
β-Carotene Increases Activity of Cytochrome P450 2E1 during Ethanol Consumption

 ,
, 


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Euthanasia
2.3. Liver Tissue
2.4. Extraction of RNA and cDNA Synthesis
2.5. Quantification of RNA from Liver Tissue
2.6. Quantitative Real-Time PCR
2.7. Statistical Analysis
3. Results
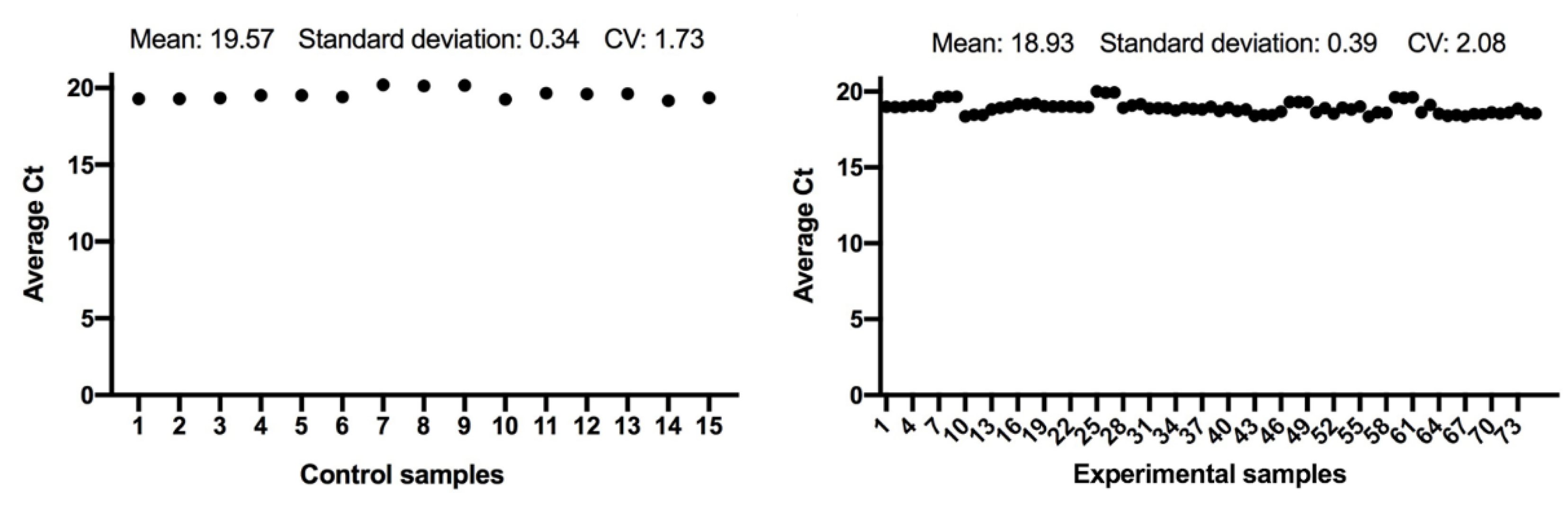
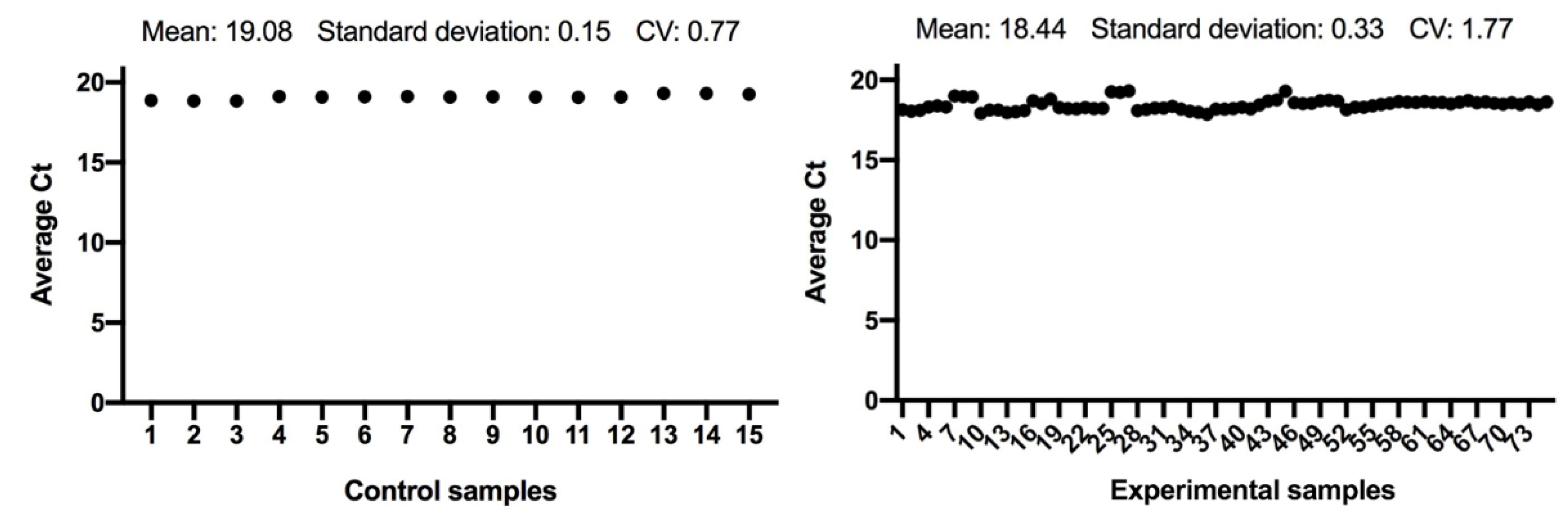
3.1. Statistical Analysis
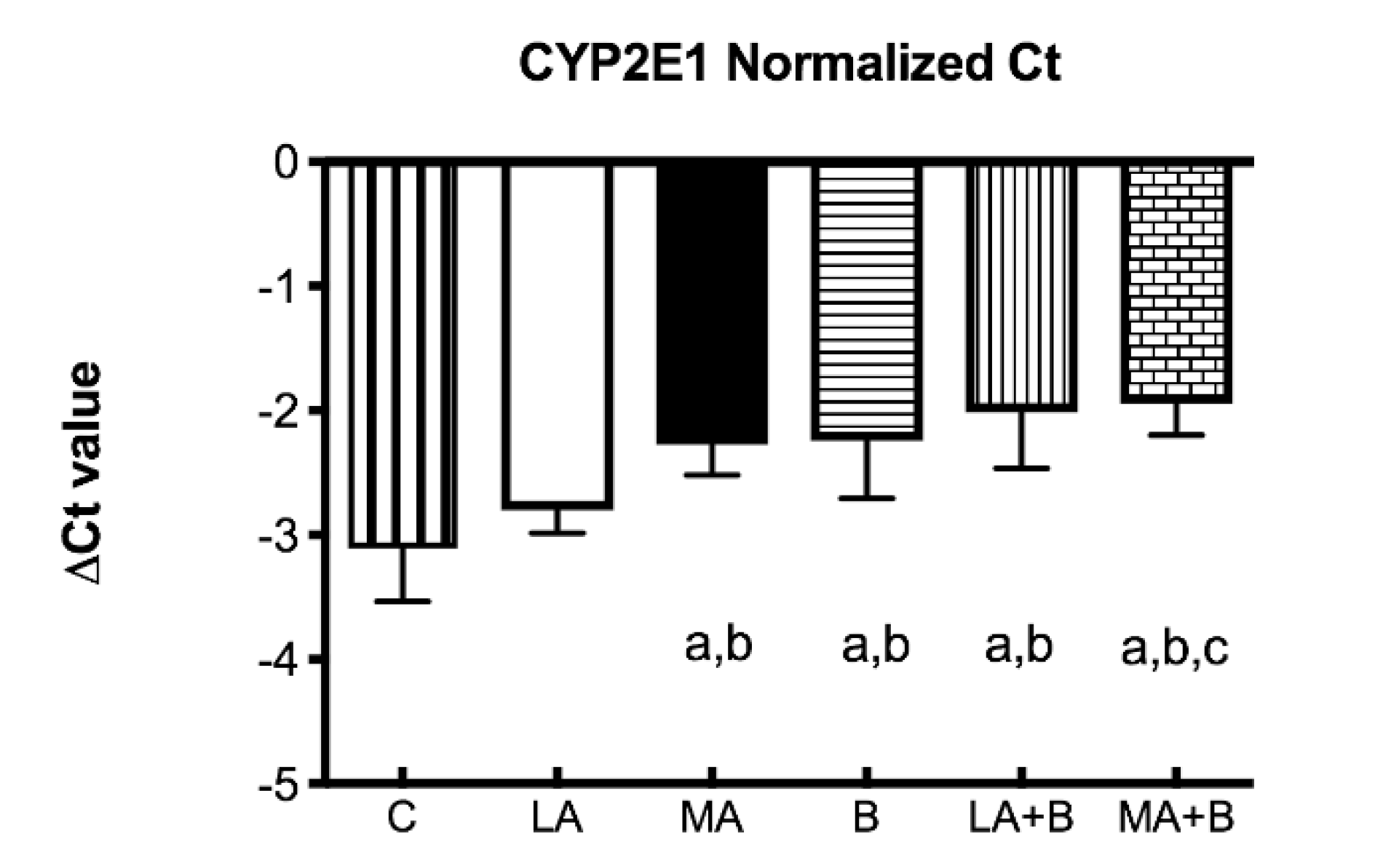
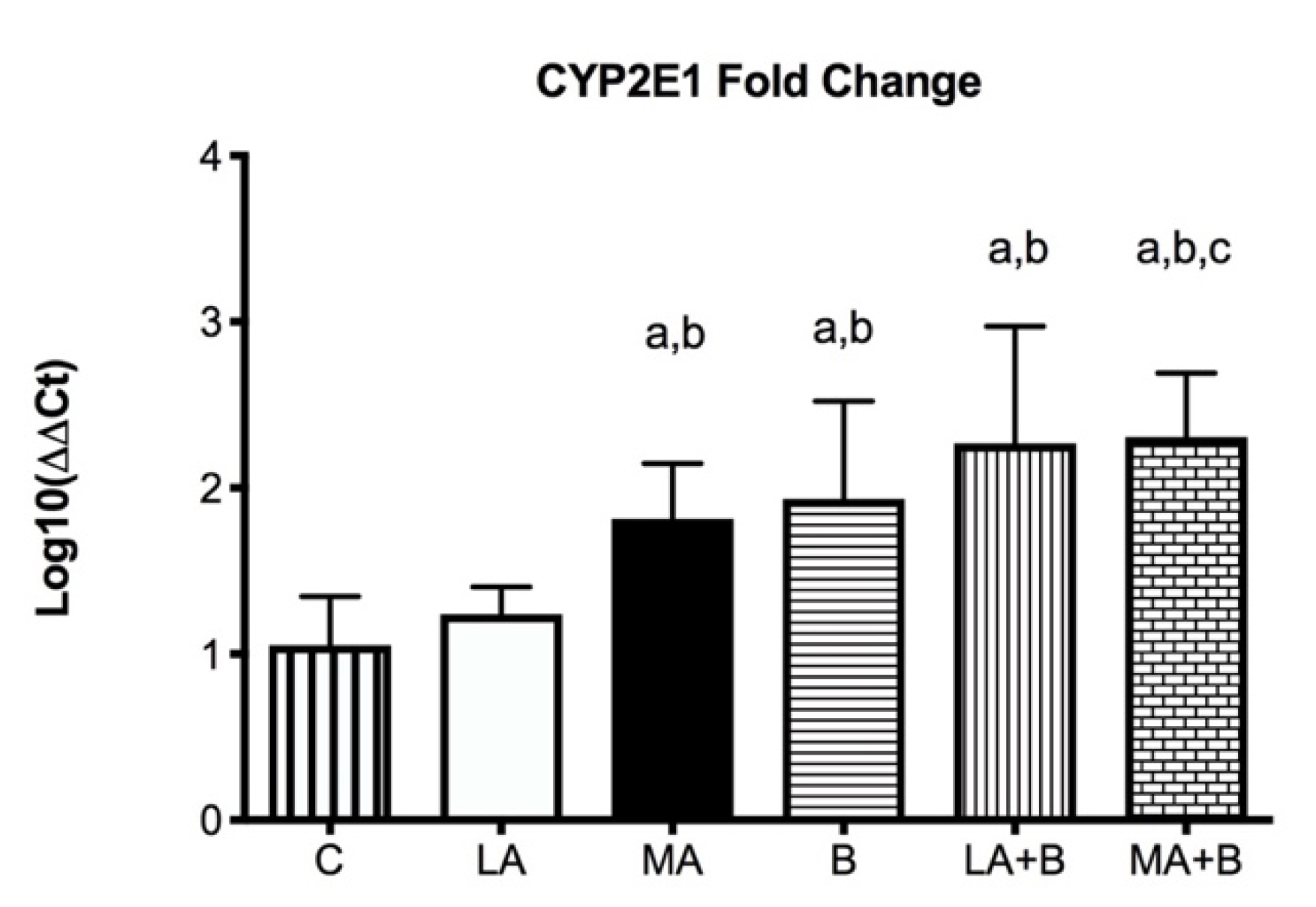
3.2. Quantitative Real-Time PCR
Control group: ΔCt = Ct (target) − Ct (housekeeping genes)
ΔΔCt = ΔCt (experimental groups) − ΔCt (control group)
Ratio = 2−ΔΔCt
4. Discussion
4.1. Summary of Key Findings and Interpretation
4.2. Scope and Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Haber, P.S.; Apte, M.V.; Moran, C.; Applegate, T.L.; Pirola, R.C.; Korsten, M.A.; McCaughan, G.W.; Wilson, J.S. Non-oxidative Metabolism of Ethanol by Rat Pancreatic Acini. Pancreatology 2004, 4, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Brust, J.C.M. Ethanol and Cognition: Indirect Effects, Neurotoxicity, and Neuroprotection: A Review. Int. J. Environ. Res. Public Health 2010, 7, 1540–1557. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Alcohol. Descriptive Note Nº 349; World Health Organization: New York, NY, USA, 2011.
- Cederbaum, A.I. Cytochrome P450 2E1-dependent Oxidant Stress and Upregulation of Anti-oxidant Defense in Liver Cells. J. Gastroenterol. Hepatol. 2006, 21, S22–S25. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.I.; Roychowdhury, S.; DiBello, P.M.; Jacobsen, D.W.; Nagy, L.E. Exogenous Thioredoxin Prevents Ethanol-induced Oxidative Damage and Apoptosis in Mouse Liver. Hepatology 2009, 49, 1709–1717. [Google Scholar] [CrossRef] [Green Version]
- García Gutiérrez, E.; Lima Mompó, G.; Aldana Vilas, L.; Casanova Carrillo, P.; Feliciano Álvarez, V. Alcoholismo y sociedad, tendencias actuales. Rev. Cub. Med. Mil. 2004, 33, 3. [Google Scholar]
