
Effects of Nitrite Exposure on the Hematological Properties, Antioxidant and Stress Responses of Juvenile Hybrid Groupers, Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀


Abstract
:1. Introduction
2. Materials and Methods
2.1. Pre-Acute Nitrite Exposure Experiment
2.2. Experimental Fish
2.3. Blood Physiology
2.4. Antioxidant Responses
2.5. Stress Indicators
2.6. Statistical Analysis
2.7. Ethics Approval and Consent to Participate
3. Results
3.1. Blood Physiology
3.2. Antioxidant Responses
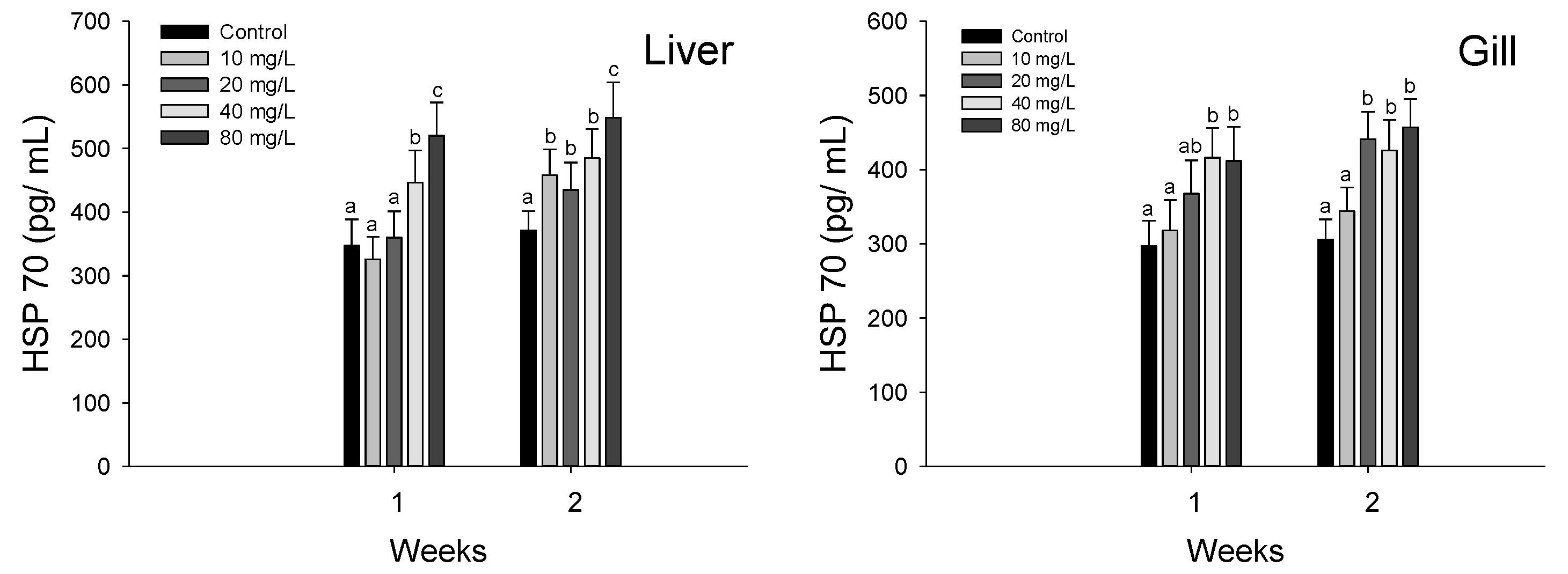
3.3. Stress Responses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gao, X.Q.; Fei, F.; Huo, H.H.; Huang, B.; Meng, X.S.; Zhang, T.; Liu, B.L. Effect of acute exposure to nitrite on physiological parameters, oxidative stress, and apoptosis in Takifugu rubripes. Ecotoxicol. Environ. Saf. 2020, 188, 109878. [Google Scholar] [CrossRef]
- Cheng, C.H.; Su, Y.L.; Ma, H.L.; Deng, Y.Q.; Feng, J.; Chen, X.L.; Jie, Y.K.; Guo, Z.X. Effect of nitrite exposure on oxidative stress, DNA damage and apoptosis in mud crab (Scylla paramamosain). Chemosphere 2020, 239, 124668. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, S.K.; Hur, Y.B. Toxic effects of waterborne nitrite exposure on antioxidant responses, acetylcholinesterase inhibition, and immune responses in olive flounders, Paralichthys olivaceus, reared in bio-floc and seawater. Fish Shellfish Immunol. 2020, 97, 581–586. [Google Scholar] [CrossRef]
- Gao, X.Q.; Fei, F.; Huo, H.H.; Huang, B.; Meng, X.S.; Zhang, T.; Liu, W.B.; Liu, B.L. Exposure to nitrite alters thyroid hormone levels and morphology in Takifugu rubripes. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 225, 108578. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, J.Y.; Lim, L.J.; Kim, S.K.; Choi, H.S.; Hur, Y.B. Effects of waterborne nitrite on hematological parameters and stress indicators in olive flounders, Paralichthys olivaceus, raised in bio-floc and seawater. Chemosphere 2018, 209, 28–34. [Google Scholar] [CrossRef]
- Zhang, W.; Li, J.; Chen, Y.; Si, Q.; Tian, J.; Jiang, Q.; Yang, J. Exposure time relevance of response to nitrite exposure: Insight from transcriptional responses of immune and antioxidant defense in the crayfish, Procambarus clarkii. Aquat. Toxicol. 2019, 214, 105262. [Google Scholar] [CrossRef] [PubMed]
- Kroupová, H.K.; Valentová, O.; Svobodová, Z.; Šauer, P.; Máchová, J. Toxic effects of nitrite on freshwater organisms: A review. Rev. Aquac. 2018, 10, 525–542. [Google Scholar] [CrossRef]
- Deane, E.E.; Woo, N.Y. Impact of nitrite exposure on endocrine, osmoregulatory and cytoprotective functions in the marine teleost Sparus sarba. Aquat. Toxicol. 2007, 82, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Grosell, M.; Jensen, F.B. Uptake and effects of nitrite in the marine teleost fish Platichthys flesus. Aquat. Toxicol. 2000, 50, 97–107. [Google Scholar] [CrossRef]
- Xiao, J.; Liu, Q.Y.; Du, J.H.; Zhu, W.L.; Li, Q.Y.; Chen, X.L.; Chen, X.H.; Liu, H.; Zhou, X.Y.; Zhao, Y.Z.; et al. Integrated analysis of physiological, transcriptomic and metabolomic responses and tolerance mechanism of nitrite exposure in Litopenaeus vannamei. Sci. Total Environ. 2020, 711, 134416. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.Q.; Fei, F.; Huo, H.H.; Huang, B.; Meng, X.S.; Zhang, T.; Liu, B.L. Impact of nitrite exposure on plasma biochemical parameters and immune-related responses in Takifugu rubripes. Aquat. Toxicol. 2020, 218, 105362. [Google Scholar] [CrossRef]
