory Cytokines under Glucose Starvation Conditions
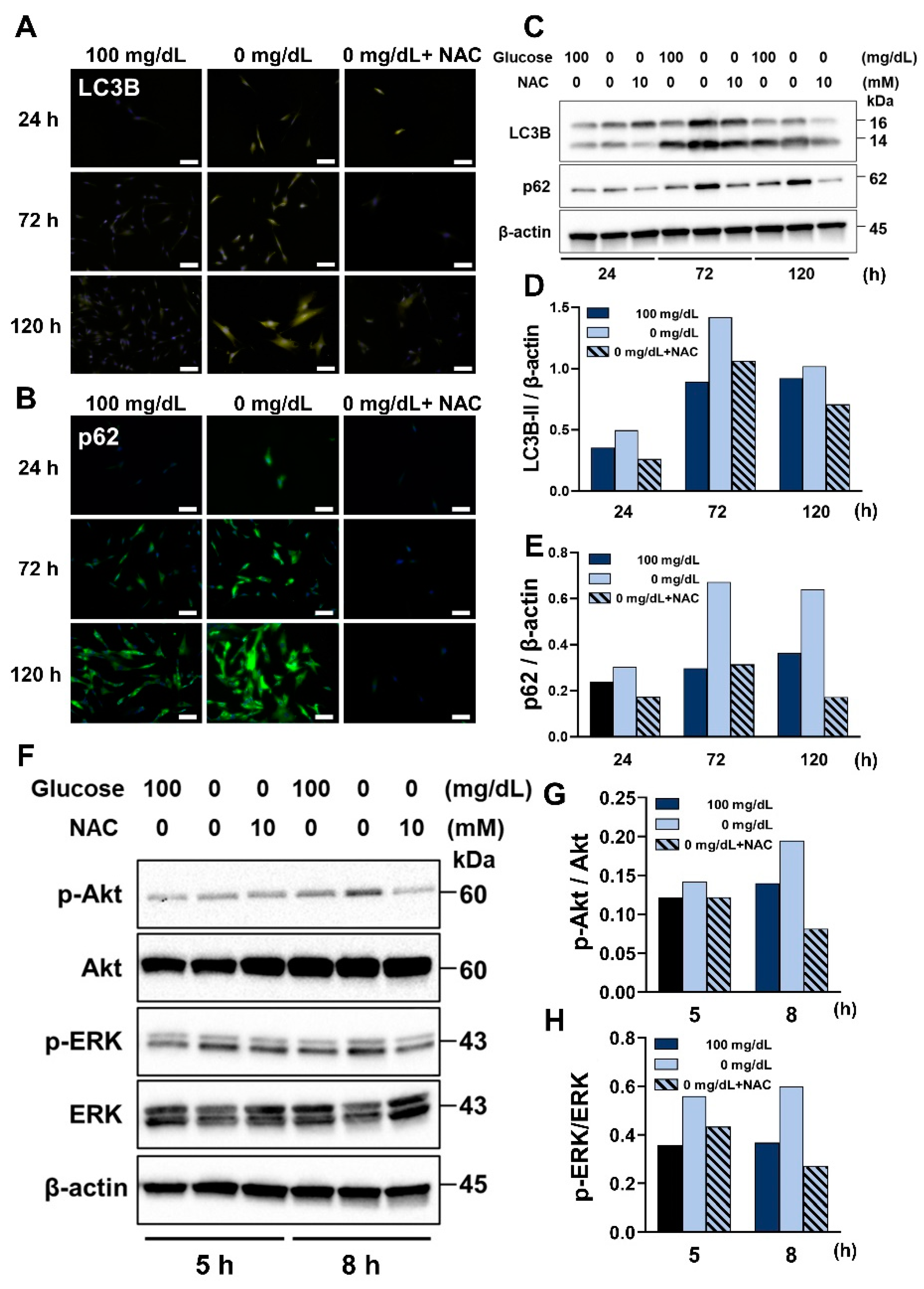
3.6. NAC Treatment Suppressed the Process of Autophagy and the ROS-Akt/ERK Signaling Pathway under Glucose Starvation Conditions
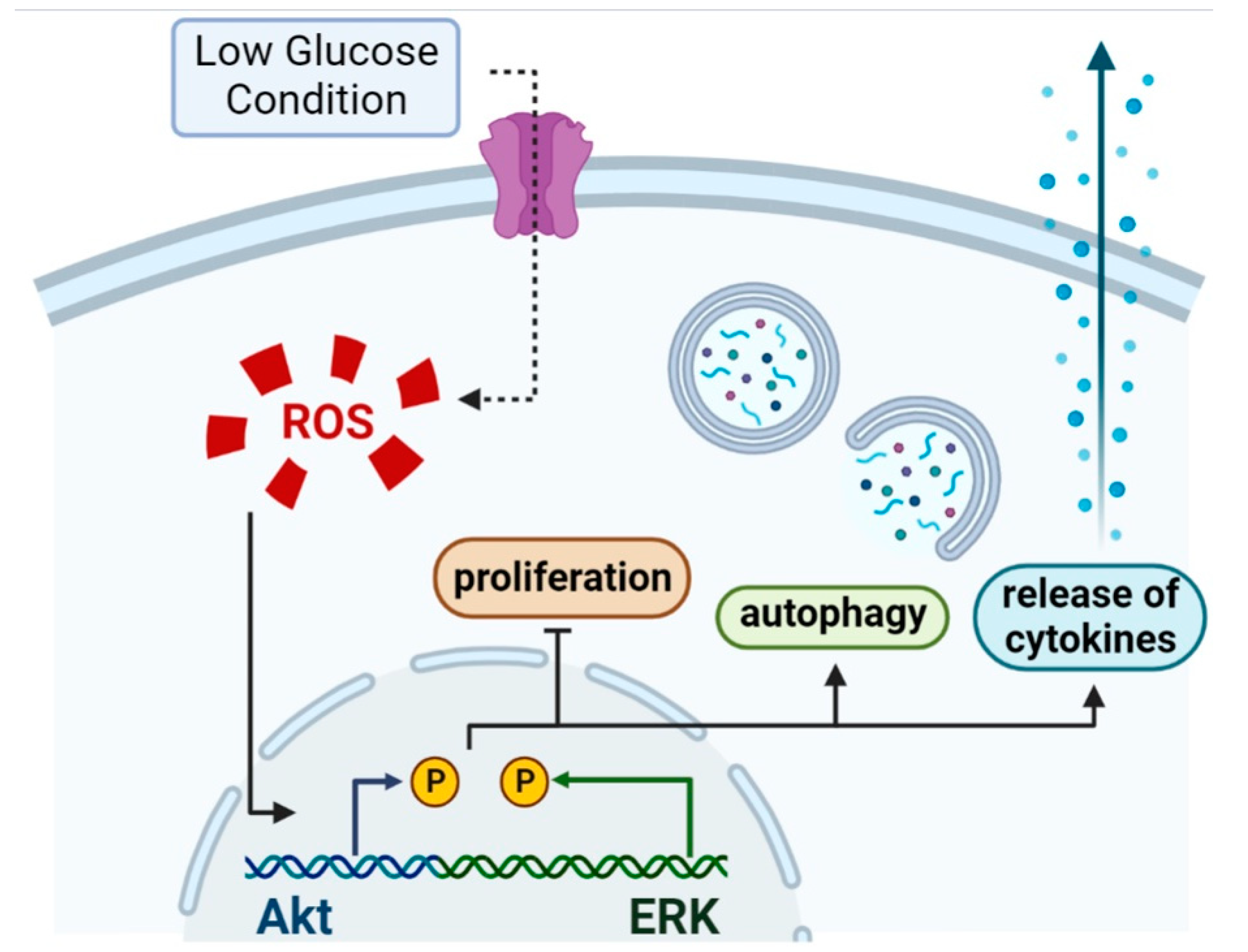
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galant, A.L.; Kaufman, R.C.; Wilson, J.D. Glucose: Detection and Analysis. Food Chem. 2015, 188, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Verdonk, C.A.; Rizza, R.A.; Gerich, J.E. Effects of Plasma Glucose Concentration on Glucose Utilization and Glucose Clearance in Normal Man. Diabetes 1981, 30, 535–537. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, L.; Giaccari, A.; DeFronzo, R.A. Glucose Toxicity. Diabetes Care 1990, 13, 610–630. [Google Scholar] [CrossRef] [PubMed]
- Wolff, S.P.; Dean, R.T. Glucose Autoxidation and Protein Modification. The Potential Role of “autoxidative Glycosylation” in Diabetes. Biochem. J. 1987, 245, 243–250. [Google Scholar] [CrossRef]
- Lee, T.S.; Saltsman, K.A.; Ohashi, H.; King, G.L. Activation of Protein Kinase C by Elevation of Glucose Concentration: Proposal for a Mechanism in the Development of Diabetic Vascular Complications. Proc. Natl. Acad. Sci. USA 1989, 86, 5141–5145. [Google Scholar] [CrossRef]
- Rosca, M.G.; Mustata, T.G.; Kinter, M.T.; Ozdemir, A.M.; Kern, T.S.; Szweda, L.I.; Brownlee, M.; Monnier, V.M.; Weiss, M.F. Glycation of Mitochondrial Proteins from Diabetic Rat Kidney Is Associated with Excess Superoxide Formation. Am. J. Physiol.-Ren. Physiol. 2005, 289, F420–F430. [Google Scholar] [CrossRef]
- Yang, H.; Jin, X.; Lam, C.W.K.; Yan, S.K. Oxidative Stress and Diabetes Mellitus. Clin. Chem. Lab. Med. 2011, 49, 1773–1782. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef]
- Li, R.; Kato, H.; Taguchi, Y.; Umeda, M. Intracellular Glucose Starvation Affects Gingival Homeostasis and Autophagy. Sci. Rep. 2022, 12, 1–10. [Google Scholar] [CrossRef]
- Dent, P. Crosstalk between ERK, AKT, and Cell Survival. Cancer Biol. Ther. 2014, 15, 245–246. [Google Scholar] [CrossRef] [Green Version]
- Rabinowitz, J.D.; White, E. Autophagy and Metabolism. Science 2010, 330, 1344–1348. [Google Scholar] [CrossRef] [PubMed]
