




Bacillus subtilis Protects the Ducks from Oxidative Stress Induced by Escherichia coli: Efficacy and Molecular Mechanism

 ,
, 
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of E. coli O88 and B. subtilis L6
2.2. Experimental Design
2.3. Oral Challenge
2.4. Sampling
2.5. Cecal E. coli
2.6. Serum Indices
2.7. Intestinal Morphology
2.8. ROS Production and ATP Level of Jejunum
2.9. RNA-Seq Analysis of the Jejunum
2.10. RT-qPCR
2.11. Statistical Analysis
3. Results
3.1. Oral Challenge, Growth Performance and Serum Parameters
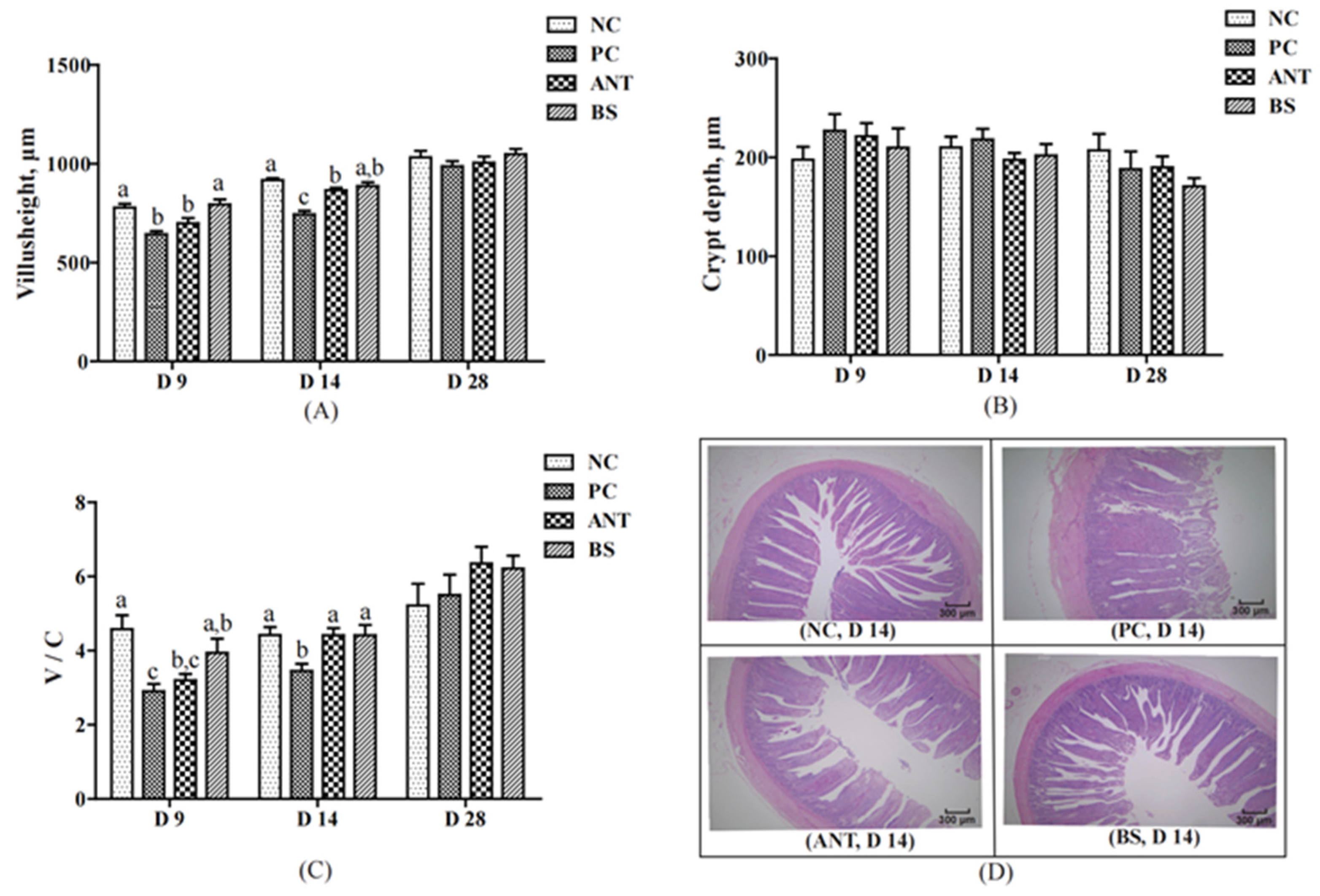
3.2. Intestinal Morphology, ROS Production and ATP Level
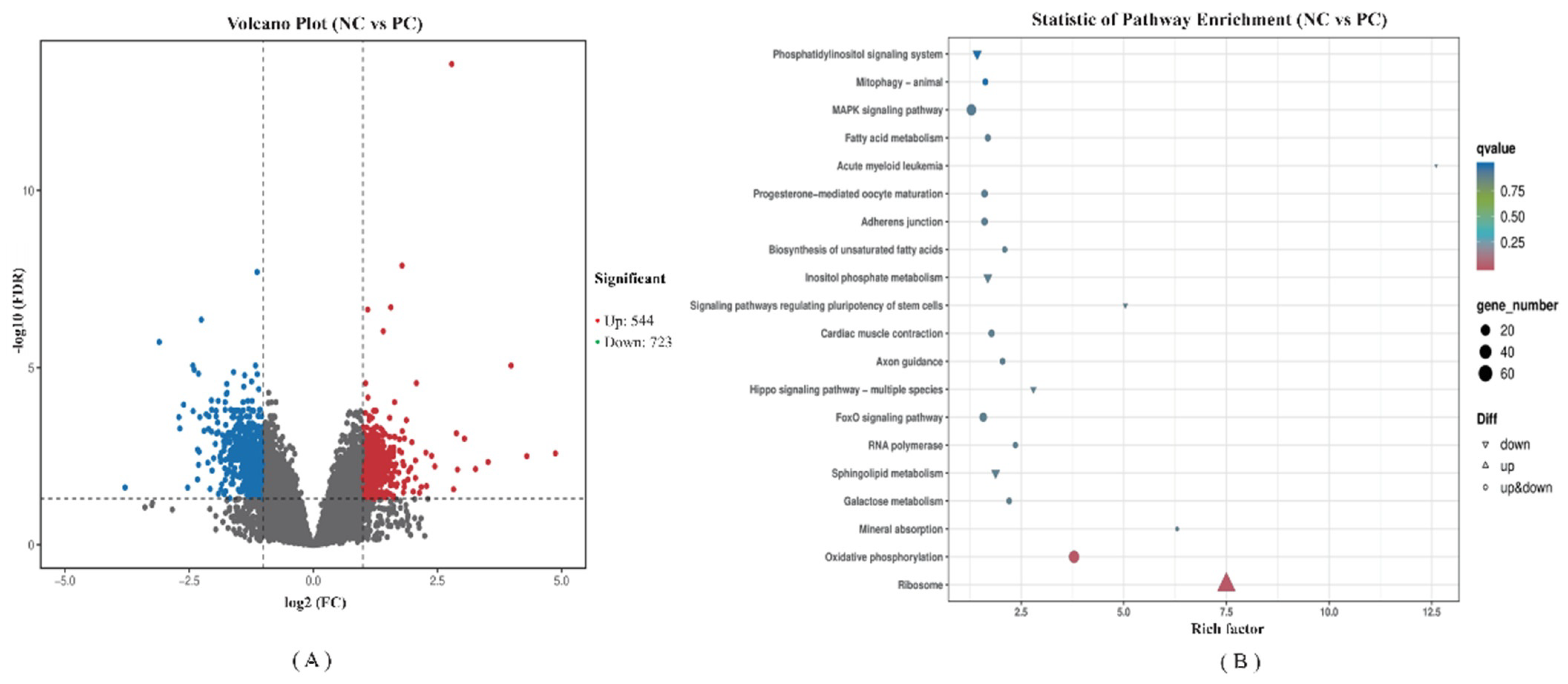
3.3. RNA-Seq Analysis of the Jejunum
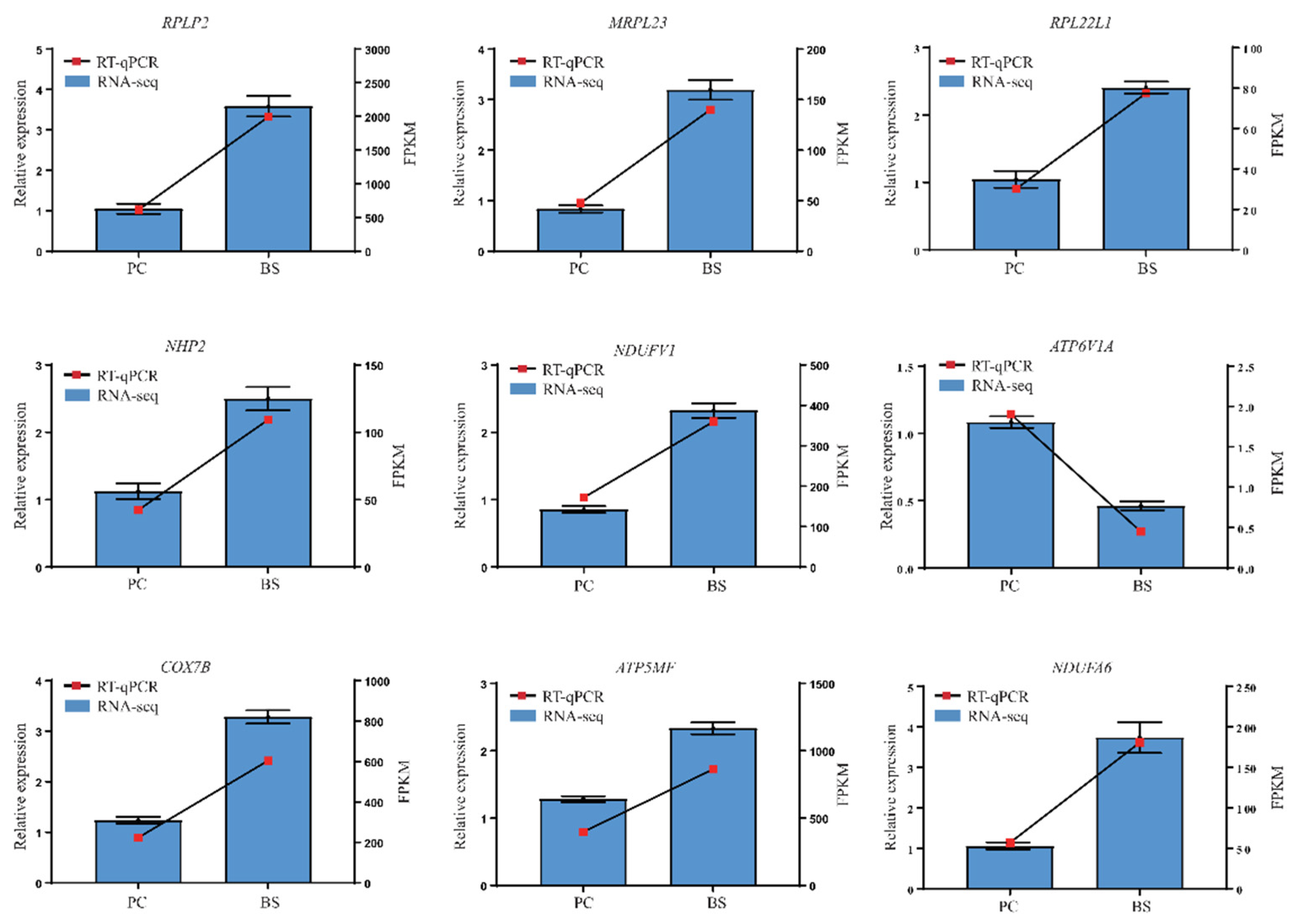
3.4. Validation of Gene Expression by Using RT-qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Da Rosa, G.; Da Silva, A.S.; Souza, C.F.; Baldissera, M.D.; Mendes, R.E.; Araujo, D.N.; Alba, D.F.; Boiago, M.M.; Stefani, L.M. Impact of Colibacillosis on Production in Laying Hens Associated with Interference of the Phosphotransfer Network and Oxidative Stress. Microb. Pathog. 2019, 130, 131–136. [Google Scholar] [CrossRef]
- Abbas, A.O.; Alaqil, A.A.; El-Beltagi, H.S.; Abd El-Atty, H.K.; Kamel, N.N. Modulating Laying Hens Productivity and Immune Performance in Response to Oxidative Stress Induced by E. Coli Challenge Using Dietary Propolis Supplementation. Antioxidants 2020, 9, 893. [Google Scholar] [CrossRef]
- Bi, S.; Qu, Y.; Shao, J.; Zhang, J.; Li, W.; Zhang, L.; Ni, J.; Cao, L. Ginsenoside Rg3 Ameliorates Stress of Broiler Chicks Induced by Escherichia Coli Lipopolysaccharide. Front. Vet. Sci. 2022, 9, 384. [Google Scholar] [CrossRef]
- Li, R.; Lin, J.; Hou, X.; Han, S.; Weng, H.; Xu, T.; Li, N.; Chai, T.; Wei, L. Characterization and Roles of Cherry Valley Duck Nlrp3 in Innate Immunity during Avian Pathogenic Escherichia Coliinfection. Front. Immunol. 2018, 9, 2300. [Google Scholar] [CrossRef]
- Jeong, J.; Lee, J.Y.; Kang, M.S.; Lee, H.J.; Kang, S.I.L.; Lee, O.M.; Kwon, Y.K.; Kim, J.H. Comparative Characteristics and Zoonotic Potential of Avian Pathogenic Escherichia Coli (APEC) Isolates from Chicken and Duck in South Korea. Microorganisms 2021, 9, 946. [Google Scholar] [CrossRef]
- Song, Y.; Yu, L.; Zhang, Y.; Dai, Y.; Wang, P.; Feng, C.; Liu, M.; Sun, S.; Xie, Z.; Wang, F. Prevalence and Characteristics of Multidrug-Resistant Mcr-1-Positive Escherichia Coli Isolates from Broiler Chickens in Tai’an, China. Poult. Sci. 2020, 99, 1117–1123. [Google Scholar] [CrossRef]
- Ma, J.; Mahfuz, S.; Wang, J.; Piao, X. Effect of Dietary Supplementation with Mixed Organic Acids on Immune Function, Antioxidative Characteristics, Digestive Enzymes Activity, and Intestinal Health in Broiler Chickens. Front. Nutr. 2021, 8, 673316. [Google Scholar] [CrossRef]