- Arias, R. Reacciones fisiológicas y neuroquímicas del alcoholismo. Diversitas 2005, 1, 138–147. [Google Scholar] [CrossRef]
- Lakshman, R.; Cederbaum, A.I.; Hoek, J.B.; Konishi, M.; Koop, D.; Donohue, T.M. Use of CYP2E1-transfected Human Liver Cell Lines in Elucidating the Actions of Ethanol. Alcohol Clin. Exp. Res. 2006, 29, 1726–1734. [Google Scholar] [CrossRef]
- Wu, H.; Cai, P.; Clemens, D.L.; Jerrells, T.R.; Shakeel Ansari, G.A.; Kaphalia, B.S. Metabolic Basis of Ethanol-induced Cytotoxicity in Recombinant HepG2 Cells: Role of Nonoxidative Metabolism. Toxicol. Appl. Pharmacol. 2006, 216, 238–247. [Google Scholar] [CrossRef]
- Lu, Y.; Cederbaum, A.I. CYP2E1 and Oxidative Liver Injury by Alcohol. Free Rad. Biol. Med. 2008, 44, 723–738. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Klionsky, D.J. Eaten Alive: A History of Macroautophagy. Nat. Cell Biol. 2010, 12, 814–822. [Google Scholar] [CrossRef] [Green Version]
- Bailey, S.M.; Cunningham, C.C. Contribution of Mitochondria to Oxidative Stress Associated with Alcoholic Liver Disease. Free Rad. Biol. Med. 2002, 32, 11–16. [Google Scholar] [CrossRef]
- Hoek, J.B.; Cahill, A.; Pastorino, J.G. Alcohol and Mitochondria: A Dysfunctional Relationship. Gastroenterology 2002, 122, 2049–2063. [Google Scholar] [CrossRef] [Green Version]
- Tuma, D.J.; Casey, C.A. Dangerous Byproducts of Alcohol Breakdown: Focus on Adducts. Alcohol Res. Health 2003, 27, 285–290. [Google Scholar]
- Sandoval, C.; Vaásquez, B.; Mandarim-de-Lacerda, C.; del Sol, M. Ethanol Intake and Toxicity: In Search of New Treatments. Int. J. Morphol. 2017, 35, 942–949. [Google Scholar] [CrossRef] [Green Version]
