
Dietary Tryptophan Supplementation Improves Antioxidant Status and Alleviates Inflammation, Endoplasmic Reticulum Stress, Apoptosis, and Pyroptosis in the Intestine of Piglets after Lipopolysaccharide Challenge

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pig Care and Experimental Design
2.2. Intestinal Sample Collections
2.3. Intestinal Morphology Analysis
2.4. Measurement of the Antioxidant Parameters
2.5. Real-time PCR Analysis
2.6. Western Blot Assay
2.7. Statistical Analysis
3. Results
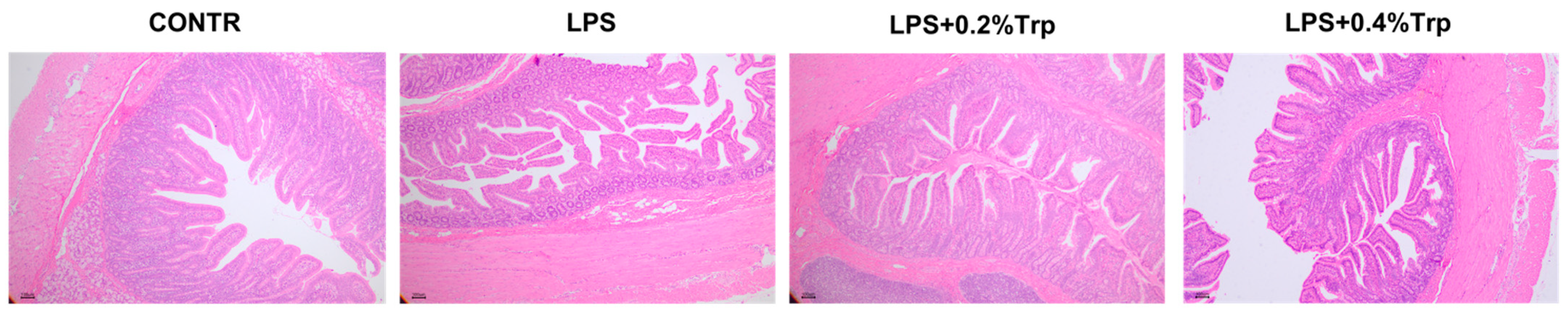
3.1. Intestinal Morphology
3.2. Antioxidant Indicators in the Ileum
3.3. The Concentration and mRNA Expression of Ileal Cytokines and Activity of IDO
3.4. mRNA Expression Levels of ER Stress-, Pyroptosis, and Apoptosis-Related Genes in Ileal Tissues
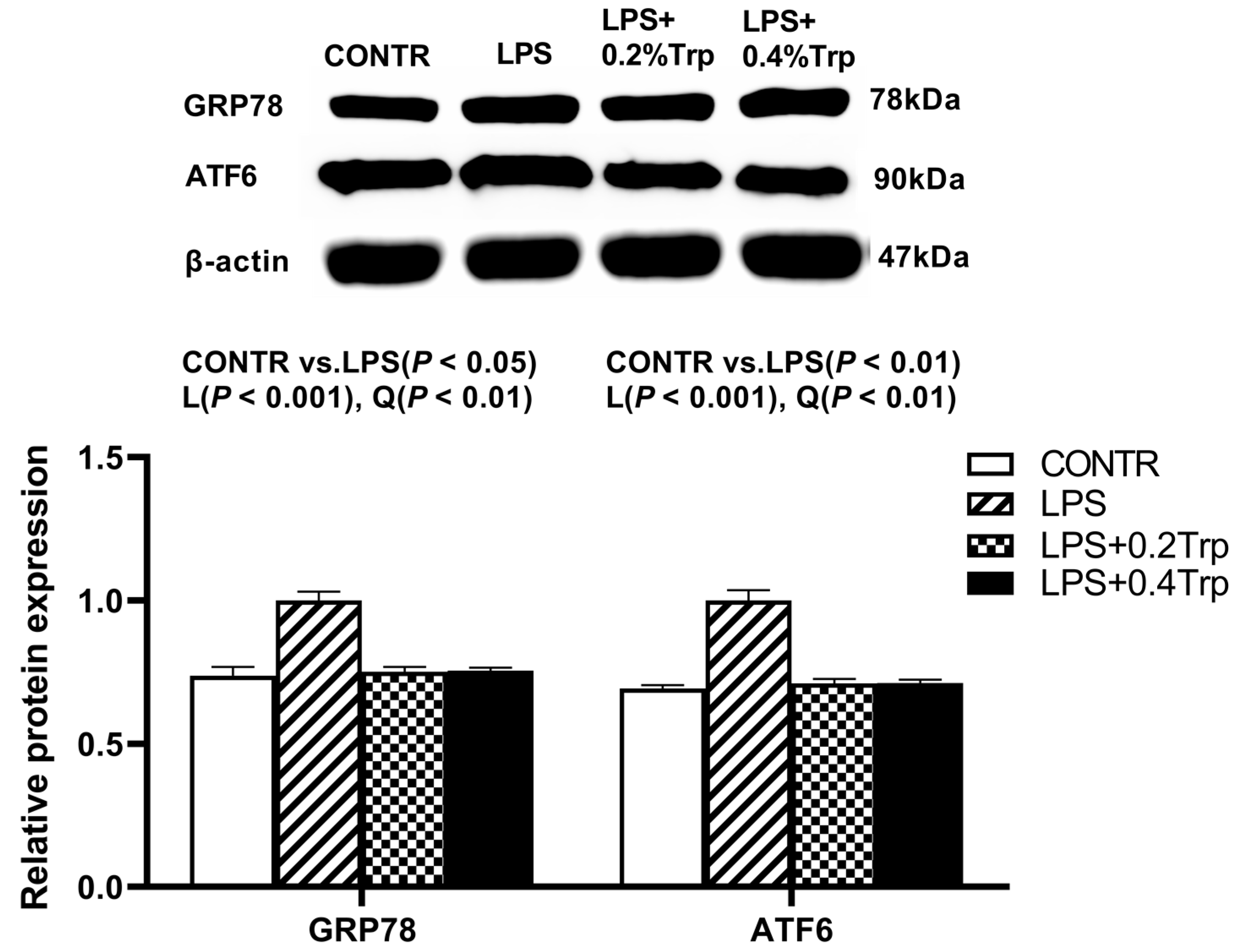
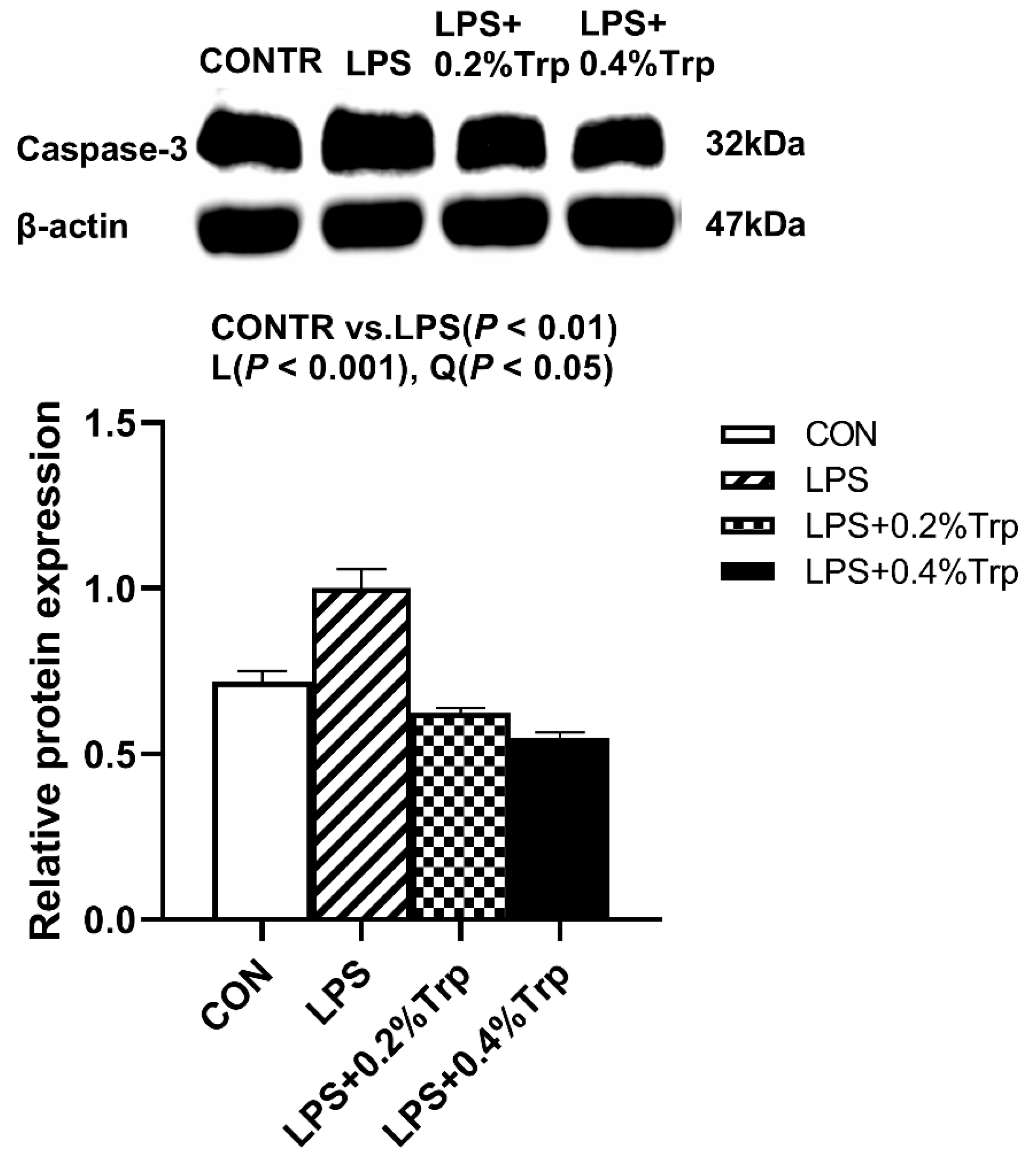
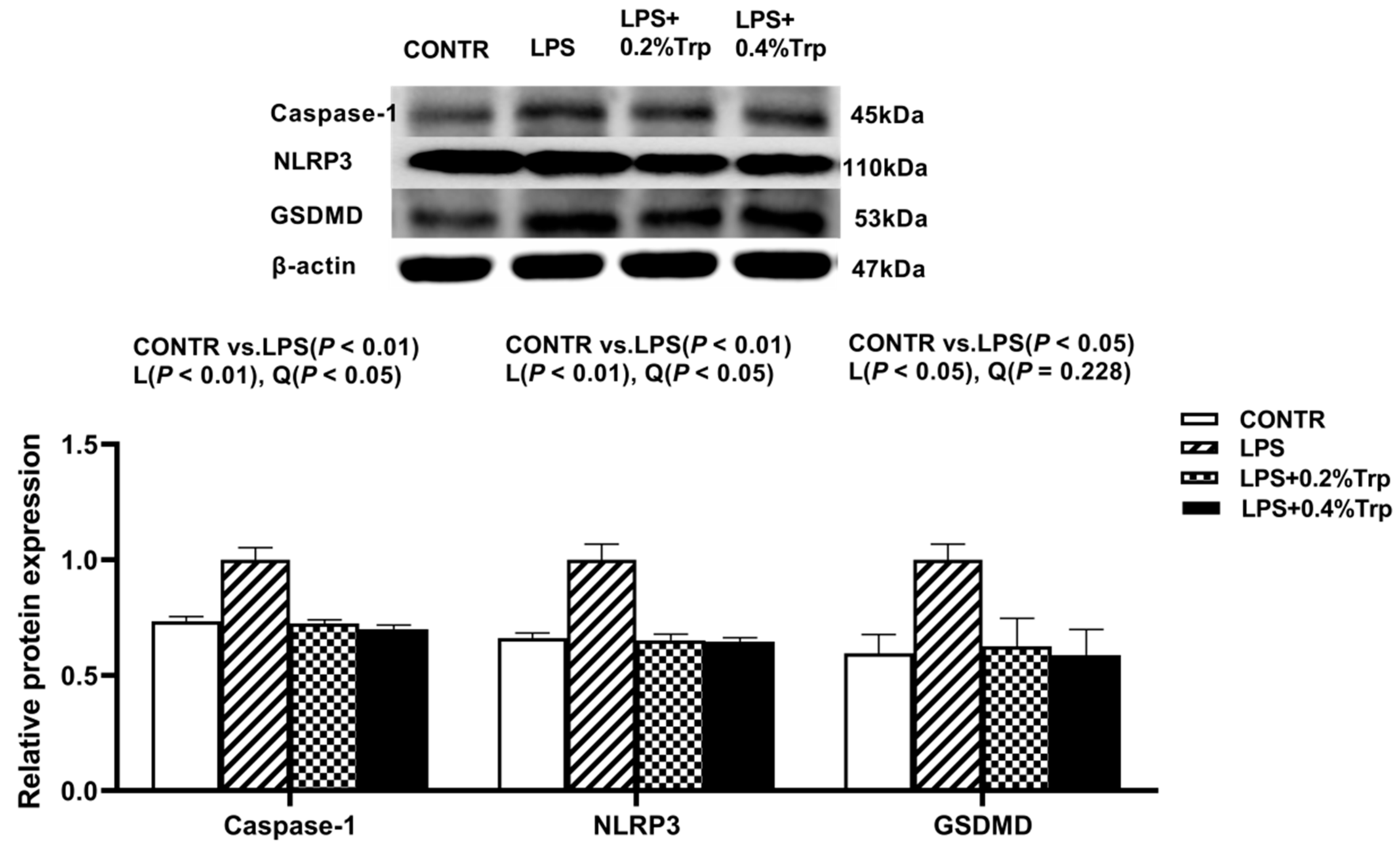
3.5. ER Stress-, Pyroptosis-, and Apoptosis-Related Protein Expression Levels in Ileal Tissues of Piglets
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zechner, E.L. Inflammatory disease caused by intestinal pathobionts. Curr. Opin. Microbiol. 2017, 35, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhong, S.; Wang, F.; Wang, H.; Xu, D.; Li, G. Microcystin-LR exposure decreased the fetal weight of mice by disturbance of placental development and ROS-mediated endoplasmic reticulum stress in the placenta. Environ. Pollut. 2020, 256, 113362. [Google Scholar] [CrossRef]
- Tian, W.; Wang, Z.; Tang, N.N.; Li, J.T.; Liu, Y.; Chu, W.F.; Yang, B.F. Ascorbic acid sensitizes colorectal carcinoma to the cytotoxicity of arsenic trioxide via promoting reactive oxygen species-dependent apoptosis and pyroptosis. Front. Pharmacol. 2020, 11, 123. [Google Scholar] [CrossRef] [PubMed]
