
Ornipural® Mitigates Malathion-Induced Hepato-Renal Damage in Rats via Amelioration of Oxidative Stress Biomarkers, Restoration of Antioxidant Activity, and Attenuation of Inflammatory Response


 ,
,  , and
, and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Experimental Animals, Treatment Protocol, and Ethical Considerations
2.3. Assessment of Hepato-Renal Functional and Metabolic Serum Parameters
2.4. Investigation of Hepato-Renal Oxidant and Antioxidant Tissue Parameters
2.5. Assay of Pro-Inflammatory Cytokines Gene Expression in Livers and Kidneys
2.6. Histopathological Examination
2.7. Statistical Analysis
3. Results
3.1. Performance and Relative Weight of Livers and Kidneys
3.2. Hepato-Renal Functional and Metabolic Serum Parameters
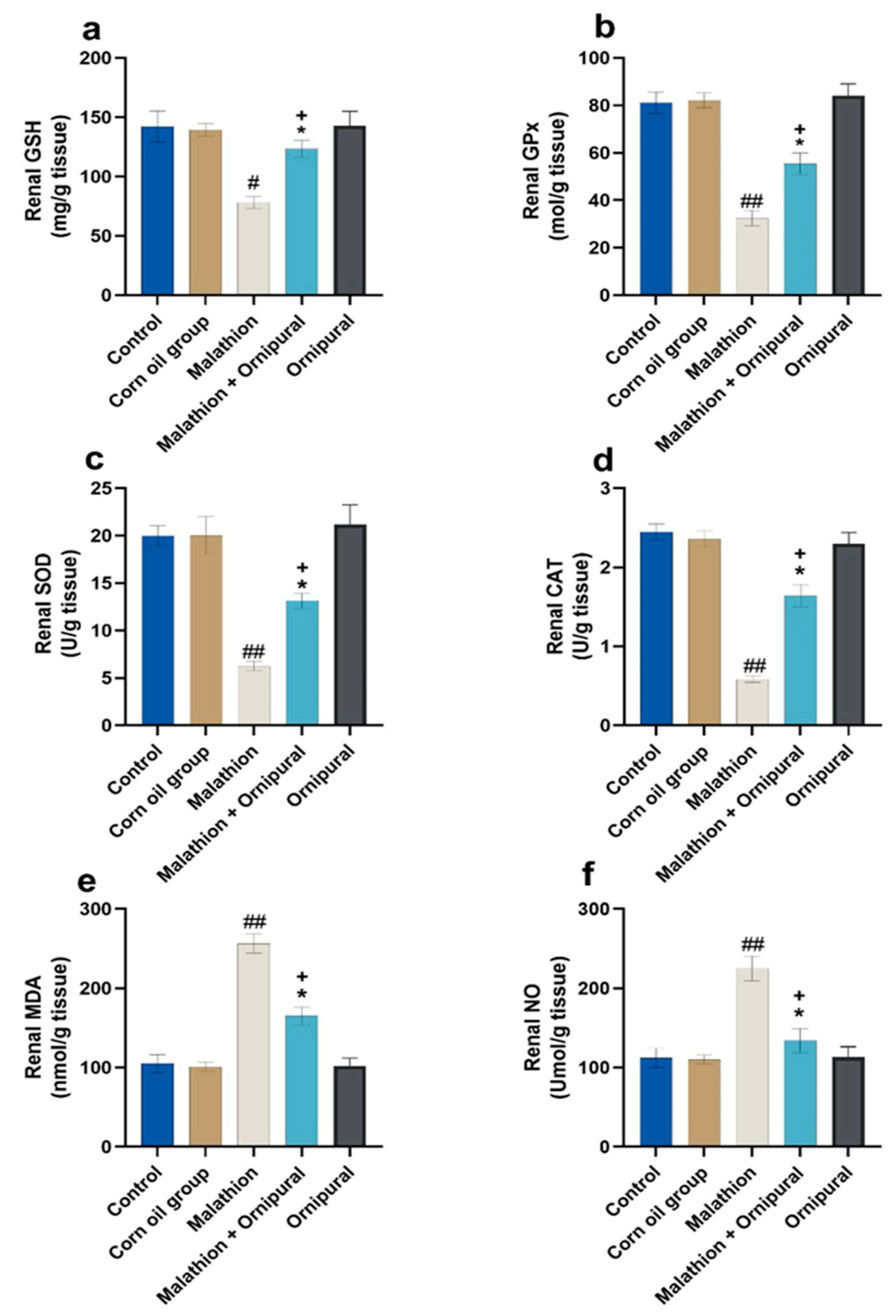
3.3. Evaluation of Oxidative Stress in Liver and Kidney Tissue
3.4. Inflammatory Cytokines Genes Expression
3.5. Changes on 8-Hydroxydeoxyguanosine (8-OHdG) and TNF-Alpha
3.6. Histopathological Findings
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Selmi, S.; Rtibi, K.; Grami, D.; Sebai, H.; Marzouki, L. Malathion, an organophosphate insecticide, provokes metabolic, histopathologic and molecular disorders in liver and kidney in prepubertal male mice. Toxicol. Rep. 2018, 5, 189–195. [Google Scholar] [CrossRef]
- Zidan, N.E.-H.A. Hepato-and nephrotoxicity in male albino rats exposed to malathion and spinosad in stored wheat grains. Acta Biol. Hung. 2015, 66, 133–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badr, A.M. Organophosphate toxicity: Updates of malathion potential toxic effects in mammals and potential treatments. Environ. Sci. Pollut. Res. 2020, 27, 26036–26057. [Google Scholar] [CrossRef] [PubMed]
- Fortunato, J.J.; Feier, G.; Vitali, A.M.; Petronilho, F.C.; Dal-Pizzol, F.; Quevedo, J. Malathion-induced oxidative stress in rat brain regions. Neurochem. Res. 2006, 31, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Varol, S.; Başarslan, S.; Fırat, U.; Alp, H.; Uzar, E.; Arıkanoğlu, A.; Evliyaoğlu, O.; Acar, A.; Yücel, Y.; Kıbrıslı, E. Detection of borderline dosage of malathion intoxication in a rat’s brain. Eur. Rev. Med. Pharm. Sci. 2015, 19, 2318–2323. [Google Scholar]
- Del-Rahman, A.; Dechkovskaia, A.M.; Goldstein, L.B.; Bullman, S.H.; Khan, W.; El-Masry, E.M.; Abou-Donia, M.B. Neurological deficits induced by malathion, DEET, and permethrin, alone or in combination in adult rats. J. Toxicol. Environ. Health Part A 2004, 67, 331–356. [Google Scholar] [CrossRef]
- Dos Santos, A.A.; dos Santos, D.B.; Ribeiro, R.P.; Colle, D.; Peres, K.C.; Hermes, J.; Barbosa, A.M.; Dafré, A.L.; de Bem, A.F.; Kuca, K. Effects of K074 and pralidoxime on antioxidant and acetylcholinesterase response in malathion-poisoned mice. Neurotoxicology 2011, 32, 888–895. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, A.A.; Naime, A.A.; de Oliveira, J.; Colle, D.; Dos Santos, D.B.; Hort, M.A.; Moreira, E.L.G.; Suñol, C.; de Bem, A.F.; Farina, M. Long-term and low-dose malathion exposure causes cognitive impairment in adult mice: Evidence of hippocampal mitochondrial dysfunction, astrogliosis and apoptotic events. Arch. Toxicol. 2016, 90, 647–660. [Google Scholar] [CrossRef]