- Schattenberg, J.M.; Czaja, M.J. Regulation of the Effects of CYP2E1-induced Oxidative Stress by JNK Signaling. Redox Biol. 2014, 3, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Diesinger, T.; Buko, V.; Lautwein, A.; Dvorsky, R.; Belonovskaya, E.; Lukivskaya, O.; Naruta, E.; Kirko, S.; Andreev, V.; Buckert, D.; et al. Drug Targeting CYP2E1 for the Treatment of Early-stage Alcoholic Steatohepatitis. PLoS ONE 2020, 15, e0235990. [Google Scholar] [CrossRef]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide Dismutase Multigene Family: A Comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) Gene Structures, Evolution, and Expression. Free Rad. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Chang, P.; Cheng, E.; Brooke, S.; Sapolsky, R. Marked Differences in the Efficacy of Post-insult Gene Therapy with Catalase versus Glutathione Peroxidase. Brain Res. 2005, 1063, 27–31. [Google Scholar] [CrossRef]
- Sandoval, C.; Vásquez, B.; Souza-Mello, V.; Adeli, K.; Mandarim-de-Lacerda, C.; del Sol, M. Morphoquantitative Effects of Oral β-carotene Supplementation on Liver of C57BL/6 Mice Exposed to Ethanol Consumption. Int. J. Clin. Exp. Pathol. 2019, 12, 1713–1722. [Google Scholar]
- Schott, M.B.; Rasineni, K.; Weller, S.G.; Schulze, R.J.; Sletten, A.C.; Casey, C.A.; McNiven, M.A. β-Adrenergic Induction of Lipolysis in Hepatocytes Is Inhibited by Ethanol Exposure. J. Biol. Chem. 2017, 292, 11815–11828. [Google Scholar] [CrossRef] [Green Version]
- Werling, K. A Májbetegségek Kialakulásának Új Szempontjai—Különös Tekintettel Az Autophagiára Ès A mikro-RNS Szerepére. Orvosi Hetilap 2020, 161, 1449–1455. [Google Scholar] [CrossRef]
- Committee for the Update of the Guide for the Care and Use of Laboratory Animals; Institute for Laboratory Animal Research; Division on Earth and Life Studies. Guide for the Care and Use of Laboratory Animals, 8th ed.; The National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Furuya, D.T.; Binsack, R.; Machado, U.F. Low ethanol consumption increases insulin sensitivity in Wistar