
Dietary Supplementation with Cysteine during Pregnancy Rescues Maternal Chronic Kidney Disease-Induced Hypertension in Male Rat Offspring: The Impact of Hydrogen Sulfide and Microbiota-Derived Tryptophan Metabolites

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care and Experimental Design
2.2. High Performance Liquid Chromatography–Mass Spectrometry (HPLC-MS/MS)
2.3. Liquid Chromatograph Tandem–Mass Spectrometry (LC-MS/MS)
2.4. Quantitative RT-PCR
2.5. Analysis of Gut-Microbiota Composition
2.6. Analysis of Nitric Oxide Parameters
2.7. Renal H2S-Releasing Activity
2.8. Immunohistochemistry Staining for 8-OHdG
2.9. Statistical Analysis
3. Results
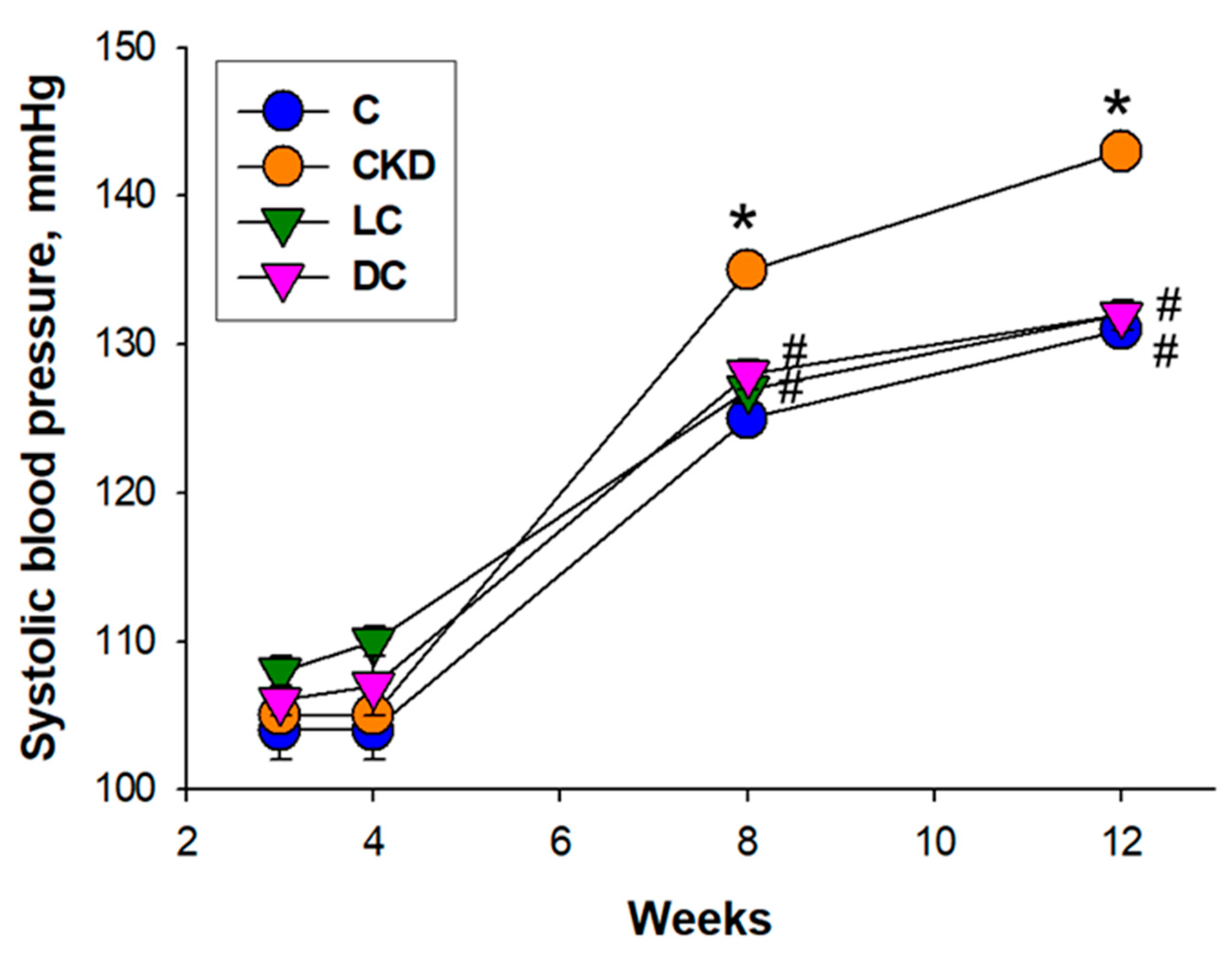
3.1. Body Weight and Blood Pressure of Male Offspring
3.2. H2S Signaling Pathway
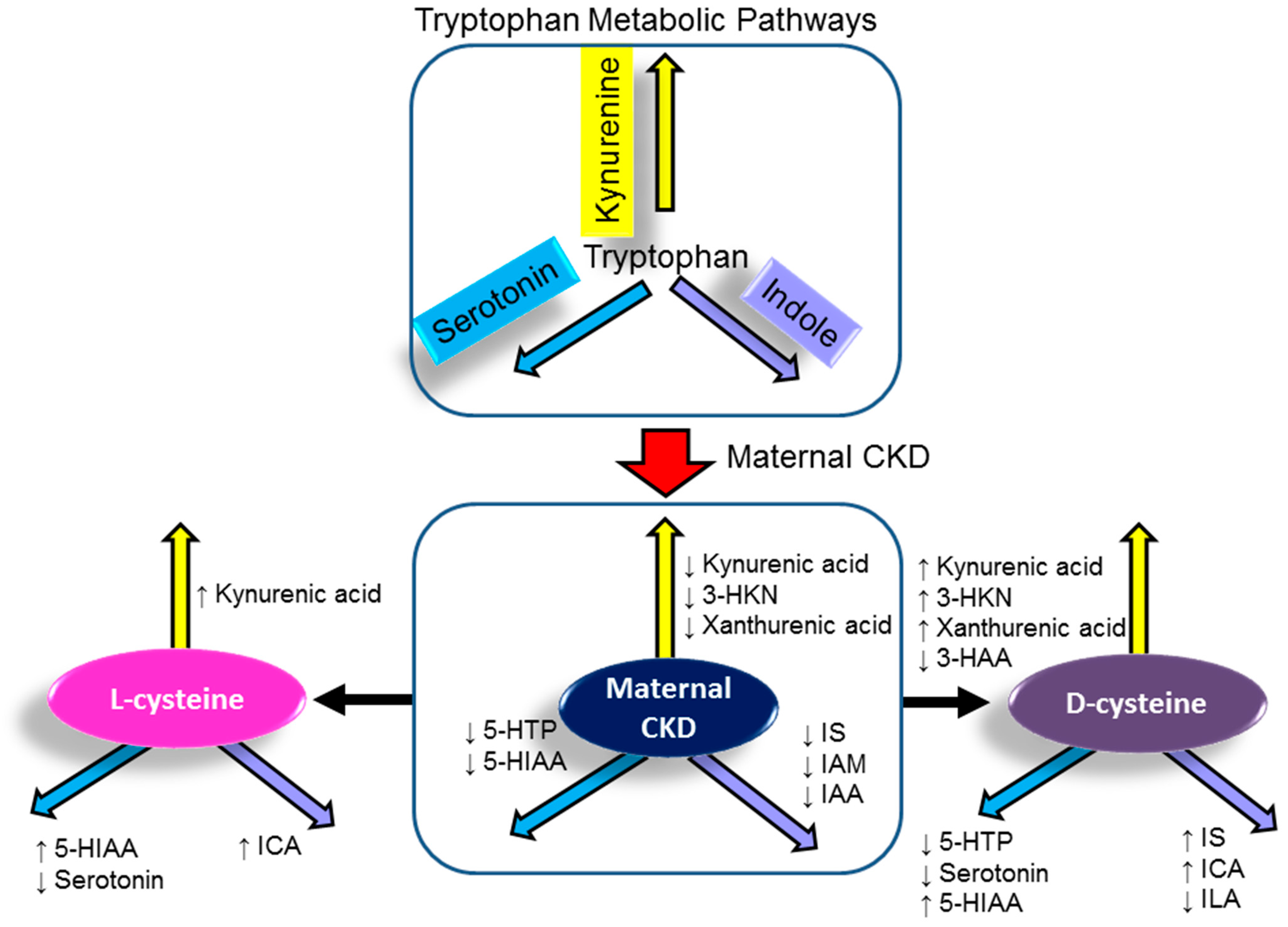
3.3. Tryptophan Metabolites
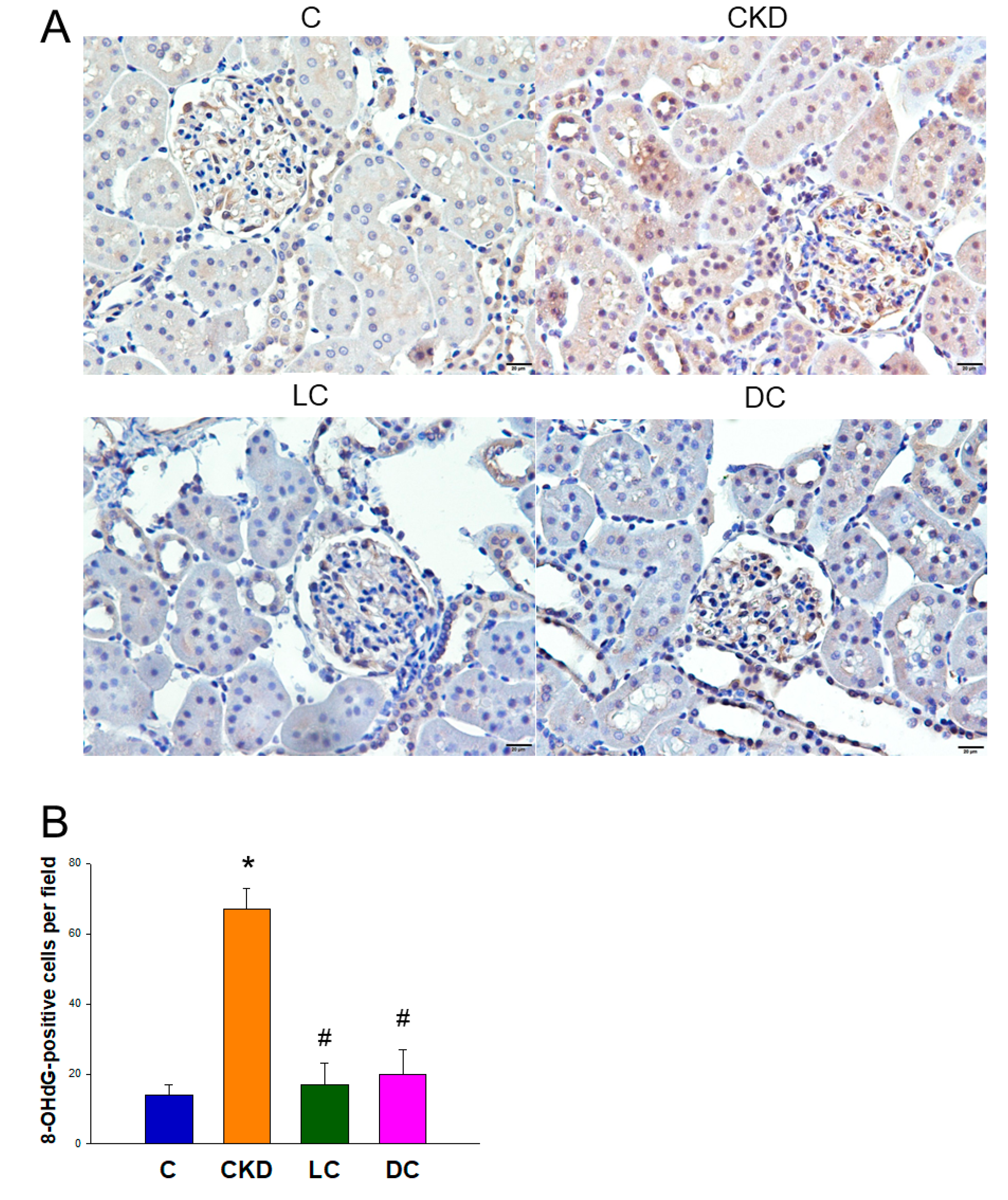
3.4. Oxidative Stress
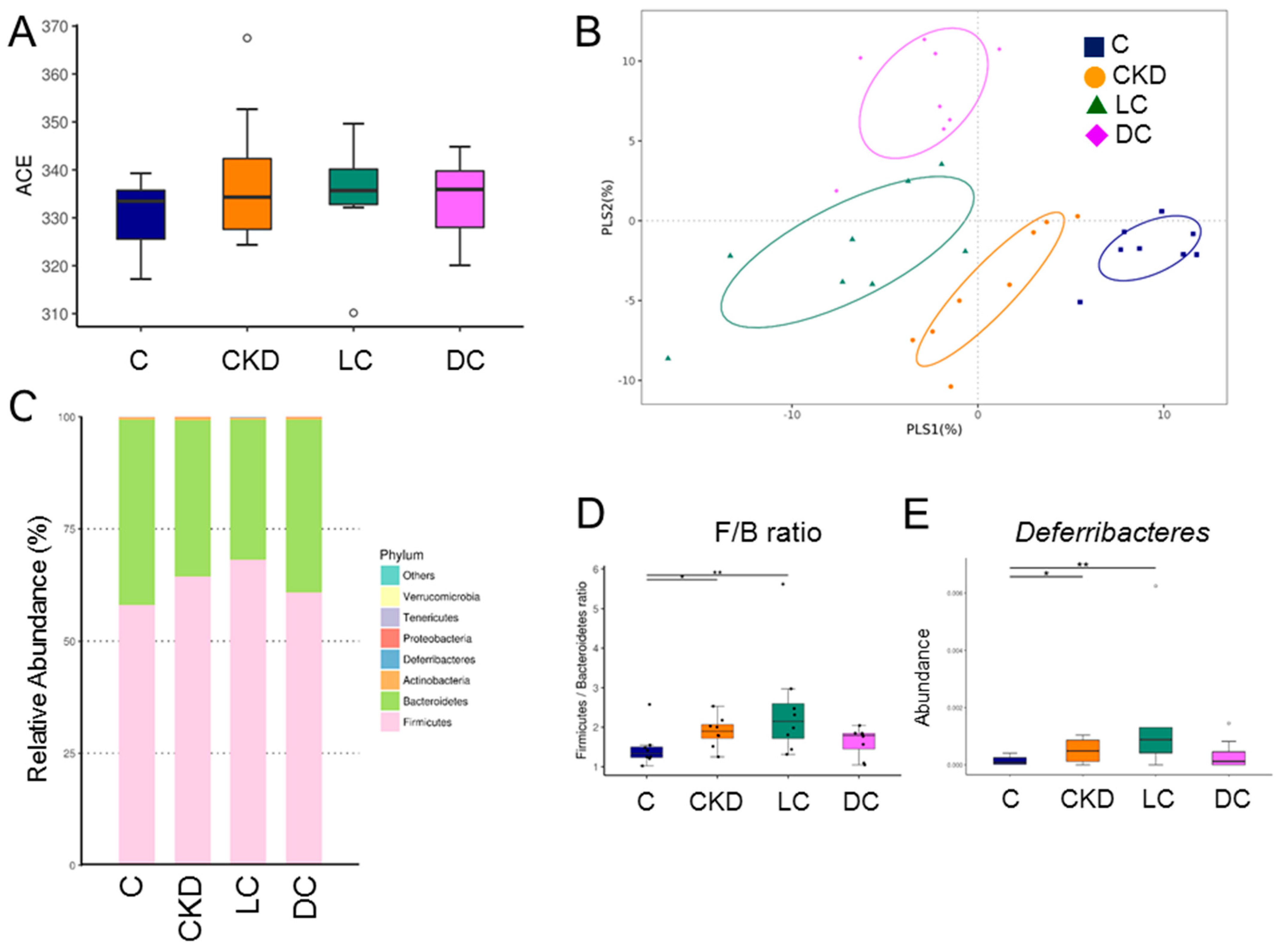
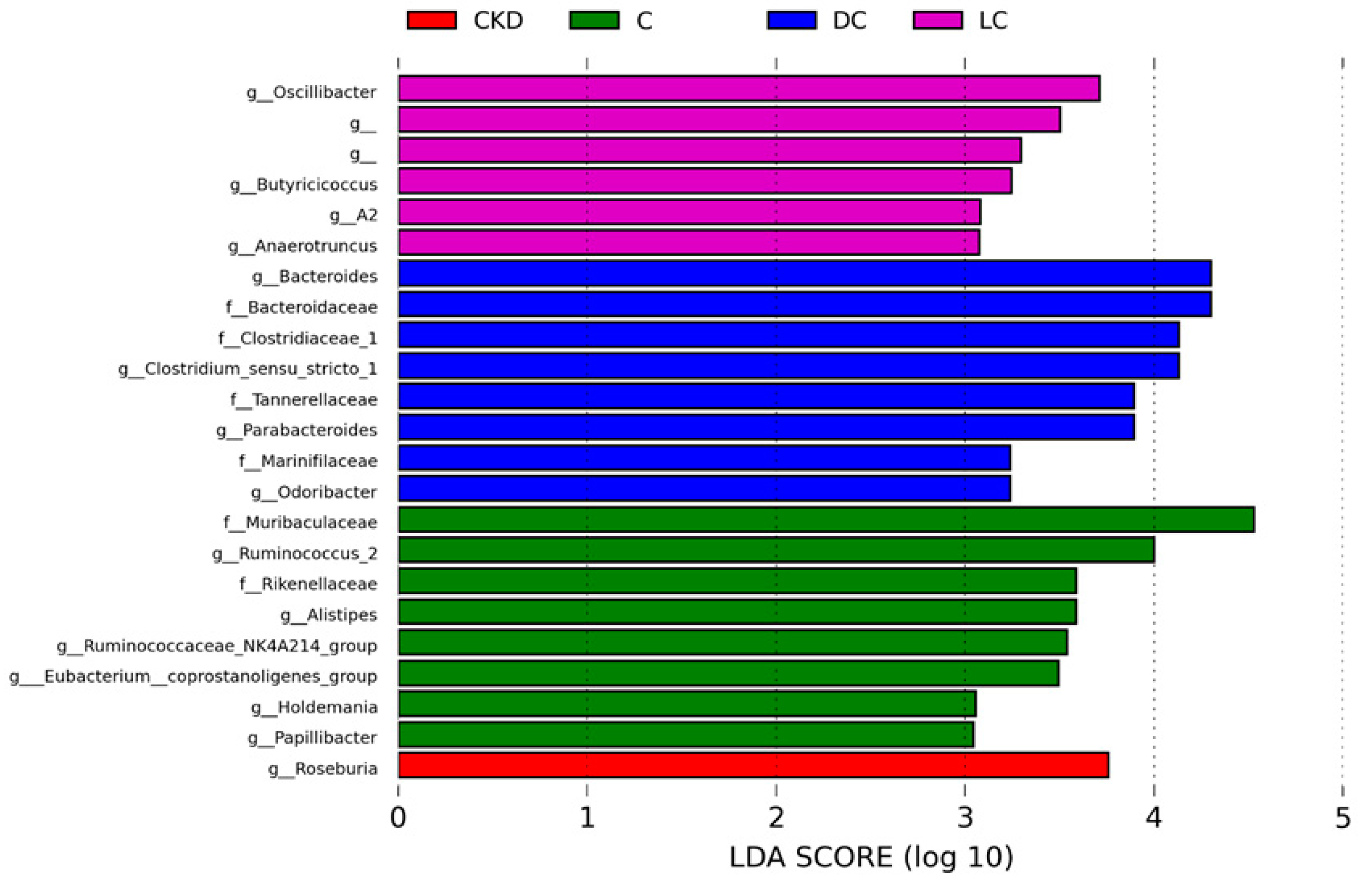
3.5. Alterations in Microbiome
3.6. NO Pathway
3.7. Renin–Angiotensin System
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanson, M.; Gluckman, P. Developmental origins of noncommunicable disease: Population and public health implications. Am. J. Clin. Nutr. 2011, 94, 1754S–1758S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccoli, G.B.; Alrukhaimi, M.; Liu, Z.H.; Zakharova, E.; Levin, A.; World Kidney Day Steering Committee. What we do and do not know about women and kidney diseases; Questions unanswered and answers unquestioned: Reflection on World Kidney Day and International Woman’s Day. Physiol. Int. 2018, 105, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Yang, H.W.