- Navarrete-Meneses, M.; Salas-Labadía, C.; Sanabrais-Jiménez, M.; Santana-Hernández, J.; Serrano-Cuevas, A.; Juárez-Velázquez, R.; Olaya-Vargas, A.; Pérez-Vera, P. Exposure to the insecticides permethrin and malathion induces leukemia and lymphoma-associated gene aberrations in vitro. Toxicol. Vitr. 2017, 44, 17–26. [Google Scholar] [CrossRef]
- Waheed, S.; Halsall, C.; Sweetman, A.J.; Jones, K.C.; Malik, R.N. Pesticides contaminated dust exposure, risk diagnosis and exposure markers in occupational and residential settings of Lahore, Pakistan. Environ. Toxicol. Pharmacol. 2017, 56, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Selmi, S.; El-Fazaa, S.; Gharbi, N. Oxidative stress and cholinesterase inhibition in plasma, erythrocyte and brain of rats’ pups following lactational exposure to malathion. Environ. Toxicol. Pharmacol. 2012, 34, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Baiomy, A.A.; Attia, H.F.; Soliman, M.M.; Makrum, O. Protective effect of ginger and zinc chloride mixture on the liver and kidney alterations induced by malathion toxicity. Int. J. Immunopathol. Pharmacol. 2015, 28, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Xiang, B.; Wang, D.; Tang, S.; Teng, M.; Yan, S.; Zhou, Z.; Zhu, W. Different toxic effects of racemate, enantiomers, and metabolite of malathion on HepG2 cells using high-performance liquid chromatography–quadrupole–time-of-flight-based metabolomics. J. Agric. Food Chem. 2019, 67, 1784–1794. [Google Scholar] [CrossRef] [PubMed]
- Shieh, P.; Jan, C.-R.; Liang, W.-Z. The protective effects of the antioxidant N-acetylcysteine (NAC) against oxidative stress-associated apoptosis evoked by the organophosphorus insecticide malathion in normal human astrocytes. Toxicology 2019, 417, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, J.K.; Saraf, P.; Kumari, P.; Mittal, M.; Kumar, V. N-Acetyl-cysteine mediated inhibition of spermatogonial cells apoptosis against malathion exposure in testicular tissue. J. Biochem. Mol. Toxicol. 2018, 32, e22046. [Google Scholar] [CrossRef]
- Abdel-Salam, O.M.; Youness, E.R.; Mohammed, N.A.; Yassen, N.N.; Khadrawy, Y.A.; El-Toukhy, S.E.; Sleem, A.A. Nitric oxide synthase inhibitors protect against brain and liver damage caused by acute malathion intoxication. Asian Pac. J. Trop. Med. 2017, 10, 773–786. [Google Scholar] [CrossRef]
- Akbel, E.; Arslan-Acaroz, D.; Demirel, H.H.; Kucukkurt, I.; Ince, S. The subchronic exposure to malathion, an organophosphate pesticide, causes lipid peroxidation, oxidative stress, and tissue damage in rats: The protective role of resveratrol. Toxicol. Res. 2018, 7, 503–512. [Google Scholar] [CrossRef] [Green Version]
- Selmi, S.; El-Fazaa, S.; Gharbi, N. Oxidative stress and alteration of biochemical markers in liver and kidney by malathion in rat pups. Toxicol. Ind. Health 2015, 31, 783–788. [Google Scholar] [CrossRef]
- Ramadan, G.; El-Beih, N.M.; Ahmed, R.S. Aged garlic extract ameliorates immunotoxicity, hematotoxicity and impaired burn-healing in malathion-and carbaryl-treated male albino rats. Environ. Toxicol. 2017, 32, 789–798. [Google Scholar] [CrossRef]