- Shini, S.; Aland, R.C.; Bryden, W.L. Avian Intestinal Ultrastructure Changes Provide Insight into the Pathogenesis of Enteric Diseases and Probiotic Mode of Action. Sci. Rep. 2021, 11, 167. [Google Scholar] [CrossRef]
- Al-Khalaifa, H.; Al-Nasser, A.; Al-Surayee, T.; Al-Kandari, S.; Al-Enzi, N.; Al-Sharrah, T.; Ragheb, G.; Al-Qalaf, S.; Mohammed, A. Effect of Dietary Probiotics and Prebiotics on the Performance of Broiler Chickens. Poult. Sci. 2019, 98, 4465–4479. [Google Scholar] [CrossRef]
- Gharib-Naseri, K.; Dorigam, J.C.P.; Doranalli, K.; Morgan, N.; Swick, R.A.; Choct, M.; Wu, S.B. Bacillus Amyloliquefaciens CECT 5940 Improves Performance and Gut Function in Broilers Fed Different Levels of Protein and/or under Necrotic Enteritis Challenge. Anim. Nutr. 2021, 7, 185–197. [Google Scholar] [CrossRef]
- Rehman, A.; Arif, M.; Sajjad, N.; Al-Ghadi, M.Q.; Alagawany, M.; Abd El-Hack, M.E.; Alhimaidi, A.R.; Elnesr, S.S.; Almutairi, B.O.; Amran, R.A.; et al. Dietary Effect of Probiotics and Prebiotics on Broiler Performance, Carcass, and Immunity. Poult. Sci. 2020, 99, 6946–6953. [Google Scholar] [CrossRef]
- Zheng, A.; Luo, J.; Meng, K.; Li, J.; Bryden, W.L.; Chang, W.; Zhang, S.; Wang, L.X.N.; Liu, G.; Yao, B. Probiotic (Enterococcus Faecium) Induced Responses of the Hepatic Proteome Improves Metabolic Efficiency of Broiler Chickens (Gallus Gallus). BMC Genom. 2016, 17, 89. [Google Scholar] [CrossRef]
- Jeong, J.S.; Kim, I.H. Effect of Bacillus Subtilis C-3102 Spores as a Probiotic Feed Supplement on Growth Performance, Noxious Gas Emission, and Intestinal Microflora in Broilers. J. Name Poult. Sci. 2014, 93, 3097–3103. [Google Scholar] [CrossRef]
- Bai, K.; Huang, Q.; Zhang, J.; He, J.; Zhang, L.; Wang, T. Supplemental Effects of Probiotic Bacillus Subtilis FmbJ on Growth Performance, Antioxidant Capacity, and Meat Quality of Broiler Chickens. Poult. Sci. 2017, 96, 74–82. [Google Scholar] [CrossRef]
- Nguyen, A.T.V.; Nguyen, D.V.; Tran, M.T.; Nguyen, L.T.; Nguyen, A.H.; Phan, T. Isolation and Characterization of Bacillus Subtilis CH16 Strain from Chicken Gastrointestinal Tracts for Use as a Feed Supplement to Promote Weight Gain in Broilers. Lett. Appl. Microbiol. 2015, 6, 580–588. [Google Scholar] [CrossRef]
- Liu, J.; Liu, G.; Chen, Z.; Zheng, A.; Cai, H.; Chang, W.; Li, C.; Chen, J.; Wu, Z. Effects of Glucose Oxidase on Growth Performance, Immune Function, and Intestinal Barrier of Ducks Infected with Escherichia Coli O88. Poult. Sci. 2020, 99, 6549–6558. [Google Scholar] [CrossRef]
- Nikoskelainen, S.; Ouwehand, A.C.; Bylund, G.; Salminen, S.; Lilius, E.M. Immune Enhancement in Rainbow Trout (Oncorhynchus Mykiss) by Potential Probiotic Bacteria (Lactobacillus Rhamnosus). Fish. Shellfish Immunol. 2003, 15, 443–452. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.J.; Cuthi, I.; Emerson, M.; Altman, D.G. Improving Bioscience Research Reporting: The ARRIVE Guidelines for Reporting Animal Research. Vet. Clin. Pathol. 2012, 41, 27–31. [Google Scholar] [CrossRef]