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Tain, Y.L. Maternal adenine-induced chronic kidney disease programs hypertension in adult male rat offspring: Implications of nitric oxide and gut microbiome derived metabolites. Int. J. Mol. Sci. 2020, 21, 7237. [Google Scholar] [CrossRef]
- Badawy, A. Tryptophan metabolism, disposition and utilization in pregnancy. Biosci. Rep. 2015, 35, e00261. [Google Scholar] [CrossRef] [PubMed]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Sallée, M.; Dou, L.; Cerini, C.; Poitevin, S.; Brunet, P.; Burtey, S. The aryl hydrocarbon receptor-activating effect of uremic toxins from tryptophan metabolism: A new concept to understand cardiovascular complications of chronic kidney disease. Toxins 2014, 6, 934–949. [Google Scholar] [CrossRef]
- Brito, J.S.; Borges, N.A.; Esgalhado, M.; Magliano, D.C.; Soulage, C.O.; Mafra, D. Aryl hydrocarbon receptor activation in chronic kidney disease: Role of uremic toxins. Nephron 2017, 137, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Tain, Y.L. Developmental Origins of Kidney Disease: Why Oxidative Stress Matters? Antioxidants 2021, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Tain, Y.L. Developmental programming and reprogramming of hypertension and kidney disease: Impact of tryptophan metabolism. Int. J. Mol. Sci. 2020, 21, 8705. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. The physiological role of hydrogen sulfide and beyond. Nitric Oxide 2014, 41, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Tain, Y.L. Preventing developmental origins of cardiovascular disease: Hydrogen sulfide as a potential target? Antioxidants 2021, 10, 247. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, C.S. Oxidative stress and nitric oxide deficiency in the kidney: A critical link to hypertension? Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R913–R935. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Tain, Y.L. Gasotransmitters for the therapeutic prevention of hypertension and kidney disease. Int. J. Mol. Sci. 2021, 22, 7808. [Google Scholar] [CrossRef]
- Hsu, C.N.; Lin, Y.J.; Lu, P.C.; Tain, Y.L. Early supplementation of d-cysteine or l-cysteine prevents hypertension and kidney damage in spontaneously hypertensive rats exposed to high-salt intake. Mol. Nutr. Food Res. 2018, 62, 2. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N-Acetylcysteine as an antioxidant and disulphide breaking agent: The reasons why. Free Radic Res. 2018, 52, 751–762. [Google Scholar] [CrossRef]
- Lobel, L.; Cao, Y.G.; Fenn, K.; Glickman, J.N.; Garrett, W.S. Diet posttranslationally modifies the mouse gut microbial proteome to modulate renal function. Science 2020, 369, 1518–1524. [Google Scholar] [CrossRef] [PubMed]
- Reckelhoff, J.F. Gender differences in the regulation of blood pressure. Hypertension 2001, 37, 1199–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.N.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Tain, Y.L. Maternal Garlic Oil Supplementation Prevents High-Fat Diet-Induced Hypertension in Adult Rat Offspring: Implications of H2S-Generating Pathway in the Gut and Kidneys. Mol. Nutr. Food Res. 2021, 65, e2001116. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.R.; Deleon, E.R.; Gao, Y.; Hurley, K.; Sadauskas, V.; Batz, C.; Stoy, G.F. Thiosulfate: A readily accessible source of hydrogen sulfide in oxygen sensing. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R592–R603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut dysbiosis is linked to hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Tain, Y.L. Amino acids and developmental origins of hypertension. Nutrients 2020, 12, 1763. [Google Scholar] [CrossRef]
- Wen, Y.D.; Wang, H.; Zhu, Y.Z. The Drug developments of hydrogen sulfide on cardiovascular disease. Oxid. Med. Cell. Longev. 2018, 2018, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibuya, N.; Kimura, H. Production of hydrogen sulfide from d-cysteine and its therapeutic potential. Front. Endocrinol. 2013, 4, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boesmans, L.; Valles-Colomer, M.; Wang, J.; Eeckhaut, V.; Falony, G.; Ducatelle, R.; Van Immerseel, F.; Raes, J.; Verbeke, K. Butyrate Producers as Potential Next-Generation Probiotics: Safety Assessment of the Administration of Butyricicoccus pullicaecorum to Healthy Volunteers. mSystems 2018, 3, e00094-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafar, H.; Saier, M.H., Jr. Gut Bacteroides species in health and disease. Gut Microbes 2021, 13, 1848158. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Arango, L.F.; Barrett, H.L.; McIntyre, H.D.; Callaway, L.K.; Morrison, M.; Dekker Nitert, M.; SPRING Trial Group. Increased Systolic and Diastolic Blood Pressure Is Associated with Altered Gut Microbiota Composition and Butyrate Production in Early Pregnancy. Hypertension 2016, 68, 974–981. [Google Scholar] [CrossRef]