- Bullon, P.; Cordero, M.D.; Quiles, J.L.; Ramirez-Tortosa, M.D.C.; Gonzalez-Alonso, A.; Alfonsi, S.; García-Marín, R.; de Miguel, M.; Battino, M. Autophagy in Periodontitis Patients and Gingival Fibroblasts: Unraveling the Link between Chronic Diseases and Inflammation. BMC Med. 2012, 10, 122. [Google Scholar] [CrossRef] [PubMed]
- Greabu, M.; Giampieri, F.; Melescanu Imre, M.; Mohora, M.; Totan, A.; Pituru, S.M.; Ionescu, E. Autophagy, One of the Main Steps in Periodontitis Pathogenesis and Evolution. Molecules 2020, 25, 4338. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.J.; Park, S.Y.; Kim, O.S.; Park, H.S.; Jung, J.Y. Autophagy Upregulates Inflammatory Cytokines in Gingival Tissue of Patients with Periodontitis and Lipopolysaccharide-Stimulated Human Gingival Fibroblasts. J. Periodontol. 2022, 93, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa-Ishimoto, Y.; Hwang, S.; Cadwell, K. Autophagy and Inflammation. Annu. Rev. Immunol. 2018, 36, 73–101. [Google Scholar] [CrossRef] [PubMed]
- Buranasin, P.; Mizutani, K.; Iwasaki, K.; Mahasarakham, C.P.N.; Kido, D.; Takeda, K.; Izumi, Y. High Glucose-Induced Oxidative Stress Impairs Proliferation and Migration of Human Gingival Fibroblasts. PLoS ONE 2018, 13, e0201855. [Google Scholar] [CrossRef]
- Zheng, Y.; Liu, Y.; Ge, J.; Wang, X.; Liu, L.; Bu, Z.; Liu, P. Resveratrol Protects Human Lens Epithelial Cells against H2O2- Induced Oxidative Stress by Increasing Catalase, SOD-1, and HO-1 Expression. Mol. Vis. 2010, 16, 1467–1474. [Google Scholar]
- Yu, T.; Jhun, B.S.; Yoon, Y. High-Glucose Stimulation Increases Reactive Oxygen Species Production through the Calcium and Mitogen-Activated Protein Kinase-Mediated Activation of Mitochondrial Fission. Antioxid. Redox Signal. 2011, 14, 425–437. [Google Scholar] [CrossRef]
- Kashiwagi, Y.; Takedachi, M.; Mori, K.; Kubota, M.; Yamada, S.; Kitamura, M.; Murakami, S. High Glucose-Induced Oxidative Stress Increases IL-8 Production in Human Gingival Epithelial Cells. Oral Dis. 2016, 22, 578–584. [Google Scholar] [CrossRef]
- Haekkinen, L.A.R.I.; Uitto, V.J.; Larjava, H. Cell Biology of Gingival Wound Healing. Periodontol 2000, 24, 127–152. [Google Scholar]