- World Health Organization. The World Health Report 2006: Working Together for Health; World Health Organization: Geneva, Switzerland, 2006.
- World Health Organization. International Classification of Functioning, Disability, and Health: Children & Youth Version: ICF-CY; World Health Organization: Geneva, Switzerland, 2007.
- Kiely, T.; Donaldson, D.; Grube, A. Pesticides Industry Sales and Usage: 2000 and 2001 Market Estimates; US Environmental Protection Agency: Washington, DC, USA, 2004; p. 114.
- Oneta, C.M. Non-alcoholic fatty liver disease: Treatment options based on pathogenic considerations. Swiss Med. Wkly. 2002, 132, 493–505. [Google Scholar]
- Frame, L.A.; Costa, E.; Jackson, S.A. Current explorations of nutrition and the gut microbiome: A comprehensive evaluation of the review literature. Nutr. Rev. 2020, 78, 798–812. [Google Scholar] [CrossRef] [PubMed]
- Breuillard, C.; Cynober, L.; Moinard, C. Citrulline and nitrogen homeostasis: An overview. Amino Acids 2015, 47, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.M.; Walser, M. Short term regulation of ureagenesis. J. Biol. Chem. 1980, 255, 5270–5280. [Google Scholar] [CrossRef]
- Uzun, F.G.; Kalender, S.; Durak, D.; Demir, F.; Kalender, Y. Malathion-induced testicular toxicity in male rats and the protective effect of vitamins C and E. Food Chem. Toxicol. 2009, 47, 1903–1908. [Google Scholar] [CrossRef] [PubMed]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Tietz, N.; Burtis, C.; Duncan, P.; Ervin, K.; Petitclerc, C.; Rinker, A.; Shuey, D.; Zygowicz, E. A reference method for measurement of alkaline phosphatase activity in human serum. Clin. Chem. 1983, 29, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Doumas, B.; Baysa, D.; Carter, R.; Peters, T.; Schaffer, R. Determination of serum total protein. Clin. Chem. 1981, 27, 1642. [Google Scholar] [CrossRef] [PubMed]
- Coulombe, J.J.; Favreau, L. A new simple semimicro method for colorimetric determination of urea. Clin. Chem. 1963, 9, 102–108. [Google Scholar] [CrossRef]
- Bartels, H. Serum creatinine without interference. Clin. Chem. Acta 1972, 37, 193–197. [Google Scholar] [CrossRef]
- CARAWAY, W.T.; HALD, P.M. Uric acid. In Standard Methods of Clinical Chemistry; Elsevier: Amsterdam, The Netherlands, 1963; Volume 4, pp. 239–247. [Google Scholar]
- Lopes-Virella, M.F.; Stone, P.G.; Colwell, J.A. Serum high density lipoprotein in diabetic patients. Diabetologia 1977, 13, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Lasram, M.M.; Annabi, A.B.; Rezg, R.; Elj, N.; Slimen, S.; Kamoun, A.; El-Fazaa, S.; Gharbi, N. Effect of short-time malathion administration on glucose homeostasis in Wistar rat. Pestic. Biochem. Physiol. 2008, 92, 114–119. [Google Scholar] [CrossRef]
- Nili-Ahmadabadi, A.; Pourkhalili, N.; Fouladdel, S.; Pakzad, M.; Mostafalou, S.; Hassani, S.; Baeeri, M.; Azizi, E.; Ostad, S.N.; Hosseini, R. On the biochemical and molecular mechanisms by which malathion induces dysfunction in pancreatic islets in vivo and in vitro. Pestic. Biochem. Physiol. 2013, 106, 51–60. [Google Scholar] [CrossRef]