- Watkins, E.J.; Butler, P.J.; Kenyon, B.P. Posthatch Growth of the Digestive System in Wild and Domesticated Ducks. Br. Poult. Sci. 2004, 45, 331–341. [Google Scholar] [CrossRef]
- Manafi, M.; Khalaji, S.; Hedayati, M.; Pirany, N. Efficacy of Bacillus Subtilis and Bacitracin Methylene Disalicylate on Growth Performance, Digestibility, Blood Metabolites, Immunity, and Intestinal Microbiota after Intramuscular Inoculation with Escherichia Coli in Broilers. Poult. Sci. 2017, 96, 1174–1183. [Google Scholar] [CrossRef]
- Abo El-Maaty, H.A.; EL-Khateeb, A.Y.; Al-Khalaifah, H.; El-Sayed, E.S.A.; Hamed, S.; El-Said, E.A.; Mahrose, K.M.; Metwally, K.; Mansour, A.M. Effects of Ecofriendly Synthesized Calcium Nanoparticles with Biocompatible Sargassum Latifolium Algae Extract Supplementation on Egg Quality and Scanning Electron Microscopy Images of the Eggshell of Aged Laying Hens. Poult. Sci. 2021, 100, 675. [Google Scholar] [CrossRef]
- Lin, Q.; Zhao, J.; Xie, K.; Wang, Y.; Hu, G.; Jiang, G.; Dai, Q.; Fan, Z.; He, J.; He, X.; et al. Magnolol Additive as a Replacer of Antibiotic Enhances the Growth Performance of Linwu Ducks. Anim. Nutr. 2017, 3, 132–138. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Y.; Han, H.; Zhang, L.; Wang, T. Bisdemethoxycurcumin Protects Small Intestine from Lipopolysaccharide-Induced Mitochondrial Dysfunction via Activating Mitochondrial Antioxidant Systems and Mitochondrial Biogenesis in Broiler Chickens. Oxid. Med. Cell. Longev. 2021, 2021, 9927864. [Google Scholar] [CrossRef]
- Zhang, J.; Bai, K.W.; He, J.; Niu, Y.; Lu, Y.; Zhang, L.; Wang, T. Curcumin Attenuates Hepatic Mitochondrial Dysfunction through the Maintenance of Thiol Pool, Inhibition of MtDNA Damage, and Stimulation of the Mitochondrial Thioredoxin System in Heat-Stressed Broilers. J. Anim. Sci. 2018, 96, 867–879. [Google Scholar] [CrossRef]
- Florea, L.; Song, L.; Salzberg, S.L. Thousands of Exon Skipping Events Differentiate among Splicing Patterns in Sixteen Human Tissues. F1000Research 2013, 2, 188. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, J.Y.; Yu, Y.H. Bacillus Subtilis–Fermented Products Ameliorate the Growth Performance and Alter Cecal Microbiota Community in Broilers under Lipopolysaccharide Challenge. Poult. Sci. 2021, 100, 875–886. [Google Scholar] [CrossRef]
- Dong, Y.; Li, R.; Liu, Y.; Ma, L.; Zha, J.; Qiao, X.; Chai, T.; Wu, B. Benefit of Dietary Supplementation with Bacillus Subtilis BYS2 on Growth Performance, Immune Response, and Disease Resistance of Broilers. Probiot. Antimicrob. Proteins 2020, 12, 1385–1397. [Google Scholar] [CrossRef]