- Jensen, F.B.; Rohde, S. Comparative analysis of nitrite uptake and hemoglobin-nitrite reactions in erythrocytes: Sorting out uptake mechanisms and oxygenation dependencies. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R972–R982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pottinger, T.G. Modulation of the stress response in wild fish is associated with variation in dissolved nitrate and nitrite. Environ. Pollut. 2017, 225, 550–558. [Google Scholar] [CrossRef] [Green Version]
- Keszler, A.; Piknova, B.; Schechter, A.N.; Hogg, N. The Reaction between Nitrite and Oxyhemoglobin A Mechanistic Study. J. Biol. Chem. 2008, 283, 9615–9622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, R.; Han, C.; Lei, J.L.; Liu, B.L.; Huang, B.; Huo, H.H.; Yin, S.T. Effects of nitrite exposure on haematological parameters, oxidative stress and apoptosis in juvenile turbot (Scophthalmus maximus). Aquat. Toxicol. 2015, 169, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Xian, J.A.; Li, B.; Ye, C.X.; Wang, A.L.; Miao, Y.T.; Liao, S.A. Gene expression of apoptosis-related genes, stress protein and antioxidant enzymes in hemocytes of white shrimp Litopenaeus vannamei under nitrite stress. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2013, 157, 366–371. [Google Scholar] [CrossRef]
- Xian, J.A.; Wang, A.L.; Chen, X.D.; Gou, N.N.; Miao, Y.T.; Liao, S.A.; Ye, C.X. Cytotoxicity of nitrite on haemocytes of the tiger shrimp, Penaeus monodon, using flow cytometric analysis. Aquaculture 2011, 317, 240–244. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.K.; Hur, Y.B. Hematological parameters and antioxidant responses in olive flounder Paralichthys olivaceus in biofloc depend on water temperature. J. Therm. Biol. 2019, 82, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Cossu, C.; Doyotte, A.; Jacquin, M.C.; Babut, M.; Exinger, A.; Vasseur, P. Glutathione reductase, selenium-dependent glutathione peroxidase, glutathione levels, and lipid peroxidation in freshwater bivalves, Unio tumidus, as biomarkers of aquatic contamination in field studies. Ecotoxicol. Environ. Saf. 1997, 38, 122–131. [Google Scholar] [CrossRef]
- Lin, Y.; Miao, L.H.; Pan, W.J.; Huang, X.; Dengu, J.M.; Zhang, W.X.; Ge, X.P.; Liu, B.; Ren, M.C.; Zhou, Q.L.; et al. Effect of nitrite exposure on the antioxidant enzymes and glutathione system in the liver of bighead carp, Aristichthys nobilis. Fish Shellfish Immunol. 2018, 76, 126–132. [Google Scholar] [CrossRef]
- Lee, J.W.; Choi, H.; Hwang, U.K.; Kang, J.C.; Kang, Y.J.; Kim, K.I.; Kim, J.H. Toxic effects of lead exposure on bioaccumulation, oxidative stress, neurotoxicity, and immune responses in fish: A review. Environ. Toxicol. Pharmacol. 2019, 68, 101–108. [Google Scholar] [CrossRef]
- Das, P.C.; Ayyappan, S.; Das, B.K.; Jena, J.K. Nitrite toxicity in Indian major carps: Sublethal effect on selected enzymes in fingerlings of Catla catla, Labeo rohita and Cirrhinus mrigala. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2004, 138, 3–10. [Google Scholar] [CrossRef]
