
Antioxidant and Anti-Inflammatory Activities of Sargassum macrocarpum Extracts

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Preparation of Ethanol and Water Extracts of S. macrocarpum
2.3. Assessment of Antioxidant Activities Using ABTS, DPPH, and FRAP
2.4. Assessment of Total Polyphenol and Flavonoid Content
2.5. Analysis of General Composition and Sugars
2.6. Cytotoxicity and Anti-Inflammatory Activity in RAW 264.7 Cells
2.7. Cytotoxicity and Anti-Inflammatory Activity Assessment in HaCaT Cells
2.8. Monosaccharide Composition Analysis
2.9. Statistical Analysis
3. Results and Discussion
3.1. Comparison of Antioxidant Activity
3.2. General Composition of S. macrocarpum Extracts
3.3. Anti-Inflammatory Effects of S. macrocarpum Extracts in LPS-Induced RAW 264.7 Cells
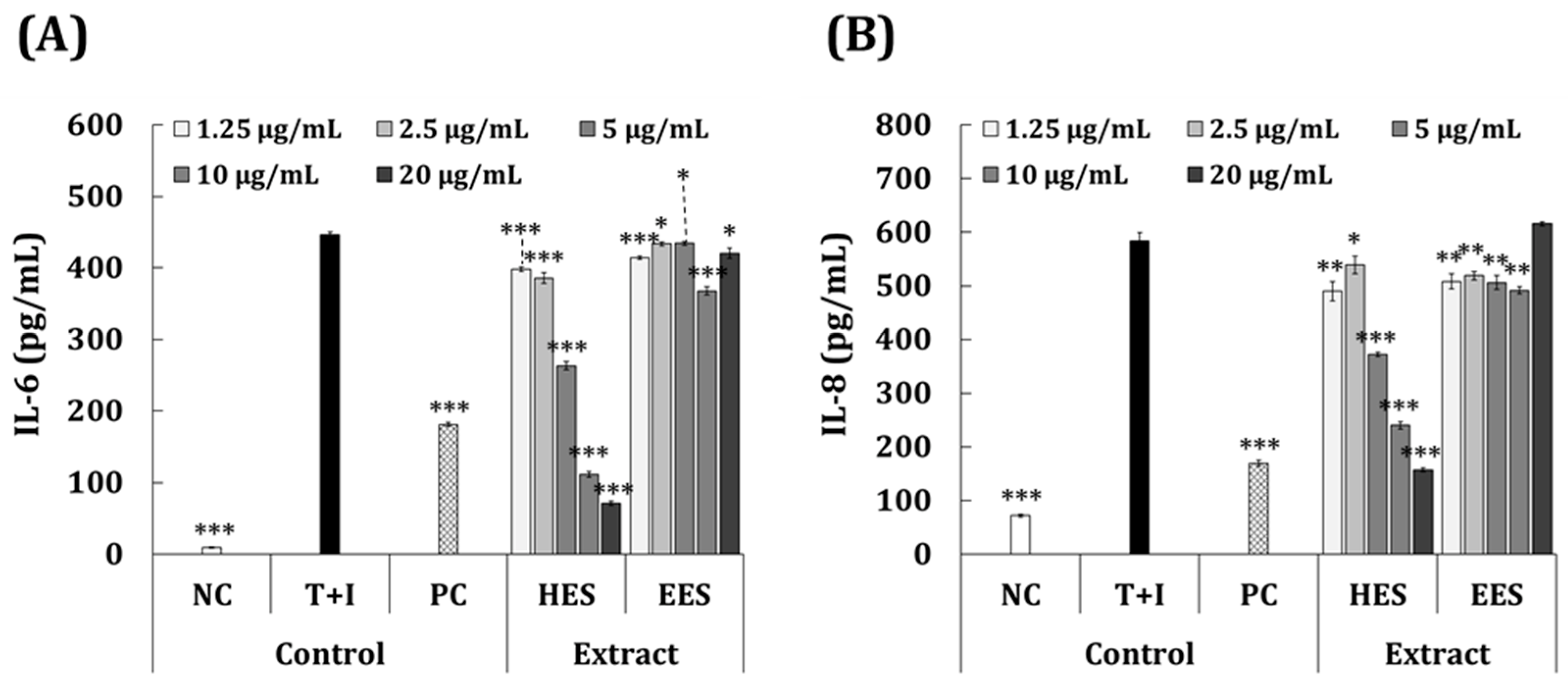
3.4. Anti-Inflammatory Effects of S. macrocarpum Extracts in T + I-Induced HaCaT Cells
3.5. Monosaccharide Composition of HES
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salmon, J.K.; Armstrong, C.A.; Ansel, J.C. The skin as an immune organ. West. J. Med. 1994, 160, 146. [Google Scholar] [PubMed]
- Bos, J.D. The skin as an organ of immunity. Clin. Exp. Immunol. 1997, 107 (Suppl. 1), 3–5. [Google Scholar] [PubMed]
- Di Meglio, P.; Perera, G.K.; Nestle, F.O. The multitasking organ: Recent insights into skin immune function. Immunity 2011, 35, 857–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadenas, E.; Davies, K.J.A. Mitochondrial free radical generation, oxidative stress, and aging. Free Radic. Biol. Med. 2000, 29, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Barja, G. The mitochondrial free radical theory of aging. Prog. Mol. Biol. Transl. Sci. 2014, 127, 1–27. [Google Scholar]
- Ratz-Lyko, A.; Arct, J.; Pytkowska, K. Methods for evaluation of cosmetic antioxidant capacity. Ski. Res. Technol. 2012, 18, 421–430. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, P.K.; Maity, N.; Nema, N.K.; Sarkar, B.K. Bioactive compounds from natural resources against skin aging. Phytomedicine 2011, 19, 64–73. [Google Scholar] [CrossRef]
- Chavoshnejad, P.; Foroughi, A.H.; Dhandapani, N.; German, G.K.; Razavi, M.J. Effect of collagen degradation on the mechanical behavior and wrinkling of skin. Phys. Rev. E 2021, 104, 034406. [Google Scholar] [CrossRef]
- Leirós, G.J.; Kusinsky, A.G.; Balañá, M.E.; Hagelin, K. Triolein reduces mmp-1 upregulation in dermal fibroblasts generated by ros production in uvb-Irradiated keratinocytes. J. Dermatol. Sci. 2017, 85, 124–130. [Google Scholar] [CrossRef]