- Li, K.; Fu, W.; Bo, Y.; Zhu, Y. Effect of Albumin-Globulin Score and Albumin to Globulin Ratio on Survival in Patients with Heart Failure: A Retrospective Cohort Study in China. BMJ Open 2018, 8, e022960. [Google Scholar] [CrossRef] [Green Version]
- Lumeij, J.T. Avian Clinical Biochemistry. Clin. Biochem. Domest. Anim. 1997, 5, 857–883. [Google Scholar] [CrossRef]
- Javed, M.; Ahmad, M.I.; Usmani, N.; Ahmad, M. Multiple Biomarker Responses (Serum Biochemistry, Oxidative Stress, Genotoxicity and Histopathology) in Channa Punctatus Exposed to Heavy Metal Loaded Waste Water. Sci. Rep. 2017, 7, 1675. [Google Scholar] [CrossRef]
- Erinle, T.; MacIsaac, J.; Yang, C.; Adewole, D. Effect of Red Osier Dogwood Extract on Growth Performance, Blood Biochemical Parameters, and Gut Functionality of Broiler Chickens Challenged or Unchallenged Intraperitoneally with Salmonella Enteritidis Lipopolysaccharide. Poult. Sci. 2022, 101, 101861. [Google Scholar] [CrossRef]
- Sharma, V.; Jakhar, K.K.; Nehra, V.; Kumar, S. Biochemical Studies in Experimentally Escherichia Coli Infected Broiler Chicken Supplemented with Neem (Azadirachta Indica) Leaf Extract. Vet. World 2015, 8, 1340. [Google Scholar] [CrossRef]
- Tomlinson, J.E.; Blikslager, A.T. Interactions between Lipopolysaccharide and the Intestinal Epithelium. J. Am. Vet. Med. Assoc. 2004, 224, 1446–1452. [Google Scholar] [CrossRef]
- Qu, L.; Tan, W.; Yang, J.; Lai, L.; Liu, S.; Wu, J.; Zou, W. Combination Compositions Composed of L-Glutamine and Si-Jun-Zi-Tang Might Be a Preferable Choice for 5-Fluorouracil-Induced Intestinal Mucositis: An Exploration in a Mouse Model. Front. Pharmacol. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Leng, Y.; Yi, M.; Fan, J.; Bai, Y.; Ge, Q.; Yao, G. Effects of Acute Intra-Abdominal Hypertension on Multiple Intestinal Barrier Functions in Rats. Sci. Rep. 2016, 6, 22814. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free Radicals and Antioxidants in Normal Physiological Functions and Human Disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 Regulatory Network Provides an Interface between Redox and Intermediary Metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef]
- Giustarini, D.; Dalle-Donne, I.; Tsikas, D.; Rossi, R. Oxidative Stress and Human Diseases: Origin, Link, Measurement, Mechanisms, and Biomarkers. Crit. Rev. Clin. Lab. Sci. 2009, 46, 241–281. [Google Scholar] [CrossRef]
- Urso, M.L.; Clarkson, P.M. Oxidative Stress, Exercise, and Antioxidant Supplementation. Toxicology 2003, 189, 41–54. [Google Scholar] [CrossRef]
- Yang, T.C.; Zhang, S.W.; Sun, L.N.; Wang, H.; Ren, A.M. Magnolol Attenuates Sepsis-Induced Gastrointestinal Dysmotility in Rats by Modulating Inflammatory Mediators. World J. Gastroenterol. 2008, 14, 7353. [Google Scholar] [CrossRef]