- Liu, J.R.; Miao, H.; Deng, D.Q.; Vaziri, N.D.; Li, P.; Zhao, Y.Y. Gut microbiota-derived tryptophan metabolism mediates renal fibrosis by aryl hydrocarbon receptor signaling activation. Cell. Mol. Life Sci. 2021, 78, 909–922. [Google Scholar] [CrossRef]
- Zhang, N. The role of endogenous aryl hydrocarbon receptor signaling in cardiovascular physiology. J. Cardiovasc. Dis. Res. 2011, 2, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Dalton, T.P.; Puga, A.; Shertzer, H.G. Induction of cellular oxidative stress by aryl hydrocarbon receptor activation. Chem. Biol. Interact. 2002, 141, 77–95. [Google Scholar] [CrossRef]
- Wilck, N.; Matus, M.G.; Kearney, S.M.; Olesen, S.W.; Forslund, K.; Bartolomaeus, H.; Haase, S.; Mähler, A.; Balogh, A.; Markó, L.; et al. Salt-responsive gut commensal modulates TH17 axis and disease. Nature 2017, 551, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.C.M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria with Emerging Implications to Inflammation, Cancer, and Mental Health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef] [PubMed]
- Yacoub, R.; Wyatt, C.M. Manipulating the gut microbiome to decrease uremic toxins. Kidney Int. 2017, 91, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Dai, Z.; Kou, J.; Sun, K.; Chen, J.; Yang, Y.; Wu, G.; Wu, Z. Dietary l-Tryptophan supplementation enhances the intestinal mucosal barrier function in weaned piglets: Implication of Tryptophan-metabolizing microbiota. Int. J. Mol. Sci. 2019, 20, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraf, M.K.; Piccolo, B.D.; Bowlin, A.K.; Mercer, K.E.; LeRoith, T.; Chintapalli, S.V.; Shankar, K.; Badger, T.M.; Yeruva, L. Formula