- Lasram, M.M.; Dhouib, I.B.; Bouzid, K.; Lamine, A.J.; Annabi, A.; Belhadjhmida, N.; Ahmed, M.B.; El Fazaa, S.; Abdelmoula, J.; Gharbi, N. Association of inflammatory response and oxidative injury in the pathogenesis of liver steatosis and insulin resistance following subchronic exposure to malathion in rats. Environ. Toxicol. Pharmacol. 2014, 38, 542–553. [Google Scholar] [CrossRef]
- Khalifa, F.K.; Alkhalaf, M.I. Effects of black seed and thyme leaves dietary supplements against malathion insecticide-induced toxicity in experimental rat model. J. King Saud Univ.-Sci. 2020, 32, 914–919. [Google Scholar] [CrossRef]
- Jalili, C.; Roshankhah, S.; Moradi, Y.; Salahshoor, M.R. Resveratrol attenuates malathion-induced renal damage by declining oxidative stress in rats. Int. J. Pharm. Investig. 2018, 8, 192–199. [Google Scholar]
- Esen, M.; Uysal, M. Protective effects of intravenous lipid emulsion on malathion-induced hepatotoxicity. Bratisl. Lek. Listy 2018, 119, 373–378. [Google Scholar] [CrossRef]
- Moore, P.D.; Patlolla, A.K.; Tchounwou, P.B. Cytogenetic evaluation of malathion-induced toxicity in Sprague-Dawley rats. Mutat. Res./Genet. Toxicol. Environ. Mutagenes. 2011, 725, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Buratti, F.M.; D’aniello, A.; Volpe, M.T.; Meneguz, A.; Testai, E. Malathion bioactivation in the human liver: The contribution of different cytochrome P450 isoforms. Drug Metab. Dispos. 2005, 33, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Shedid, S.M.; Abdel-Magied, N.; Saada, H.N. Role of betaine in liver injury induced by the exposure to ionizing radiation. Environ. Toxicol. 2019, 34, 123–130. [Google Scholar] [CrossRef]
- Elbassuoni, E.A.; Ragy, M.M.; Ahmed, S.M. Evidence of the protective effect of l-arginine and vitamin D against monosodium glutamate-induced liver and kidney dysfunction in rats. Biomed. Pharmacother. 2018, 108, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Clark, T.; Tinsley, J.; Sigholt, T.; Macqueen, D.; Martin, S. Supplementation of arginine, ornithine and citrulline in rainbow trout (Oncorhynchus mykiss): Effects on growth, amino acid levels in plasma and gene expression responses in liver tissue. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 241, 110632. [Google Scholar] [CrossRef] [PubMed]
- Vong, L.B.; Ibayashi, Y.; Lee, Y.; Ngo, D.-N.; Nishikawa, Y.; Nagasaki, Y. Poly (ornithine)-based self-assembling drug for recovery of hyperammonemia and damage in acute liver injury. J. Control. Release 2019, 310, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Rashid, J.; Kumar, S.S.; Job, K.M.; Liu, X.; Fike, C.D.; Sherwin, C.M. Therapeutic potential of citrulline as an arginine supplement: A clinical pharmacology review. Pediatric Drugs 2020, 22, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Lie, A.H.; Chandra-Hioe, M.V.; Arcot, J. Sorbitol enhances the physicochemical stability of B12 vitamers. Int. J. Vitam. Nutr. Res. 2019, 90, 1–9. [Google Scholar]
- Umeda, M.; Hiramoto, M.; Watanabe, A.; Tsunoda, N.; Imai, T. Arginine-induced insulin secretion in endoplasmic reticulum. Biochem. Biophys. Res. Commun. 2015, 466, 717–722. [Google Scholar] [CrossRef] [Green Version]
- Kanaley, J.A. Growth hormone, arginine and exercise. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 50–54. [Google Scholar] [CrossRef]