- Lasker, S.; Rahman, M.M.; Parvez, F.; Zamila, M.; Miah, P.; Nahar, K.; Kabir, F.; Sharmin, S.B.; Subhan, N.; Ahsan, G.U.; et al. High-Fat Diet-Induced Metabolic Syndrome and Oxidative Stress in Obese Rats Are Ameliorated by Yogurt Supplementation. Sci. Rep. 2019, 9, 20026. [Google Scholar] [CrossRef]
- Tovar-Ramírez, D.; Mazurais, D.; Gatesoupe, J.F.; Quazuguel, P.; Cahu, C.L.; Zambonino-Infante, J.L. Dietary Probiotic Live Yeast Modulates Antioxidant Enzyme Activities and Gene Expression of Sea Bass (Dicentrarchus Labrax) Larvae. Aquaculture 2010, 300, 142–147. [Google Scholar] [CrossRef]
- Plessas, S.; Bosnea, L.; Alexopoulos, A.; Bezirtzoglou, E. Potential Effects of Probiotics in Cheese and Yogurt Production: A Review. Eng. Life Sci. 2012, 12, 433–440. [Google Scholar] [CrossRef]
- Vasanth, G.K.; Kiron, V.; Kulkarni, A.; Dahle, D.; Lokesh, J.; Kitani, Y. A Microbial Feed Additive Abates Intestinal Inflammation in Atlantic Salmon. Front. Immunol. 2015, 6, 409. [Google Scholar] [CrossRef]
- Daneshmand, A.; Kermanshahi, H.; Sekhavati, M.H.; Javadmanesh, A.; Ahmadian, M. Antimicrobial Peptide, CLF36, Affects Performance and Intestinal Morphology, Microflora, Junctional Proteins, and Immune Cells in Broilers Challenged with E. coli. Sci. Rep. 2019, 9, 14167. [Google Scholar] [CrossRef]
- Inatomi, T.; Otomaru, K. Effect of Dietary Probiotics on the Semen Traits and Antioxidative Activity of Male Broiler Breeders. Sci. Rep. 2018, 8, 5874. [Google Scholar] [CrossRef]
- Ekim, B.; Calik, A.; Ceylan, A.; Saçaklı, P. Effects of Paenibacillus Xylanexedens on Growth Performance, Intestinal Histomorphology, Intestinal Microflora, and Immune Response in Broiler Chickens Challenged with Escherichia Coli K88. Poult. Sci. 2020, 99, 214–223. [Google Scholar] [CrossRef]
- Liew, W.P.P.; Mohd-Redzwan, S.; Than, L.T.L. Gut Microbiota Profiling of Aflatoxin B1-Induced Rats Treated with Lactobacillus Casei Shirota. Toxins 2019, 11, 49. [Google Scholar] [CrossRef] [Green Version]
- Cimolai, M.C.; Vanasco, V.; Marchini, T.; Magnani, N.D.; Evelson, P.; Alvarez, S. α-Lipoic Acid Protects Kidney from Oxidative Stress and Mitochondrial Dysfunction Associated to Inflammatory Conditions. Food Funct. 2014, 5, 3143–3150. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Pearce, E.J. Immunometabolism Governs Dendritic Cell and Macrophage Function. J. Exp. Med. 2016, 213, 15–23. [Google Scholar] [CrossRef]
- Kingdom, U.; Kingdom, U.; London, F.; Health, N.; Foundation, S.; Kingdom, U.; Kingdom, U. Mitochondrial Dynamics Controls T Cell Fate Through Metabolic Programming. Cell 2017, 166, 63–76. [Google Scholar] [CrossRef]
- Ashton, T.M.; Gillies McKenna, W.; Kunz-Schughart, L.A.; Higgins, G.S. Oxidative Phosphorylation as an Emerging Target in Cancer Therapy. Clin. Cancer Res. 2018, 24, 2482–2490. [Google Scholar] [CrossRef]