diet driven microbiota shifts tryptophan metabolism from serotonin to tryptamine in neonatal porcine colon. Microbiome 2017, 5, 77. [Google Scholar] [CrossRef] [PubMed]
- Forrester, S.J.; Booz, G.W.; Sigmund, C.D.; Coffman, T.M.; Kawai, T.; Rizzo, V.; Scalia, R.; Eguchi, S. Angiotensin II Signal Transduction: An Update on Mechanisms of Physiology and Pathophysiology. Physiol. Rev. 2018, 98, 1627–1738. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | 5′ Primer | 3′ Primer |
|---|---|---|
| CBS | 5 atgctgcagaaaggcttcat 3 | 5 gtggaaaccagtcggtgtct 3 |
| CSE | 5 cgcacaaattgtccacaaac 3 | 5 gctctgtccttctcaggcac 3 |
| 3MST | 5 ggctcagtaaacatcccattc 3 | 5 tgtccttcacagggtcttcc 3 |
| DAO | 5 ccctttctggaaaagcacag 3 | 5 ctcctctcaccacctcttcg 3 |
| Renin | 5 aacattaccagggcaactttcact 3 | 5 acccccttcatggtgatctg 3 |
| PRR | 5 gaggcagtgaccctcaacat 3 | 5 ccctcctcacacaacaaggt 3 |
| ACE | 5 caccggcaaggtctgctt 3 | 5 cttggcatagtttcgtgaggaa 3 |
| ACE2 | 5 acccttcttacatcagccctactg 3 | 5 tgtccaaaacctaccccacatat 3 |
| AT1R | 5 gctgggcaacgagtttgtct 3 | 5 cagtccttcagctggatcttca 3 |
| AT2R | 5 caatctggctgtggctgactt 3 | 5 tgcacatcacaggtccaaaga 3 |
| MAS | 5catctctcctctcggctttgtg 3 | 5 cctcatccggaagcaaagg 3 |
| R18S | 5 gccgcggtaattccagctcca 3 | 5 cccgcccgctcccaagatc 3 |
| Groups | C | CKD | LC | DC |
|---|---|---|---|---|
| Mortality | 0% | 0% | 0% | 0% |
| Body weight (BW) (g) | 384 ± 7 | 367 ± 12 | 305 ± 9 *,# | 363 ± 6 |
| Left kidney weight (g) | 1.67 ± 0.05 | 1.62 ± 0.08 | 1.36 ± 0.06 *,# | 1.70 ± 0.06 |
| Left kidney weight/100 g BW | 0.43 ± 0.01 | 0.44 ± 0.01 | 0.44 ± 0.01 | 0.47 ± 0.01 |