- Day, C.R.; Kempson, S.A. Betaine chemistry, roles, and potential use in liver disease. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2016, 1860, 1098–1106. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Qi, Y.; ALuo, Z.; Liu, S.; Zhang, Z.; Zhou, L. Betaine increases mitochondrial content and improves hepatic lipid metabolism. Food Funct. 2019, 10, 216–223. [Google Scholar] [CrossRef]
- Davies, N.A.; Wright, G.; Ytrebø, L.M.; Stadlbauer, V.; Fuskevåg, O.M.; Zwingmann, C.; Davies, D.C.; Habtesion, A.; Hodges, S.J.; Jalan, R. L-ornithine and phenylacetate synergistically produce sustained reduction in ammonia and brain water in cirrhotic rats. Hepatology 2009, 50, 155–164. [Google Scholar] [CrossRef]
- Paßlack, N.; Zentek, J. Effects of dietary arginine, ornithine, and zeolite supplementation on uremic toxins in cats. Toxins 2018, 10, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassanpour, S.; Rezaei, H.; Razavi, S.M. Anti-nociceptive and antioxidant activity of betaine on formalin-and writhing tests induced pain in mice. Behav. Brain Res. 2020, 390, 112699. [Google Scholar] [CrossRef]
- Shin, S.; Gombedza, F.C.; Bandyopadhyay, B.C. L-ornithine activates Ca2+ signaling to exert its protective function on human proximal tubular cells. Cell. Signal. 2020, 67, 109484. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wen, S.; Zhou, J.; Ding, S. Association between malnutrition and hyperhomocysteine in Alzheimer’s disease patients and diet intervention of betaine. J. Clin. Lab. Anal. 2017, 31, e22090. [Google Scholar] [CrossRef] [PubMed]
- Nobari, H.; Cholewa, J.M.; Pérez-Gómez, J.; Castillo-Rodríguez, A. Effects of 14-weeks betaine supplementation on pro-inflammatory cytokines and hematology status in professional youth soccer players during a competition season: A double blind, randomized, placebo-controlled trial. J. Int. Soc. Sports Nutr. 2021, 18, 42. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, M.; Watari, H.; AbuAlmaaty, A.; Ohba, Y.; Sakuragi, N. Apoptosis and molecular targeting therapy in cancer. BioMed. Res. Int. 2014, 2014, 150845. [Google Scholar] [CrossRef] [Green Version]
- Doğru-Abbasoğlu, S.; Soluk-Tekkeşin, M.; Olgaç, V.; Uysal, M. Effect of betaine treatment on the regression of existing hepatic triglyceride accumulation and oxidative stress in rats fed on high fructose diet. Gen. Physiol. Biophys. 2018, 37, 563–570. [Google Scholar]
- Heidari, R.; Niknahad, H.; Sadeghi, A.; Mohammadi, H.; Ghanbarinejad, V.; Ommati, M.M.; Hosseini, A.; Azarpira, N.; Khodaei, F.; Farshad, O. Betaine treatment protects liver through regulating mitochondrial function and counteracting oxidative stress in acute and chronic animal models of hepatic injury. Biomed. Pharmacother. 2018, 103, 75–86. [Google Scholar] [CrossRef]
- Acquaviva, R.; Lanteri, R.; Li Destri, G.; Caltabiano, R.; Vanella, L.; Lanzafame, S.; Di Cataldo, A.; Li Volti, G.; Di Giacomo, C. Beneficial effects of rutin and L-arginine coadministration in a rat model of liver ischemia-reperfusion injury. Am. J. Physiol.-Gastrointest. Liver Physiol. 2009, 296, G664–G670. [Google Scholar] [CrossRef] [Green Version]