- Fujiwara, H.; Seike, K.; Brooks, M.D.; Mathew, A.V.; Kovalenko, I.; Pal, A.; Lee, H.J.; Peltier, D.; Kim, S.; Liu, C.; et al. Mitochondrial Complex II in Intestinal Epithelial Cells Regulates T Cell-Mediated Immunopathology. Nat. Immunol. 2021, 22, 1440–1451. [Google Scholar] [CrossRef]
- Xiong, W.; Wang, R.; Mao, W.; Wu, Y.; Wang, D.; Hu, Y.; Liu, J. Icariin and Its Phosphorylated Derivatives Reduce Duck Hepatitis A Virus Serotype 1-Induced Oxidative Stress and Inflammatory Damage in Duck Embryonic Hepatocytes through Mitochondrial Regulation. Res. Vet. Sci. 2021, 139, 133–139. [Google Scholar] [CrossRef]
- Czarny, P.; Wigner, P.; Galecki, P.; Sliwinski, T. The Interplay between Inflammation, Oxidative Stress, DNA Damage, DNA Repair and Mitochondrial Dysfunction in Depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 80, 309–321. [Google Scholar] [CrossRef]
- Pi, D.; Liu, Y.; Shi, H.; Li, S.; Odle, J.; Lin, X.; Zhu, H.; Chen, F.; Hou, Y.; Leng, W. Dietary Supplementation of Aspartate Enhances Intestinal Integrity and Energy Status in Weanling Piglets after Lipopolysaccharide Challenge. J. Nutr. Biochem. 2014, 25, 456–462. [Google Scholar] [CrossRef]
- Li, C.; Li, S.; Liu, J.; Cai, H.; Liu, G.; Deng, X.; Chang, W. Escherichia Coli O88 Induces Intestinal Damage and Inflammatory Response through the Oxidative Phosphorylation and Ribosome Pathway in Pekin Ducks. Front. Cell. Infect. Microbiol. 2022, 12, 1187. [Google Scholar] [CrossRef]
- Kim, S.; Li, Q.; Dang, C.V.; Lee, L.A. Induction of Ribosomal Genes and Hepatocyte Hypertrophy by Adenovirus-Mediated Expression of c-Myc in Vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 11198–11202. [Google Scholar] [CrossRef] [Green Version]
- Mohammedsaeed, W.; Cruickshank, S.; McBain, A.J.; O’Neill, C.A. Lactobacillus Rhamnosus GG Lysate Increases Re-Epithelialization of Keratinocyte Scratch Assays by Promoting Migration. Sci. Rep. 2015, 5, 16147. [Google Scholar] [CrossRef]
- Vats, D.; Mukundan, L.; Odegaard, J.I.; Zhang, L.; Smith, K.L.; Morel, C.R.; Greaves, D.R.; Murray, P.J.; Chawla, A. Oxidative Metabolism and PGC-1β Attenuate Macrophage-Mediated Inflammation. Cell Metab. 2006, 4, 13–24. [Google Scholar] [CrossRef]
- Rodríguez-Prados, J.-C.; Través, P.G.; Cuenca, J.; Rico, D.; Aragonés, J.; Martín-Sanz, P.; Cascante, M.; Boscá, L. Substrate Fate in Activated Macrophages: A Comparison between Innate, Classic, and Alternative Activation. J. Immunol. 2010, 185, 605–614. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Days | Dietary Treatment | SEM | p-Value | |||
|---|---|---|---|---|---|---|---|
| NC | PC | ANT | BS | ||||
| BW, g | 9 | 301.7 ab | 290.1 b | 311.0 a | 312.3 a | 2.746 | 0.006 |
| 14 | 600.5 a | 557.5 b | 616.4 a | 614.8 a | 7.274 | 0.005 | |
| 28 | 1690.1 ab | 1558.1 b | 1740.2 a | 1787.9 a | 28.26 | 0.019 | |