| Systolic BP (mmHg) | 131 ± 1 | 143 ± 1 * | 132 ± 1 # | 132 ± 1 # |
| Diastolic BP (mmHg) | 91 ± 1 | 98 ± 2 * | 84 ± 2 # | 90 ± 2 # |
| Mean arterial pressure (mmHg) | 104 ± 1 | 113 ± 2 * | 100 ± 1 # | 104 ± 2 # |
| Groups | C | CKD | LC | DC |
|---|---|---|---|---|
| Tryptophan (ng/mL) | 22,856 ± 946 | 22,400 ± 1216 | 21,499 ± 611 | 20,778 ± 807 |
| Kynurenic acid (ng/mL) | 7.53 ± 0.61 | 5.46 ± 0.52 * | 8.64 ± 1.15 # | 7.2 ± 0.33 # |
| 3-HKN (ng/mL) | 6.77 ± 0.29 | 5.18 ± 0.22 * | 5.38 ± 0.19 * | 7.06 ± 0.31 # |
| Xanthurenic acid (ng/mL) | 3.71 ± 0.32 | 2.71 ± 0.21 * | 4.04 ± 0.67 | 3.52 ± 0.26 # |
| 3-HAA | 4.02 ± 0.4 | 3.01 ± 0.39 | 4.13 ± 0.41 | 1.45 ± 0.27 *,# |
| 5-HTP (ng/mL) | 6.42 ± 0.21 | 5.61 ± 0.23 * | 5.29 ± 0.28 * | 4.49 ± 0.17 *,# |
| Serotonin (ng/mL) | 329 ± 121 | 120 ± 18 | 26 ± 5 *# | 21 ± 10 *,# |
| 5-HIAA (ng/mL) | 16.3 ± 0.8 | 13.1 ± 0.7 * | 17.2 ± 0.8 # | 16.2 ± 1 # |
| N-AS (ng/mL) | 3.92 ± 0.79 | 2.76 ± 0.32 | 2.52 ± 0.51 | 3.3 ± 0.72 |
| IS (ng/mL) | 3066 ± 184 | 1974 ± 153 * | 2312 ± 189 * | 3285 ± 420 # |
| IAM (ng/mL) | 84.6 ± 4.1 | 69.4 ± 3 * | 70.3 ± 3.6 * | 81.6 ± 6.3 |
| ILA (ng/mL) | 282 ± 8 | 272 ± 13 | 262 ± 6 | 238 ± 6 *,# |
| ICA (ng/mL) | 2.73 ± 0.39 | 2.18 ± 0.24 | 4.26 ± 0.72 # | 5.45 ± 1.04 *,# |
| IAA (ng/mL) | 84.6 ± 4.1 | 69.4 ± 3 * | 70.3 ± 3.6 * | 81.6 ± 6.3 |
| Groups | C | CKD | LC | DC |
|---|---|---|---|---|
| l-Arginine (μM) | 355.3 ± 11.9 | 267.2 ± 8.5 * | 330.6 ± 14.3 # | 287.4 ± 37.5 |
| Asymmetric dimethylarginine (μM) | 2.15 ± 0.08 | 2.17 ± 0.13 | 1.89 ± 0.23 | 2.06 ± 0.25 |
| Symmetric dimethylarginine (μM) | 2.15 ± 0.11 | 2.39 ± 0.18 | 2.12 ± 0.14 | 1.92 ± 0.08 |