- Mormède, P.; Andanson, S.; Aupérin, B.; Beerda, B.; Guémené, D.; Malmkvist, J.; Manteca, X.; Manteuffel, G.; Prunet, P.; van Reenen, C.G.; et al. Exploration of the hypothalamic–pituitary–adrenal function as a tool to evaluate animal welfare. Physiol. Behav. 2007, 92, 317–339. [Google Scholar] [CrossRef] [PubMed]
- Franco-Martinez, L.; Tvarijonaviciute, A.; Martinez-Subiela, S.; Teles, M.; Tort, L. Chemiluminescent assay as an alternative to radioimmunoassay for the measurement of cortisol in plasma and skin mucus of Oncorhynchus mykiss. Ecol. Indic. 2019, 98, 634–640. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. The immune responses and expression of metallothionein (MT) gene and heat shock protein 70 (HSP 70) in juvenile rockfish, Sebastes schlegelii, exposed to waterborne arsenic (As3+). Environ. Toxicol. Pharmacol. 2016, 47, 136–141. [Google Scholar] [CrossRef]
- Cho, J.H.; Kim, S.R.; Hur, Y.B.; Lee, K.M.; Kim, J.H. Tolerance limit of nitrite exposure to hybrid grouper (Epinephelus fuscoguttatus ♀× E. lanceolatus ♂): Hematological parameters and plasma components. Korean J. Environ. Biol. 2020, 38, 93–100. [Google Scholar] [CrossRef]
- Kim, J.H.; Cho, J.H.; Kim, S.R.; Hur, Y.B. Toxic effects of waterborne ammonia exposure on hematological parameters, oxidative stress and stress indicators of juvenile hybrid grouper, Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀. Environ. Toxicol. Pharmacol. 2020, 80, 103453. [Google Scholar] [CrossRef]
- Kim, J.H.; Oh, C.W.; Kang, J.C. Antioxidant responses, neurotoxicity, and metallothionein gene expression in juvenile Korean rockfish Sebastes schlegelii under dietary lead exposure. J. Aquat. Anim. Health 2017, 29, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Jensen, F.B. Nitrite disrupts multiple physiological functions in aquatic animals. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 135, 9–24. [Google Scholar] [CrossRef]
- Madison, B.N.; Wang, Y.S. Haematological responses of acute nitrite exposure in walleye (Sander vitreus). Aquat. Toxicol. 2006, 79, 16–23. [Google Scholar] [CrossRef]
- Da Costa, O.T.F.; dos Santos Ferreira, D.J.; Mendonça, F.L.P.; Fernandes, M.N. Susceptibility of the Amazonian fish, Colossoma macropomum (Serrasalminae), to short-term exposure to nitrite. Aquaculture 2004, 232, 627–636. [Google Scholar] [CrossRef]
- Avilez, I.M.; Altran, A.E.; Aguiar, L.H.; Moraes, G. Hematological responses of the Neotropical teleost matrinxa (Brycon cephalus) to environmental nitrite. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2004, 139, 135–139. [Google Scholar] [CrossRef]
- Yildiz, H.Y.; Köksal, G.; Borazan, G.; Benli, C.K. Nitrite-induced methemoglobinemia in Nile tilapia, Oreochromis niloticus. J. Appl. Ichthyol. 2006, 22, 426–427. [Google Scholar] [CrossRef]
- Huertas, M.; Gisbert, E.; Rodrıguez, A.; Cardona, L.; Williot, P.; Castello-Orvay, F. Acute exposure of Siberian sturgeon (Acipenser baeri, Brandt) yearlings to nitrite: Median-lethal concentration (LC50) determination, haematological changes and nitrite accumulation in selected tissues. Aquat. Toxicol. 2002, 57, 257–266. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.K.; Kim, J.H. Bio-floc technology application in flatfish Paralichthys olivaceus culture: Effects on water quality, growth, hematological parameters, and immune responses. Aquaculture 2018, 495, 703–709. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, H.; Sung, G.H.; Seo, S.A.; Kim, K.I.; Kang, Y.J.; Kang, J.C. Toxic effects on hematological parameters and oxidative stress in juvenile olive flounder, Paralichthys olivaceus exposed to waterborne zinc. Aquac. Rep. 2019, 15, 100225. [Google Scholar] [CrossRef]