- Aziz, N.; Kamel, M.; Rifaai, R. Effects of hemin, a heme oxygenase-1 inducer in L-arginine-induced acute pancreatitis and associated lung injury in adult male albino rats. Endocr. Regul. 2017, 51, 20–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Bazer, F.W.; Davis, T.A.; Kim, S.W.; Li, P.; Marc Rhoads, J.; Carey Satterfield, M.; Smith, S.B.; Spencer, T.E.; Yin, Y. Arginine metabolism and nutrition in growth, health and disease. Amino Acids 2009, 37, 153–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorente, J.A.; Delgado, M.A.; Tejedor, C.; Mon, E.; Hervás, M.; Pascual, T.; Fernández-Segoviano, P.; Rieppi, G.; Soler, A.; Ayuso, D. Modulation of systemic hemodynamics by exogenous L-arginine in normal and bacteremic sheep. Crit. Care Med. 1999, 27, 2474–2479. [Google Scholar] [CrossRef] [PubMed]
- Wesseling, S.; Joles, J.A.; van Goor, H.; Bluyssen, H.A.; Kemmeren, P.; Holstege, F.C.; Koomans, H.A.; Braam, B. Transcriptome-based identification of pro-and antioxidative gene expression in kidney cortex of nitric oxide-depleted rats. Physiol. Genom. 2007, 28, 158–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wileman, S.M.; Mann, G.E.; Pearson, J.D.; Baydoun, A.R. Role of L-citrulline transport in nitric oxide synthesis in rat aortic smooth muscle cells activated with LPS and interferon-γ. Br. J. Pharmacol. 2003, 140, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Kim, W.S.; Rubakhin, S.S.; Sweedler, J.V. Ubiquitous presence of argininosuccinate at millimolar levels in the central nervous system of Aplysia californica. J. Neurochem. 2007, 101, 632–640. [Google Scholar] [CrossRef]
- Datta, P.K.; Gross, E.J.; Lianos, E.A. Interactions between inducible nitric oxide synthase and heme oxygenase-1 in glomerulonephritis. Kidney Int. 2002, 61, 847–850. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Yang, J.I.; Jung, M.H.; Hwa, J.S.; Kang, K.-R.; Park, D.J.; Roh, G.S.; Cho, G.J.; Choi, W.S.; Chang, S.-H. Heme oxygenase-1 protects rat kidney from ureteral obstruction via an anti-apoptotic pathway. J. Am. Soc. Nephrol. 2006, 17, 1373–1381. [Google Scholar] [CrossRef] [Green Version]
- Ramana, K.V.; Chandra, D.; Srivastava, S.; Bhatnagar, A.; Srivastava, S.K. Nitric oxide regulates the polyol pathway of glucose metabolism in vascular smooth muscle cells. FASEB J. 2003, 17, 417–425. [Google Scholar] [CrossRef]
- Bonnefont-Rousselot, D. Glucose and reactive oxygen species. Curr. Opin. Clin. Nutr. Metab. Care 2002, 5, 561–568. [Google Scholar] [CrossRef]
- Obrosova, I.G. Increased sorbitol pathway activity generates oxidative stress in tissue sites for diabetic complications. Antioxid. Redox Signal. 2005, 7, 1543–1552. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M. Glycation products and the pathogenesis of diabetic complications. Diabetes Care 1992, 15, 1835–1843. [Google Scholar] [CrossRef] [PubMed]
- Son, H.-Y.; Kim, H.; Kwon, Y.H. Taurine prevents oxidative damage of high glucose-induced cataractogenesis in isolated rat lenses. J. Nutr. Sci. Vitaminol. 2007, 53, 324–330. [Google Scholar] [CrossRef] [Green Version]