- Young, A.; McNaught, C.E. The Physiology of Wound Healing. Surgery 2011, 29, 475–479. [Google Scholar] [CrossRef]
- Kato, H.; Taguchi, Y.; Tominaga, K.; Kimura, D.; Yamawaki, I.; Noguchi, M.; Yamauchi, N.; Tamura, I.; Tanaka, A.; Umeda, M. High Glucose Concentrations Suppress the Proliferation of Human Periodontal Ligament Stem Cells and Their Differentiation Into Osteoblasts. J. Periodontol. 2016, 87, e44–e51. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N. Autophagy: Process and Function. Genes Dev. 2007, 21, 2861–2873. [Google Scholar] [CrossRef]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of Metabolism and Mitochondrial Homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Scherz-Shouval, R.; Elazar, Z. ROS, Mitochondria and the Regulation of Autophagy. Trends Cell Biol. 2007, 17, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N-Acetylcysteine as an Antioxidant and Disulphide Breaking Agent: The Reasons Why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar] [CrossRef]
- Kelly, G.S. Clinical Applications of N-Acetylcysteine. Altern. Med. Rev. 1998, 3, 114–127. [Google Scholar]
- Wang, C.; Chen, K.; Xia, Y.; Dai, W.; Wang, F.; Shen, M.; Cheng, P.; Wang, J.; Lu, J.; Zhang, Y.; et al. N-Acetylcysteine Attenuates Ischemia-Reperfusion-Induced Apoptosis and Autophagy in Mouse Liver via Regulation of the ROS/JNK/Bcl-2 Pathway. PLoS ONE 2014, 9, e108855. [Google Scholar] [CrossRef]
- Tanida, I. Autophagy Basics. Microbiol. Immunol. 2011, 55, 1–11. [Google Scholar] [CrossRef]
- Martinez-Lopez, N.; Athonvarangkul, D.; Mishall, P.; Sahu, S.; Singh, R. Autophagy Proteins Regulate ERK Phosphorylation. Nat. Commun. 2013, 4, 2799. [Google Scholar] [CrossRef]
- Corcelle, E.; Djerbi, N.; Mari, M.; Nebout, M.; Fiorini, C.; Fénichel, P.; Hofman, P.; Poujeol, P.; Mograbi, B. Control of the Autophagy Maturation Step by the MAPK ERK and P38: Lessons from Environmental Carcinogens. Autophagy 2007, 3, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Hu, X.; Liu, Y.; Dong, S.; Wen, Z.; He, W.; Zhang, S.; Huang, Q.; Shi, M. ROS Signaling under Metabolic Stress: Cross-Talk between AMPK and AKT Pathway. Mol. Cancer 2017, 16, 79. [Google Scholar] [CrossRef] [PubMed]