rats. Braz. J. Med. Biol. Res. 2003, 36, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.C.; Chen, Y.L.; Yang, S.Y.; Ho, P.Y.; Yang, S.S.; Hu, J.T.; Yang, S.C. The antiapoptotic effects of different doses of β-carotene in chronic ethanol-fed rats. Hepatobiliary Surg. Nutr. 2013, 2, 132–141. [Google Scholar] [CrossRef]
- Diao, Y.; Nie, J.; Tan, P.; Zhao, Y.; Zhao, T.; Tu, J.; Ji, H.; Cao, Y.; Wu, Z.; Liang, H.; et al. Long-term low-dose ethanol intake improves healthspan and resists high-fat diet-induced obesity in mice. Aging 2020, 12, 13128–13146. [Google Scholar] [CrossRef]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA Using TRIzol (TRI Reagent). Cold Spring Harb. Protoc. 2010, 2010, pdb.prot5439. [Google Scholar] [CrossRef]
- Sheng, Q.; Vickers, K.; Zhao, S.; Wang, J.; Samuels, D.C.; Koues, O.; Shyr, Y.; Guo, Y. Multi-perspective quality control of Illumina RNA sequencing data analysis. Brief. Funct. Genom. 2017, 16, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Bustin, S.A. Quantification of mRNA using real-time reverse transcription PCR (RT-PCR): Trends and problems. J. Mol. Endocrinol. 2002, 29, 23–29. [Google Scholar] [CrossRef]
- Pfaffl, M.W. Quantification Strategies in Real-Time PCR. In A-Z of Quantitative PCR; International University Line: La Jolla, CA, USA, 2004. [Google Scholar]
- Finis, K.; Sültmann, H.; Ruschhaupt, M.; Buness, A.; Helmchen, B.; Ruprecht, K.; Gross, M.L.; Fink, B.; Schirmacher, P.; Poustka, A. Analysis of pigmented villonodular synovitis with genome-wide complementary DNA microarray and tissue array technology reveals insight into potential novel therapeutic approaches. Arthritis Rheum. 2006, 54, 1009–1019. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantification PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Arocho, A.; Chen, B.; Ladanyi, M.; Pan, Q. Validation of the 2-DeltaDeltaCt calculation as an alternate method of data analysis for quantitative PCR of BCR-ABL P210 transcripts. Diagn. Mol. Pathol. 