- Pié, S.; Lallès, J.P.; Blazy, F.; Laffitte, J.; Sève, B.; Oswald, I.P. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.; Xiong, M.; Xu, X.; Wu, X.; Xu, J.; Cai, X.; Lu, L.; Zhou, H. Bile acids elevated by high-fat feeding induce endoplasmic reticulum stress in intestinal stem cells and contribute to mucosal barrier damage. Biochem. Biophys. Res. Commun. 2020, 529, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wu, G.; Zhou, Z.; Dai, Z.; Sun, Y.; Ji, Y.; Li, W.; Wang, W.; Liu, C.; Han, F.; et al. Glutamine and intestinal barrier function. Amino Acids 2015, 47, 2143–2154. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Gu, K.; Wang, F.; Jia, G.; Zhao, H.; Chen, X.; Wu, C.; Zhang, R.; Tian, G.; Cai, J.; et al. Tryptophan ameliorates barrier integrity and alleviates the inflammatory response to enterotoxigenic Escherichia coli K88 through the CaSR/Rac1/PLC-γ1 signaling pathway in porcine intestinal epithelial cells. Front. Immunol. 2021, 12, 748497. [Google Scholar] [CrossRef]
- Naatjes, M.; Htoo, J.K.; Walter, K.; Tölle, K.H.; Susenbeth, A. Effect of dietary tryptophan to lysine ratio on growth of young pigs fed wheat–barley or corn based diets. Livest. Sci. 2014, 163, 102–109. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Y.; Li, Y.; Yan, H.; Zhang, H. L-tryptophan enhances intestinal integrity in diquat-challenged piglets associated with improvement of redox status and mitochondrial function. Animals 2019, 9, 266. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, S.; Togashi, S.; Takahashi, N.; Fukui, T. L-tryptophan as an antioxidant in human placenta extract. J. Nutr. Sci. Vitaminol. 2002, 48, 36–39. [Google Scholar] [CrossRef]