| ADG, g/(duck-d) | 1–9 | 27.6 ab | 27.1 b | 29.1 ab | 29.2 a | 0.331 | 0.036 |
| 9–14 | 59.8 a | 52.8 b | 60.1 a | 60.2 a | 1.063 | 0.021 | |
| 14–28 | 77.5 ab | 72.4 b | 80.1 ab | 83.2 a | 1.559 | 0.080 | |
| 1–28 | 58.6 ab | 53.8 b | 60.3 a | 62.0 a | 1.018 | 0.017 | |
| ADFI, g/(duck-d) | 1–9 | 35.5 | 36.3 | 38.1 | 37.2 | 0.407 | 0.114 |
| 9–14 | 77.2 | 74.4 | 80.4 | 80.9 | 1.219 | 0.209 | |
| 14–28 | 134.7 | 137.7 | 140.6 | 143.7 | 2.185 | 0.525 | |
| 1–28 | 90.2 | 92.2 | 94.9 | 95.8 | 1.227 | 0.364 | |
| F/G ratio | 1–9 | 1.29 | 1.34 | 1.30 | 1.27 | 0.010 | 0.076 |
| 9–14 | 1.30 | 1.39 | 1.33 | 1.34 | 0.017 | 0.420 | |
| 14–28 | 1.74 b | 1.91 a | 1.76 b | 1.73 b | 0.025 | 0.019 | |
| 1–28 | 1.54 b | 1.72 a | 1.58 b | 1.55 b | 0.023 | 0.006 | |
| Parameter | Days | Dietary Treatment | SEM | p-Value | |||
|---|---|---|---|---|---|---|---|
| NC | PC | ANT | BS | ||||
| Albumin, (g/L) | 9 | 10.18 a | 7.60 b | 8.25 ab | 9.30 ab | 0.351 | 0.034 |
| 14 | 10.03 a | 7.50 b | 8.90 ab | 9.07 ab | 0.294 | 0.012 | |
| 28 | 9.02 | 8.27 | 9.37 | 8.45 | 0.426 | 0.808 | |
| Globulin, (g/L) | 9 | 11.43 b | 15.08 a | 13.35 ab | 13.27 ab | 0.466 | 0.040 |
| 14 | 10.83 | 13.06 | 11.35 | 11.00 | 0.827 | 0.164 | |
| 28 | 14.63 | 13.57 | 14.95 | 13.13 | 0.749 | 0.825 | |
| A/G ratio | 9 | 0.86 a | 0.54 b | 0.61 b | 0.71 ab | 0.011 | 0.042 |
| 14 | 0.89 a | 0.59 b | 0.77 ab | 0.81 a | 0.094 | 0.029 | |
| 28 | 0.66 | 0.60 | 0.63 | 0.65 | 0.009 | 0.073 | |
| LPS, (EU/mL) | 9 | 0.56 b | 0.63 a | 0.60 a | 0.59 ab | 0.007 | 0.002 |
| 14 | 0.58 c | 0.62 ab | 0.65 a | 0.61 bc | 0.008 | 0.002 | |
| 28 | 0.66 | 0.68 | 0.73 | 0.66 | 0.011 | 0.114 | |
| Samples ID | Clean Reads | Clean Bases | GC Content (%) | % ≥ Q30 (%) | Mapped Reads | Uniq Mapped Reads |
|---|---|---|---|---|---|---|
| PC-1 | 20,587,116 | 6,140,986,574 | 50.45% | 95.07% | 36,927,117(89.69%) | 33,262,066(80.78%) |
| PC-2 | 26,608,953 | 7,945,804,612 | 51.41% | 95.34% | 47,310,598(88.90%) | 42,009,221(78.94%) |
| PC-3 | 30,147,965 | 9,017,205,702 | 51.88% | 94.67% | 53,396,826(88.56%) | 48,909,124(81.12%) |
| PC-4 | 31,570,825 | 9,433,393,492 | 51.96% | 94.50% | 57,606,737(91.23%) | 53,044,012(84.01%) |
| PC-5 | 29,190,041 | 8,715,512,362 | 51.83% | 94.60% | 52,130,891(89.30%) | 47,333,397(81.08%) |
| PC-6 | 30,843,988 | 9,213,607,330 | 51.72% | 94.44% | 54,666,734(88.62%) | 48,497,682(78.62%) |
| BS-1 | 28,473,759 | 8,511,148,562 | 51.82% | 94.94% | 51,884,478(91.11%) | 46,221,094(81.16%) |
| BS-2 | 29,369,238 | 8,772,354,542 | 51.87% | 94.84% | 53,531,602(91.14%) | 49,849,931(84.87%) |
| BS-3 | 23,512,394 | 7,028,365,784 | 53.05% | 94.78% | 54,958,510(88.58%) | 49,777,431(80.23%) |
| BS-4 | 27,698,720 | 8,275,074,754 | 51.89% | 94.74% | 48,687,386(87.89%) | 42,982,959(77.59%) |
| BS-5 | 25,829,435 | 7,728,346,758 | 51.82% | 94.32% | 47,421,641(91.80%) | 42,797,776(82.85%) |