2006, 15, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Silvestri, L.; Sonzogni, L.; De Silvestri, A. CYP Enzyme Polymorphısms and Susceptıbılıty to HCV-Related Chronıc Liver Disease and Liver Cancer. Int. J. Cancer 2003, 104, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Lieber, C.S. The Discovery of the Microsomal Ethanol Oxidizing System and Its Physiologic and Pathologic Role. Drug Metab. Rev. 2004, 36, 511–529. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, C.; Vásquez, B.; Souza-Mello, V.; Mandarim-de-Lacerda, C.A.; del Sol, M. Rol del consumo de alcohol y antioxidantes sobre la metilación global del ADN y cáncer. Int. J. Morphol. 2018, 36, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Carrasco, C.; Carrasco, C.; Souza-Mello, V.; Sandoval, C. Effectiveness of antioxidant treatments on cytochrome P450 2E1 (CYP2E1) activity after alcohol exposure in humans and in vitro models: A systematic review. Int. J. Food Prop. 2021, 24, 1300–1317. [Google Scholar] [CrossRef]
- Sandoval, C.; Vásquez, B.; Vasconcellos, A.; Souza-Mello, V.; Adeli, K.; Mandarim-de-Lacerda, C.; del Sol, M. Oral supplementation of b-carotene benefits the hepatic structure and metabolism in mice exposed to chronic ethanol consumption. Sains Malays. 2022, 51, 285–296. [Google Scholar] [CrossRef]
- Niemelä, O.; Parkkila, S.; Pasanen, M.; Viitala, K.; Villanueva, J.A.; Halsted, C.H. Induction of cytochrome P450 enzymes and generation of protein-aldehyde adducts are associated with sex-dependent sensitivity to alcohol-induced liver disease in micropigs. Hepatology 1999, 30, 1011–1017. [Google Scholar] [CrossRef]
- Liangpunsakul, S.; Kolwankar, D.; Pinto, A.; Gotski, J.C.; Hall, S.D.; Chalasani, N. Activity of CYP2E1 and CYP3A enzymes in adults with moderate alcohol consumption: A comparison with nonalcoholics. Hepatology 2005, 41, 1144–1150. [Google Scholar] [CrossRef]