| l-Arginine-to-ADMA ratio (μM/μM) | 167.3 ± 8.7 | 125.5 ± 6.1 * | 197.8 ± 30.4 # | 156.6 ± 29.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, C.-N.; Hou, C.-Y.; Chang-Chien, G.-P.; Lin, S.; Tain, Y.-L. Dietary Supplementation with Cysteine during Pregnancy Rescues Maternal Chronic Kidney Disease-Induced Hypertension in Male Rat Offspring: The Impact of Hydrogen Sulfide and Microbiota-Derived Tryptophan Metabolites. Antioxidants 2022, 11, 483. https://doi.org/10.3390/antiox11030483
Hsu C-N, Hou C-Y, Chang-Chien G-P, Lin S, Tain Y-L. Dietary Supplementation with Cysteine during Pregnancy Rescues Maternal Chronic Kidney Disease-Induced Hypertension in Male Rat Offspring: The Impact of Hydrogen Sulfide and Microbiota-Derived Tryptophan Metabolites. Antioxidants. 2022; 11(3):483. https://doi.org/10.3390/antiox11030483
Chicago/Turabian StyleHsu, Chien-Ning, Chih-Yao Hou, Guo-Ping Chang-Chien, Sufan Lin, and You-Lin Tain. 2022. "Dietary Supplementation with Cysteine during Pregnancy Rescues Maternal Chronic Kidney Disease-Induced Hypertension in Male Rat Offspring: The Impact of Hydrogen Sulfide and Microbiota-Derived Tryptophan Metabolites" Antioxidants 11, no. 3: 483. https://doi.org/10.3390/antiox11030483
APA StyleHsu, C. -N., Hou, C. -Y., Chang-Chien, G. -P., Lin, S., & Tain, Y. -L. (2022). Dietary Supplementation with Cysteine during Pregnancy Rescues Maternal Chronic Kidney Disease-Induced Hypertension in Male Rat Offspring: The Impact of Hydrogen Sulfide and Microbiota-Derived Tryptophan Metabolites. Antioxidants, 11(3), 483. https://doi.org/10.3390/antiox11030483