- Mongkhon, J.-M.; Thach, M.; Shi, Q.; Fernandes, J.C.; Fahmi, H.; Benderdour, M. Sorbitol-modified hyaluronic acid reduces oxidative stress, apoptosis and mediators of inflammation and catabolism in human osteoarthritic chondrocytes. Inflamm. Res. 2014, 63, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.-W.; Kwak, S.-H.; Yun, S.-Y.; Kim, S.-K. Evaluation of antioxidant activity of sugar alcohols using TOSC (total oxy-radical scavenging capacity) assay. Toxicol. Res. 2007, 23, 143–150. [Google Scholar] [CrossRef]
- Chukwuma, C.I.; Islam, M.S. Sorbitol increases muscle glucose uptake ex vivo and inhibits intestinal glucose absorption ex vivo and in normal and type 2 diabetic rats. Appl. Physiol. Nutr. Metab. 2017, 42, 377–383. [Google Scholar] [CrossRef]
- Livesey, G. Health potential of polyols as sugar replacers, with emphasis on low glycaemic properties. Nutr. Res. Rev. 2003, 16, 163–191. [Google Scholar] [CrossRef] [Green Version]
- Akgün, S.; Ertel, N.H. A comparison of carbohydrate metabolism after sucrose, sorbitol, and fructose meals in normal and diabetic subjects. Diabetes Care 1980, 3, 582–585. [Google Scholar] [CrossRef]
- McClain, C.J.; Kromhout, J.P.; Zieve, L.; Duane, W.C. Effect of sorbitol on psychomotor function: Its use in alcoholic cirrhosis. Arch. Intern. Med. 1981, 141, 901–903. [Google Scholar] [CrossRef]
- Peters, R.; Lock, R. Laxative effect of sorbitol. Br. Med. J. 1958, 2, 677. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Direction | Primer Sequence | Accession Number |
|---|---|---|---|
| Bax | Sense | GGCGAATTGGCGATGAACTG | NM_017059.2 |
| Antisense | ATGGTTCTGATCAGCTCGGG | ||
| Bcl-2 | Sense | GATTGTGGCCTTCTTTGAGT | NM_016993.1 |
| Antisense | ATAGTTCCACAAAGGCATCC | ||
| GAPDH | Sense | TCAAGAAGGTGGTGAAGCAG | NM_017008.4 |
| Antisense | AGGTGGAAGAATGGGAGTTG | ||
| IL-1β | Sense | ACC CAA GCA CCT TCT TTT CCT T | NM_031512.2 |
| Antisense | ACG GGA AAC CCA TCA CCA T | ||
| HMOX1 | Sense | AGCATGTCCCAGGATTTGTC | NM_012580.2 |
| Antisense | TCACCAGCTTAAAGCCTTCC | ||
| NRF2 | Sense | TTGTAGATGACCATGAGTCGC | NM_031789 |
| Antisense | TGTCCTGCTGTATGCTGCTT | ||
| IFN-γ | Sense | AGGTGAACAACCCACAGAT | NM_138880.3 |
| Antisense | CTTCTTATTGGCACACTCTCTAC |
| Control | Corn Oil Group (Sham) | Malathion | Malathion + Ornipural® | Ornipural® | |
|---|---|---|---|---|---|
| Initial body Weight (g) | 135.05 ± 5.14 | 137.5 ± 5.3 | 144.45 ± 5.4 | 139.48 ± 6.48 | 141.45 ± 4.15 |
| Final body Weight (g) | 268.15 ± 6.45 | 267.9 ± 6.5 | 231.01 ± 6.9 ## | 248.15 ± 8.45 *+ | 265.1 ± 5.3 |
| Bodyweight gain | 133.15 ± 4.8 | 130.4 ± 7.45 | 86.56 ± 4.5 ## | 108.67 ± 4.15 *+ | 123.65 ± 4.15 |
| Absolute Weight of Liver (g) | 6.45 ± 0.52 | 6.63 ± 0.15 | 5.321 ± 0.22 | 5.48 ± 0.6 | 6.51 ± 0.2 |
| Relative liver Weight (g/100 g BW) | 2.40 ± 0.14 | 2.47 ± 0.15 | 2.30 ± 0.14 | 2.20 ± 0.14 | 2.45 ± 0.01 |
| Absolute Weight of Kidney (g) | 1.79 ± 0.12 | 1.77 ± 0.14 | 1.69 ± 0.014 | 1.75 ± 0.1 | 1.76 ± 0.05 |
| Relative Weight of Kidney (g/100 g BW) | 0.66 ± 0.01 | 0.660 ± 0.014 | 0.733 ± 0.01 | 0.70 ± 0.01 | 0.66 ± 0.1 |
| Control | Corn Oil Group | Malathion | Malathion + Ornipural® | Ornipural® | |
|---|---|---|---|---|---|
| AST (U/mL) | 80.58 ± 5.2 | 79.00 ± 5.9 | 172.86 ± 8.6 ## | 89.82 ± 4.5 *+ | 75.26 ± 2.3 |