| BS-6 | 25,960,910 | 7,758,803,536 | 52.15% | 95.19% | 45,702,452(88.02%) | 40,362,236(77.74%) |
| Gene ID | Gene Name | p-Value | Regulated | KEGG Pathway Annotation |
|---|---|---|---|---|
| 101801662 | RPL27A | <0.001 | up | Ribosome (ko03010) |
| 101792873 | RPL32 | <0.001 | up | Ribosome (ko03010) |
| 101803698 | RPL22L1 | <0.001 | up | Ribosome (ko03010) |
| 101794125 | RPLP2 | <0.001 | up | Ribosome (ko03010) |
| 101798421 | RPL35A | <0.001 | up | Ribosome (ko03010) |
| 101793195 | RPL8 | <0.001 | up | Ribosome (ko03010) |
| 101793091 | MRPS11 | <0.001 | up | Ribosome (ko03010) |
| 101794248 | MRPL23 | <0.001 | up | Ribosome (ko03010) |
| 101798947 | NHP2 | <0.001 | up | Ribosome biogenesis in eukaryotes (ko03008) |
| 101804664 | RPL30 | <0.001 | up | Ribosome (ko03010) |
| Gene ID | Gene Symbol | p-Value | Regulated | KEGG Pathway Annotation |
|---|---|---|---|---|
| 101802630 | NDUFV1 | <0.001 | up | Oxidative phosphorylation (ko00190) |
| 101794884 | NDUFB5 | <0.001 | up | Oxidative phosphorylation (ko00190) |
| 101798419 | NDUFA2 | <0.001 | up | Oxidative phosphorylation (ko00190) |
| 101791901 | NDUFA6 | <0.001 | up | Oxidative phosphorylation (ko00190) |
| 101800107 | NDUFB3 | <0.001 | up | Oxidative phosphorylation (ko00190) |
| 101795772 | COX5A | <0.001 | up | Oxidative phosphorylation (ko00190) |
| 119713461 | COX5B | <0.001 | up | Oxidative phosphorylation (ko00190) |
| 101796089 | COX7A | <0.001 | up | Oxidative phosphorylation (ko00190) |
| 101800937 | COX7B | <0.001 | up | Oxidative phosphorylation (ko00190) |
| 101802805 | ATP5MF | <0.001 | up | Oxidative phosphorylation (ko00190) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Li, Y.; Li, S.; Chen, S.; Liu, G.; Deng, X.; Chang, W.; Cai, H. Bacillus subtilis Protects the Ducks from Oxidative Stress Induced by Escherichia coli: Efficacy and Molecular Mechanism. Antioxidants 2022, 11, 1951. https://doi.org/10.3390/antiox11101951
Li C, Li Y, Li S, Chen S, Liu G, Deng X, Chang W, Cai H. Bacillus subtilis Protects the Ducks from Oxidative Stress Induced by Escherichia coli: Efficacy and Molecular Mechanism. Antioxidants. 2022; 11(10):1951. https://doi.org/10.3390/antiox11101951
Chicago/Turabian StyleLi, Chong, Yang Li, Shuzhen Li, Si Chen, Guohua Liu, Xuejuan Deng, Wenhuan Chang, and Huiyi Cai. 2022. "Bacillus subtilis Protects the Ducks from Oxidative Stress Induced by Escherichia coli: Efficacy and Molecular Mechanism" Antioxidants 11, no. 10: 1951. https://doi.org/10.3390/antiox11101951
APA StyleLi, C., Li, Y., Li, S., Chen, S., Liu, G., Deng, X., Chang, W., & Cai, H. (2022). Bacillus subtilis Protects the Ducks from Oxidative Stress Induced by Escherichia coli: Efficacy and Molecular Mechanism. Antioxidants, 11(10), 1951. https://doi.org/10.3390/antiox11101951