- Lu, Y.; Cederbaum, A.I. Cisplatin-Induced Hepatotoxicity Is Enhanced by Elevated Expression of Cytochrome P450 2E1. Toxıcol. Sci. 2006, 89, 515–523. [Google Scholar] [CrossRef]
- Lin, W.T.; Huang, C.C.; Lin, T.J.; Chen, J.R.; Shieh, M.J.; Peng, H.C.; Yang, S.C.; Huang, C.Y. Effects of beta-carotene on antioxidant status in rats with chronic alcohol consumption. Cell Biochem. Funct. 2009, 27, 344–350. [Google Scholar] [CrossRef]
- Sandoval, C.; Farías, J.; Zamorano, M.; Herrera, C. Vitamin Supplements as a Nutritional Strategy against Chronic Alcohol Consumption? An Updated Review. Antioxidants 2022, 11, 564. [Google Scholar] [CrossRef]
- Ronis, M.J.; Huang, J.; Crouch, J.; Mercado, C.; Irby, D.; Valentine, C.R.; Lumpkin, C.K.; Ingelman-Sundberg, M.; Badger, T.M. Cytochrome P450 CYP 2E1 induction during chronic alcohol exposure occurs by a two-step mechanism associated with blood alcohol concentrations in rats. J. Pharmacol. Exp. Ther. 1993, 264, 944–950. [Google Scholar]
- Eliasson, E.; Johansson, I.; Ingelman-Sundberg, M. Ligand-dependent maintenance of ethanol-inducible cytochrome P-450 in primary rat hepatocyte cell cultures. Biochem. Biophys. Res. Commun. 1988, 150, 436–443. [Google Scholar] [CrossRef]
- Lieber, C.S. Microsomal ethanol-oxidizing system (MEOS): The first 30 years (1968–1998)—A review. Alcohol. Clin. Exp. Res. 1999, 23, 991–1007. [Google Scholar] [CrossRef]
- Ziegler, R.G. A review of epidemiologic evidence that carotenoids reduce the risk of cancer. J. Nutr. 1989, 119, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, R.G. Vegetables, fruits, and carotenoids and the risk of cancer. Am. J. Clin. Nutr. 1991, 53 (Suppl. S1), 251S–259S. [Google Scholar] [CrossRef]
- Stahelin, H.B.; Gey, K.F.; Eichholzer, M.; Ludin, E. B-carotene and cancer prevention: The Base1 Study. Am. J. Clin. Nutr. 1991, 53 (Suppl. S1), 265S–269S. [Google Scholar] [CrossRef]