- Zelante, T.; Iannitti, R.G.; Cunha, C.; De Luca, A.; Giovannini, G.; Pieraccini, G.; Zecchi, R.; D’Angelo, C.; Massi-Benedetti, C.; Fallarino, F.; et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 2013, 39, 372–385. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Dai, Z.; Liu, N.; Ji, Y.; Chen, J.; Zhang, Y.; Yang, Y.; Li, J.; Wu, Z.; Wu, G. Dietary L-tryptophan modulates the structural and functional composition of the intestinal microbiome in weaned piglets. Front. Microbiol. 2018, 9, 1736. [Google Scholar] [CrossRef]
- Tang, Z.; Shi, B.; Sun, W.; Yin, Y.; Chen, Q.; Mohamed, T.; Chang, W.; Sun, Z. Tryptophan promoted β-defensin-2 expression via the mtor pathway and its metabolites: Kynurenine banding to aryl hydrocarbon receptor in rat intestine. RSC Adv. 2020, 10, 3371–3379. [Google Scholar] [CrossRef] [Green Version]
- Gu, K.; Liu, G.; Wu, C.; Jia, G.; Zhao, H.; Chen, X.; Tian, G.; Cai, J.; Zhang, R.; Wang, J. Tryptophan improves porcine intestinal epithelial cell restitution through the CaSR/Rac1/PLC-γ1 signaling pathway. Food Funct. 2021, 12, 8787–8799. [Google Scholar] [CrossRef]
- Perianayagam, M.C.; Oxenkrug, G.F.; Jaber, B.L. Immune-modulating effects of melatonin, N-acetylserotonin, and N-acetyldopamine. Ann. N. Y. Acad. Sci. 2005, 1053, 386–393. [Google Scholar] [CrossRef]