- Oak, J.-H.; Lee, I.-K. Taxonomy of the genus Sargassum (Fucales, Phaeophyceae) from Korea i. Subgenus Bactrophycus Section Teretia. Algae 2005, 20, 77–90. [Google Scholar] [CrossRef]
- Yeon, J.; Seo, H.; Oh, S.; Choi, W. Bioethanol production from hydrolysate of seaweed Sargassum sagamianum. KSBB J. 2010, 25, 283–288. [Google Scholar]
- Gamal-Eldeen, A.M.; Ahmed, E.F.; Abo-Zeid, M.A. In vitro cancer chemopreventive properties of polysaccharide extract from the brown alga, Sargassum Latifolium. Food Chem. Toxicol. 2009, 47, 1378–1384. [Google Scholar] [CrossRef]
- Ananthi, S.; Raghavendran, H.R.B.; Sunil, A.G.; Gayathri, V.; Ramakrishnan, G.; Vasanthi, H.R. In vitro antioxidant and in vivo anti-inflammatory potential of crude polysaccharide from Turbinaria ornata (marine brown alga). Food Chem. Toxicol. 2010, 48, 187–192. [Google Scholar] [CrossRef]
- You, S.H.; Jang, M.; Kim, G.H. Inhibition of nitric oxide and lipid accumulation by Sargassum sp. seaweeds and their antioxidant properties. Korean J. Food Preserv. 2021, 28, 288–296. [Google Scholar] [CrossRef]
- Choi, M.-W.; Kim, H.-R.; Lee, H.-G.; Kim, J.-I. Effect of a Sargassum serratifolium extract on neuroinflammation induced by lipopolysaccharides in mice. Korean J. Fish. Aquat. Sci. 2019, 52, 81–86. [Google Scholar]
- Kang, J.Y.; Khan, M.N.A.; Park, N.H.; Cho, J.Y.; Lee, M.C.; Fujii, H.; Hong, Y.K. Antipyretic, analgesic, and anti-inflammatory activities of the seaweed Sargassum fulvellum and Sargassum thunbergii in mice. J. Ethnopharmacol. 2008, 116, 187–190. [Google Scholar] [CrossRef]
- Yoon, W.J.; Heo, S.J.; Han, S.C.; Lee, H.J.; Kang, G.J.; Kang, H.K.; Hyun, J.W.; Koh, Y.S.; Yoo, E.S. Anti-inflammatory effect of sargachromanol G isolated from Sargassum siliquastrum in raw 264.7 cells. Arch. Pharm. Res. 2012 358 2012, 35, 1421–1430. [Google Scholar] [CrossRef]
- Stefanucci, A.; Scioli, G.; Marinaccio, L.; Zengin, G.; Locatelli, M.; Tartaglia, A.; Valle, A.D.; Cichelli, A.; Novellino, E.; Pieretti, S.; et al. A comparative study on phytochemical fingerprint of two diverse Phaseolus vulgaris var. Tondino del Tavo and Cannellino bio extracts. Antioxidants 2022, 11, 2420. [Google Scholar] [CrossRef]
- GokhanZengin, G.; Aumeeruddy-Elalfi, Z.; Mollica, A.; Yilmaz, M.Y.; Mahomoodally, M.F. In vitro and in silico perspectives on biological and phytochemical profile of three halophyte species—A source of innovative phytopharmaceuticals from nature. Phytomedicine 2018, 38, 35–44. [Google Scholar]
- Kim, T.H.; Kim, W.J.; Park, S.Y.; Kim, H.; Chung, D.K. In vitro anti-wrinkle and skin-moisturizing effects of evening primrose (Oenothera biennis) sprout and identification of its active components. Processes 2021, 9, 145. [Google Scholar] [CrossRef]