- Polytarchou, C.; Hatziapostolou, M.; Yau, T.O.; Christodoulou, N.; Hinds, P.W.; Kottakis, F.; Sanidas, I.; Tsichlis, P.N. Akt3 Induces Oxidative Stress and DNA Damage by Activating the NADPH Oxidase via Phosphorylation of P47phox. Proc. Natl. Acad. Sci. USA 2020, 117, 28806–28815. [Google Scholar] [CrossRef] [PubMed]
- Kominato, H.; Takeda, K.; Mizutani, K.; Mikami, R.; Kido, D.; Buranasin, P.; Saito, N.; Takemura, S.; Nakagawa, K.; Nagasawa, T.; et al. Metformin Accelerates Wound Healing by Akt Phosphorylation of Gingival Fibroblasts in Insulin-Resistant Prediabetes Mice. J. Periodontol. 2022, 93, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Dibart, S. Free Gingival Autograft. Pract. Periodontal Plast. Surg. 2017, 21–27. [Google Scholar] [CrossRef]
- Orihuela-Campos, R.C.; Tamaki, N.; Mukai, R.; Fukui, M.; Miki, K.; Terao, J.; Ito, H.-O. Biological Impacts of Resveratrol, Quercetin, and N-Acetylcysteine on Oxidative Stress in Human Gingival Fibroblasts. J. Clin. Biochem. Nutr. 2015, 56, 220–227. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, R.; Kato, H.; Taguchi, Y.; Deng, X.; Minagawa, E.; Nakata, T.; Umeda, M. Glucose Starvation-Caused Oxidative Stress Induces Inflammation and Autophagy in Human Gingival Fibroblasts. Antioxidants 2022, 11, 1907. https://doi.org/10.3390/antiox11101907
Li R, Kato H, Taguchi Y, Deng X, Minagawa E, Nakata T, Umeda M. Glucose Starvation-Caused Oxidative Stress Induces Inflammation and Autophagy in Human Gingival Fibroblasts. Antioxidants. 2022; 11(10):1907. https://doi.org/10.3390/antiox11101907
Chicago/Turabian StyleLi, Runbo, Hirohito Kato, Yoichiro Taguchi, Xin Deng, Emika Minagawa, Takaya Nakata, and Makoto Umeda. 2022. "Glucose Starvation-Caused Oxidative Stress Induces Inflammation and Autophagy in Human Gingival Fibroblasts" Antioxidants 11, no. 10: 1907. https://doi.org/10.3390/antiox11101907
APA StyleLi, R., Kato, H., Taguchi, Y., Deng, X., Minagawa, E., Nakata, T., & Umeda, M. (2022). Glucose Starvation-Caused Oxidative Stress Induces Inflammation and Autophagy in Human Gingival Fibroblasts. Antioxidants, 11(10), 1907. https://doi.org/10.3390/antiox11101907