- Ciji, A.; Sahu, N.P.; Pal, A.K.; Akhtar, M.S. Dietary L-tryptophan modulates growth and immuno-metabolic status of Labeo rohita juveniles exposed to nitrite. Aquac. Res. 2015, 46, 2013–2024. [Google Scholar] [CrossRef]
- Das, P.C.; Ayyappan, S.; Jena, J.K.; Das, B.K. Nitrite toxicity in Cirrhinus mrigala (Ham.): Acute toxicity and sub-lethal effect on selected haematological parameters. Aquaculture 2004, 235, 633–644. [Google Scholar] [CrossRef]
- Jia, R.; Liu, B.L.; Han, C.; Huang, B.; Lei, J.L. The physiological performance and immune response of juvenile turbot (Scophthalmus maximus) to nitrite exposure. Comp. Biochem. Physiol. Part C 2016, 181–182, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Tomasso, J.R. Environmental nitrite and aquaculture: A perspective. Aquac. Int. 2012, 20, 1107–1116. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. Effects of sub-chronic exposure to lead (Pb) and ascorbic acid in juvenile rockfish: Antioxidant responses, MT gene expression, and neurotransmitters. Chemosphere 2017, 171, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kang, Y.J.; Kim, K.I.; Kim, S.K.; Kim, J.H. Toxic effects on nitrogenous compounds (ammonia, nitrite, and nitrate) on acute toxicity and antioxidant responses of juvenile olive flounder, Paralichthys olivaceus. Environ. Toxicol. Pharmacol. 2019, 67, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Ruan, G.; Fan, W.; Fang, L.; Wang, Q.; Luo, M.; Yi, T. The effect of nitrite and sulfide on the antioxidant capacity and microbial composition of the intestines of red swamp crayfish, Procambarus clarkii. Fish Shellfish Immunol. 2020, 96, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.Y.; An, K.W.; An, M.I. Molecular characterization and mRNA expression of glutathione peroxidase and glutathione S-transferase during osmotic stress in olive flounder (Paralichthys olivaceus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 149, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kang, J.C. Oxidative stress, neurotoxicity, and metallothionein (MT) gene expression in juvenile rock fish Sebastes schlegelii under the different levels of dietary chromium (Cr6+) exposure. Ecotoxicol. Environ. Saf. 2016, 125, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.A.R.; El-Houseiny, W.; El-Murr, A.E.; Ebraheim, L.L.; Ahmed, A.I.; El-Hakim, Y.M.A. Effect of hexavalent chromium exposure on the liver and kidney tissues related to the expression of CYP450 and GST genes of Oreochromis niloticus fish: Role of curcumin supplemented diet. Ecotoxicol. Environ. Saf. 2020, 188, 109890. [Google Scholar] [CrossRef]
- Rahman, I.; MacNee, W. Regulation of redox glutathione levels and gene transcription in lung inflammation: Therapeutic approaches. Free Radic. Biol. Med. 2000, 28, 1405–1420. [Google Scholar] [CrossRef]