- Stanković, M.S. Total phenolic content, flavonoid concentration and antioxidant activity of Marrubium peregrinum L. extracts. Kragujev. J. Sci. 2011, 33, 63–72. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Singleton, V.L., Orthofer, R., Lamuela-Raventós, R.M., Eds.; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Dai, J.; Wu, Y.; Chen, S.-W.; Zhu, S.; Yin, H.-P.; Wang, M.; Tang, J. Sugar compositional determination of polysaccharides from Dunaliella salina by modified RP-HPLC method of precolumn derivatization with 1-phenyl-3-methyl-5-pyrazolone. Carbohydr. Polym. 2010, 82, 629–635. [Google Scholar] [CrossRef]
- Hwang, K.C.; Shin, H.Y.; Kim, W.J.; Seo, M.S.; Kim, H. Effects of a high-molecular-weight polysaccharides isolated from Korean persimmon on the antioxidant, anti-Inflammatory, and antiwrinkle activity. Molecules 2021, 26, 1600. [Google Scholar] [CrossRef]
- Gharras, H.E. Polyphenols: Food sources, properties and applications—A review. Int. J. Food Sci. Technol. 2009, 44, 2512–2518. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Ghasemzadeh, N. Flavonoids and phenolic acids: Role and biochemical activity in plants and human. J. Med. Plants Res. 2011, 5, 6697–6703. [Google Scholar] [CrossRef]
- Laura, A.; Yahia, E.M.; Carrillo-Lopez, A. Phenolic compounds. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Yahia, E.M., Carrillo-Lopez, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 253–271. [Google Scholar]
- Ren, Y.; Wang, F.Y.; Chen, Z.J.; Lan, R.T.; Huang, R.H.; Fu, W.Q.; Gul, R.M.; Wang, J.; Xu, J.Z.; Li, Z.M. Antibacterial and anti-Inflammatory ultrahigh molecular weight polyethylene/tea polyphenol blends for artificial joint applications. J. Mater. Chem. B 2020, 8, 10428–10438. [Google Scholar] [CrossRef]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant activity of plant extracts containing phenolic compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef]
- Lee, S.-S.; Kim, H.-G.; Park, E.-H.; Kim, K.J.; Bang, M.-H.; Kim, G.; Jeon, H.-J.; Lee, C.-G.; Shin, M.C.; Kim, D.-O.; et al. Antioxidant and anti-inflammatory effects in lipopolysaccharide-induced THP-1 cells of coumarins from the bark of Hesperethusa crenulata R. Appl. Biol. Chem. 2021, 64, 90. [Google Scholar] [CrossRef]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef]
- Oishi, Y.; Manabe, I. Macrophages in inflammation, repair and regeneration. Int. Immunol. 2018, 30, 511–528. [Google Scholar] [CrossRef] [Green Version]