| ALT (U/mL) | 35.15 ± 3.1 | 34.46 ± 2.5 | 77.07 ± 4.2 ## | 48.43 ± 3.2 *+ | 36.29 ± 1.5 |
| ALP (U/L) | 85.20 ± 4.45 | 83.53 ± 3.6 | 202.49 ± 15.14 ## | 99.82 ± 5.3 **+ | 77.88 ± 4.6 & |
| LDH (U/L) | 194.66 ± 10.2 | 195.75 ± 12.3 | 431.95 ± 17.2 ## | 298.03 ± 10.2 **+ | 192.87 ± 9.01 |
| ACP(U/L) | 101.12 ± 11.2 | 102.14 ± 10.2 | 181.3 ± 10.14 ## | 119.34 ± 10.45 *+ | 100.14 ± 2.9 |
| Bilirubin (mg/dL) | 5.22 ± 0.14 | 5.10 ± 0.10 | 7.15 ± 0.6 # | 6.12 ± 0.4 *+ | 5.01 ± 0.4 |
| Total protein (g/L) | 5.10 ± 0.5 | 5.00 ± 0.1 | 3.42 ± 0.4 # | 4.20 ± 0.2 *+ | 4.95 ± 0.5 |
| Albumin (g/L) | 4.02 ± 0.4 | 3.94 ± 0.6 | 2.92 ± 0.1 # | 3.47 ± 0.1 *+ | 3.91 ± 0.1 |
| Triglycerides (g/L) | 110.29 ± 6.3 | 108.12 ± 10.4 | 67.78 ± 1.3 ## | 84.23 ± 4.5 *+ | 105.05 ± 3.8 |
| HDL-C (mg/dL) | 65.84 ± 4.45 | 63.01 ± 6.48 | 49.12 ± 3.8 ## | 53.14 ± 4.7 *+ | 66.8 ± 5.6 |
| LDL-C (mg/dL) | 101.14 ± 6.1 | 100.4 ± 6.2 | 125.14 ± 6.4 ## | 114.12 ± 4.5 *+ | 99.15 ± 5.2 |
| Cholesterol (mg/dL) | 77.44 ± 4.6 | 79.88 ± 3.4 | 149.10 ± 9.5 ## | 104.20 ± 5.6 *+ | 71.19 ± 3.5 & |
| Uric acid (mg/dL) | 25.46 ± 3.6 | 24.96 ± 1.01 | 80.05 ± 7.5 ## | 41.05 ± 2.5 *+ | 20.97 ± 1.01 |
| Urea (mg/dL) | 21.83 ± 2.3 | 21.40 ± 1.0 | 73.99 ± 4.5 ## | 40.26 ± 3.2 *+ | 18.35 ± 1.04 |
| Creatinine (mg %) | 0.66 ± 0.01 | 0.64 ± 0.01 | 2.05 ± 0.3 # | 1.76 ± 0.1 *+ | 0.57 ± 0.1 |
| AChE (U/L) | 250.01 ± 10.15 | 245.15 ± 11.2 | 75.96 ± 5.8 ## | 126.15 ± 6.48 *+ | 233.15 ± 12.4 |
| Paraoxonase (U/L) | 176.14 ± 13.1 | 177.5 ± 12.4 | 120.14 ± 5.4 ## | 138.56 ± 7.14 *+ | 181.14 ± 13.1 |
| Ammonia (μmol/L) | 128.15 ± 10.12 | 131.14 ± 9.48 | 256.1 ± 17.69 ## | 186.14 ± 12.14 **+ | 130.14 ± 12.69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Okle, O.S.; Tohamy, H.G.; Althobaiti, S.A.; Soliman, M.M.; Ghamry, H.I.; Farrag, F.; Shukry, M. Ornipural® Mitigates Malathion-Induced Hepato-Renal Damage in Rats via Amelioration of Oxidative Stress Biomarkers, Restoration of Antioxidant Activity, and Attenuation of Inflammatory Response. Antioxidants 2022, 11, 757. https://doi.org/10.3390/antiox11040757
El Okle OS, Tohamy HG, Althobaiti SA, Soliman MM, Ghamry HI, Farrag F, Shukry M. Ornipural® Mitigates Malathion-Induced Hepato-Renal Damage in Rats via Amelioration of Oxidative Stress Biomarkers, Restoration of Antioxidant Activity, and Attenuation of Inflammatory Response. Antioxidants. 2022; 11(4):757. https://doi.org/10.3390/antiox11040757
Chicago/Turabian StyleEl Okle, Osama S., Hossam G. Tohamy, Saed A. Althobaiti, Mohamed Mohamed Soliman, Heba I. Ghamry, Foad Farrag, and Mustafa Shukry. 2022. "Ornipural® Mitigates Malathion-Induced Hepato-Renal Damage in Rats via Amelioration of Oxidative Stress Biomarkers, Restoration of Antioxidant Activity, and Attenuation of Inflammatory Response" Antioxidants 11, no. 4: 757. https://doi.org/10.3390/antiox11040757
APA StyleEl Okle, O. S., Tohamy, H. G., Althobaiti, S. A., Soliman, M. M., Ghamry, H. I., Farrag, F., & Shukry, M. (2022). Ornipural® Mitigates Malathion-Induced Hepato-Renal Damage in Rats via Amelioration of Oxidative Stress Biomarkers, Restoration of Antioxidant Activity, and Attenuation of Inflammatory Response. Antioxidants, 11(4), 757. https://doi.org/10.3390/antiox11040757