- Lin, W.; Hou, J.; Guo, H.; Li, L.; Wang, L.; Zhang, D.; Li, D.; Tang, R. The synergistic effects of waterborne microcystin-LR and nitrite on hepatic pathological damage, lipid peroxidation and antioxidant responses of male zebrafish. Environ. Pollut. 2018, 235, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Ciji, A.; Sahu, N.P.; Pal, A.K.; Akhtar, M.S. Physiological changes in Labeo rohita during nitrite exposure: Detoxification through dietary vitamin E. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2013, 158, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Park, H.J.; Kang, J.C. Alterations in growth performance and stress responses in juvenile rockfish, Sebastes schlegelii, exposed to dietary chromium with varying levels of dietary ascorbic acid supplementation. Chemosphere 2017, 189, 672–678. [Google Scholar] [CrossRef]
- Jiang, J.; Shi, Y.; Shan, Z.; Yang, L.; Wang, X.; Shi, L. Bioaccumulation, oxidative stress and HSP70 expression in Cyprinus carpio L. exposed to microcystin-LR under laboratory conditions. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2012, 155, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.D.; Caron, A.W.; Bourget, L.; Meriin, A.B.; Sherman, M.Y.; Morimoto, R.I.; Massie, B. The chaperone function of hsp70 is required for protection against stress-induced apoptosis. Mol. Cell. Biol. 2000, 20, 7146–7159. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 95% Confidence Limits | |
|---|---|
| Probability | Estimate (mg/L) |
| 0.01 | 538.39 |
| 0.10 | 681.39 |
| 0.20 | 741.60 |
| 0.30 | 785.02 |
| 0.40 | 822.11 |
| 0.50 | 856.79 |
| 0.60 | 891.46 |
| 0.70 | 928.56 |
| 0.80 | 971.97 |
| 0.90 | 1032.19 |
| 0.99 | 1175.18 |
| Item | Value (Mean ± SD) |
|---|---|
| Temperature (°C) | 24.8 ± 0.5 |
| pH | 8.38 ± 0.11 |
| Salinity (‰) | 31.9 ± 0.3 |
| Dissolved Oxygen (mg/L) | 7.48 ± 0.24 |
| Ammonia (mg/L) | 0.58 ± 0.13 |
| Nitrite (mg/L) | 0.06 ± 0.02 |
| Nitrate (mg/L) | 1.12 ± 0.15 |
| Waterborne Nitrite Concentration (NO2− mg/L) | |||||
|---|---|---|---|---|---|
| Waterborne nitrite levels | 0 | 10 | 20 | 40 | 80 |
| Measured nitrite levels | 0.07 | 10.5 | 21.2 | 43.9 | 83.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-H.; Kang, Y.J.; Lee, K.M. Effects of Nitrite Exposure on the Hematological Properties, Antioxidant and Stress Responses of Juvenile Hybrid Groupers, Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀. Antioxidants 2022, 11, 545. https://doi.org/10.3390/antiox11030545
Kim J-H, Kang YJ, Lee KM. Effects of Nitrite Exposure on the Hematological Properties, Antioxidant and Stress Responses of Juvenile Hybrid Groupers, Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀. Antioxidants. 2022; 11(3):545. https://doi.org/10.3390/antiox11030545
Chicago/Turabian StyleKim, Jun-Hwan, Yue Jai Kang, and Kyung Mi Lee. 2022. "Effects of Nitrite Exposure on the Hematological Properties, Antioxidant and Stress Responses of Juvenile Hybrid Groupers, Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀" Antioxidants 11, no. 3: 545. https://doi.org/10.3390/antiox11030545
APA StyleKim, J. -H., Kang, Y. J., & Lee, K. M. (2022). Effects of Nitrite Exposure on the Hematological Properties, Antioxidant and Stress Responses of Juvenile Hybrid Groupers, Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀. Antioxidants, 11(3), 545. https://doi.org/10.3390/antiox11030545