- Zhang, H.M.; Zhang, Y. Melatonin: A well-documented antioxidant with conditional pro-oxidant actions. J. Pineal Res. 2014, 57, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.K.; Naidoo, N. The UPR and the anti-oxidant response: Relevance to sleep and sleep loss. Mol. Neurobiol. 2010, 42, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, H.; Wang, S.; Tu, Z.; Zhang, L.; Wang, X.; Hou, Y.; Wang, C.; Chen, J.; Liu, Y. Flaxseed oil attenuates intestinal damage and inflammation by regulating necroptosis and TLR4/NOD signaling pathways following lipopolysaccharide challenge in a piglet model. Mol. Nutr. Food Res. 2018, 62, e1700814. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Cao, W.; Liu, G.; Fang, T.; Wu, X.; Jia, G.; Chen, X.; Zhao, H.; Wang, J.; Wu, C.; et al. Arginine, N-carbamylglutamate, and glutamine exert protective effects against oxidative stress in rat intestine. Anim. Nutr. 2016, 2, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Xiao, L.; Liu, G.; Fang, T.; Wu, X.; Jia, G.; Zhao, H.; Chen, X.; Wu, C.; Cai, J.; et al. Dietary arginine and N-carbamylglutamate supplementation enhances the antioxidant statuses of the liver and plasma against oxidative stress in rats. Food Funct. 2016, 7, 2303–2311. [Google Scholar] [CrossRef]
- Liu, G.; Zheng, J.; Gu, K.; Wu, C.; Jia, G.; Zhao, H.; Chen, X.; Wang, J. Calcium-sensing receptor protects intestinal integrity and alleviates the inflammatory response via the Rac1/PLCγ1 signaling pathway. Anim. Biotechnol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Rao, Z.; Li, J.; Shi, B.; Zeng, Y.; Liu, Y.; Sun, Z.; Wu, L.; Sun, W.; Tang, Z. Dietary tryptophan levels impact growth performance and intestinal microbial ecology in weaned piglets via tryptophan metabolites and intestinal antimicrobial peptides. Animals 2021, 11, 817. [Google Scholar] [CrossRef] [PubMed]