- Paganini-Hill, A.; Chao, A.; Ross, R.K.; Henderson, B.E. Vitamin A, @-carotene, and the risk of cancer: A prospective study. J. Natl. Cancer Inst. 1987, 79, 443–448. [Google Scholar]
- Leo, M.A.; Kim, C.; Lowe, N.; Lieber, C.S. Interaction of ethanol with beta-carotene: Delayed blood clearance and enhanced hepatotoxicity. Hepatology 1992, 15, 883–891. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Symbol | Assay | Chromosome Location | Amplicon Length |
|---|---|---|---|---|
| cytochrome P450, family 2, subfamily e, polypeptide 1 | Cyp2e1 | Mm00491127_m1 | Chr.7: 140763832–140774981 | 83 bp |
| Gene | Gene Symbol | Assay | Chromosome Location | Amplicon-Length |
|---|---|---|---|---|
| actx, E430023M04Rik, beta-actin | Actb | Mm00607939_s1 | Chr.5: 142903116–142906724 | 115 bp |
| glyceraldehyde-3-phosphate dehydrogenase | Gapdh | Mm99999915_g1 | Chr.6: 125161338–125166511 | 107 bp |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandoval, C.; Mella, L.; Godoy, K.; Adeli, K.; Farías, J. β-Carotene Increases Activity of Cytochrome P450 2E1 during Ethanol Consumption. Antioxidants 2022, 11, 1033. https://doi.org/10.3390/antiox11051033
Sandoval C, Mella L, Godoy K, Adeli K, Farías J. β-Carotene Increases Activity of Cytochrome P450 2E1 during Ethanol Consumption. Antioxidants. 2022; 11(5):1033. https://doi.org/10.3390/antiox11051033
Chicago/Turabian StyleSandoval, Cristian, Luciana Mella, Karina Godoy, Khosrow Adeli, and Jorge Farías. 2022. "β-Carotene Increases Activity of Cytochrome P450 2E1 during Ethanol Consumption" Antioxidants 11, no. 5: 1033. https://doi.org/10.3390/antiox11051033
APA StyleSandoval, C., Mella, L., Godoy, K., Adeli, K., & Farías, J. (2022). β-Carotene Increases Activity of Cytochrome P450 2E1 during Ethanol Consumption. Antioxidants, 11(5), 1033. https://doi.org/10.3390/antiox11051033