- Moro, C.; Palacios, I.; Lozano, M.; D’Arrigo, M.; Guillamón, E.; Villares, A.; Martínez, J.A.; García-Lafuente, A. Anti-inflammatory activity of methanolic extracts from edible mushrooms in LPS activated raw 264.7 macrophages. Food Chem. 2012, 130, 350–355. [Google Scholar] [CrossRef]
- Saraswati; Giriwono, P.E.; Iskandriati, D.; Tan, C.P.; Andarwulan, N. Sargassum seaweed as a source of anti-inflammatory substances and the potential insight of the tropical species: A review. Mar. Drugs 2019, 17, 590. [Google Scholar] [CrossRef]
- Deniaud-Bouët, E.; Hardouin, K.; Potin, P.; Kloareg, B.; Hervé, C. A review about brown algal cell walls and fucose-containing sulfated polysaccharides: Cell wall context, biomedical properties and key research challenges. Carbohydr. Polym. 2017, 175, 395–408. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | ABTS | DPPH | FRAP |
|---|---|---|---|
| (IC50, mg/mL) | (IC50, mg/mL) | (mg AA/g) | |
| HES | 1.0 ± 0.0 | 6.50 ± 0.3 | 18.8 ± 0.4 |
| EES | 16.9 ± 0.7 | 35.3 ± 3.1 | n.d. |
| Sample | Total Sugar | Uronic Acid | Protein | Total Polyphenol |
|---|---|---|---|---|
| (mg Gal/g) | (mg GalA/g) | (mg BSA/g) | (mg GA/g) | |
| HES | 127.6 ± 6.5 | 29.5 ± 0.3 | 41.8 ± 2.0 | 115.9 ± 15.3 |
| EES | 6.5 ± 3.1 | 2.5 ± 0.1 | 10.3 ± 0.4 | 3.9 ± 0.5 |
| Sugar Unit | HES (mole%) |
|---|---|
| Mannose | 5.0 ± 0.2 |
| Rhamnose | – |
| Glucuronic acid | 4.9 ± 0.3 |
| Galacturonic acid | – |
| Glucose | 35.7 ± 0.3 |
| Galactose | 5.7 ± 0.1 |
| Xylose | 13.7 ± 0.1 |
| Arabinose | 0.8 ± 0.0 |
| Fucose | 34.3 ± 0.2 |
| Sum | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Shin, H.Y.; Jeong, E.-J.; Lee, H.-D.; Hwang, K.C.; Yu, K.-W.; Lee, S.; Lee, S. Antioxidant and Anti-Inflammatory Activities of Sargassum macrocarpum Extracts. Antioxidants 2022, 11, 2483. https://doi.org/10.3390/antiox11122483
Kim H, Shin HY, Jeong E-J, Lee H-D, Hwang KC, Yu K-W, Lee S, Lee S. Antioxidant and Anti-Inflammatory Activities of Sargassum macrocarpum Extracts. Antioxidants. 2022; 11(12):2483. https://doi.org/10.3390/antiox11122483
Chicago/Turabian StyleKim, Hoon, Hyun Young Shin, Eun-Jin Jeong, Hak-Dong Lee, Ki Cheol Hwang, Kwang-Won Yu, Sullim Lee, and Sanghyun Lee. 2022. "Antioxidant and Anti-Inflammatory Activities of Sargassum macrocarpum Extracts" Antioxidants 11, no. 12: 2483. https://doi.org/10.3390/antiox11122483
APA StyleKim, H., Shin, H. Y., Jeong, E. -J., Lee, H. -D., Hwang, K. C., Yu, K. -W., Lee, S., & Lee, S. (2022). Antioxidant and Anti-Inflammatory Activities of Sargassum macrocarpum Extracts. Antioxidants, 11(12), 2483. https://doi.org/10.3390/antiox11122483