- Berstad, A.; Raa, J.; Valeur, J. Tryptophan: ‘essential’ for the pathogenesis of irritable bowel syndrome? Scand. J. Gastroenterol. 2014, 49, 1493–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortamira, N.O.; Seve, B.; Lebreton, Y.; Ganier, P. Effect of dietary tryptophan on muscle, liver and whole-body protein synthesis in weaned piglets: Relationship to plasma insulin. Br. J. Nutr. 1991, 66, 423–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Floc’h, N.; Melchior, D.; Sève, B. Dietary tryptophan helps to preserve tryptophan homeostasis in pigs suffering from lung inflammation. J. Anim. Sci. 2008, 86, 3473–3479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattarai, K.R.; Riaz, T.A.; Kim, H.R.; Chae, H.J. The aftermath of the interplay between the endoplasmic reticulum stress response and redox signaling. Exp. Mol. Med. 2021, 53, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Rees, W.D.; Stahl, M.; Jacobson, K.; Bressler, B.; Sly, L.M.; Vallance, B.A.; Steiner, T.S. Enteroids derived from inflammatory bowel disease patients display dysregulated endoplasmic reticulum stress pathways, leading to differential inflammatory responses and dendritic cell maturation. J. Crohns Colitis 2020, 14, 948–961. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Chen, A.W.; Varner, J.D. A review of the mammalian unfolded protein response. Biotechnol. Bioeng. 2011, 108, 2777–2793. [Google Scholar] [CrossRef] [Green Version]
- Hetz, C. The unfolded protein response: Controlling cell fate decisions under ER stress and beyond. Nat. Rev. Mol. Cell Biol. 2012, 13, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Tabas, I.; Ron, D. Integrating the mechanisms of apoptosis induced by endoplasmic reticulum stress. Nat. Cell Biol. 2011, 13, 184–190. [Google Scholar] [CrossRef] [PubMed]
- McCullough, K.D.; Martindale, J.L.; Klotz, L.O.; Aw, T.Y.; Holbrook, N.J. Gadd153 sensitizes cells to endoplasmic reticulum stress by down-regulating Bcl2 and perturbing the cellular redox state. Mol. Cell. Biol. 2001, 21, 1249–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Zhang, Z.; Ruan, J.; Pan, Y.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 2016, 535, 153–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Zaki, M.H.; Vogel, P.; Gurung, P.; Finlay, B.B.; Deng, W.; Lamkanfi, M.; Kanneganti, T.D. Role of inflammasomes in host defense against Citrobacter rodentium infection. J. Biol. Chem. 2012, 287, 16955–16964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, C.; Duan, F.; Hu, J.; Luo, B.; Huang, B.; Lou, X.; Sun, X.; Li, H.; Zhang, X.; Yin, S.; et al. NLRP3 inflammasome-mediated pyroptosis contributes to the pathogenesis of non-ischemic dilated cardiomyopathy. Redox Biol. 2020, 34, 101523. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Song, J.; Pan, X.; Wan, Y.; Wu, Z.; Lv, S.; Mi, L.; Wang, Y.; Tian, F. Downregulating gasdermin D reduces severe acute pancreatitis associated with pyroptosis. Med. Sci. Monit. 2021, 27, e927968. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Content (%) | |
|---|---|---|
| 7–11 kg | 11–25 kg | |
| Corn | 27.37 | 30.6 |
| Extruded corn | 30.84 | 32 |
| Soybean oil | 2.5 | 1.4 |
| glucose | 2 | 2 |
| Whey powder | 5 | 5 |
| Dehulled soybean meal | 13.24 | 13.04 |
| Soybean protein concentrate | 5 | 5 |
| Extruded soybean | 7 | 5 |
| Fish meal | 3 | 2.5 |
| l-Lysine-HCl | 0.52 | 0.44 |
| dl-Methionine | 0.11 | 0.08 |
| l-Threonine | 0.2 | 0.15 |
| l-Tryptophan | 0.03 | 0.01 |
| l-Alanine | 0.46 | 0.32 |
| Choline chloride | 0.15 | 0.15 |
| Limestone | 0.68 | 0.41 |
| Monocalcium phosphate | 1.35 | 1.35 |
| NaCl | 0.25 | 0.25 |
| Vitamin premix a | 0.05 | 0.05 |
| Mineral premix b | 0.25 | 0.25 |
| Total | 100 | 100 |
| Nutrient level c | ||
| Digestible energy | 3.55 Mcal/kg | 3.49 Mcal/kg |
| Crude protein | 19.72% | 18.65% |
| Calcium | 0.80% | 0.68% |
| Total phosphorus | 0.66% | 0.64% |
| Available phosphorus | 0.48% | 0.46% |
| SID-Lysine | 1.36% | 1.24% |
| SID-Methionine | 0.40% | 0.36% |
| SID-Threonine | 0.80% | 0.73% |
| SID-Tryptophan | 0.23% | 0.20% |
| Target Genes | Forward Primer | Reverse Primer | Accession Number | Temperature (°C) | Product Size (bp) |
|---|---|---|---|---|---|
| TNF-α | CGACTCAGTGCCGAGATCAA | GACCTGCCCAGATTCAGCAA | JF831365.1 | 58 | 60 |
| IL-1β | AAGGCCGCCAAGATATAACTGA | GCCCTCTGGGTATGGCTTTC | NM_001302388.1 | 58 | 71 |
| IL-6 | ATGCTTCCAATCTGGGTTCAA | CACAAGACCGGTGGTGATTCT | AF518322.1 | 58 | 61 |
| IL-8 | ACATCCATGAGGAAGACAGTTTGA | CGGGAACTCCACGCTAGATTC | AB057440.1 | 58 | 70 |
| IL-18 | AGGGACATCAAGCCGTGTTT | CGGTCTGAGGTGCATTATCTGA | AY450287.1 | 58 | 66 |
| GRP78 | GAGATCATCGCCAACGATCA | CAGGAGTGAAGGCCACATATGAC | XM_001927795.5 | 58 | 61 |
| CHOP | CTCCAGATTCCAGTCAGAGTTCTAT | TCTCCTGCTCCTTCTCCTTCAT | NM_001144845.1 | 58 | 60 |
| ATF6 | TCAGGGACCTGCCAAGTGA | GGGTCAATGAGTGAAGAGAAAGC | XM_021089515.1 | 58 | 68 |
| ERO1α | CGGCGCAGAGGTGCTT | CAACATCACAGGTACAATCATCCA | NM_001137627.1 | 58 | 60 |
| ERO1β | GGGCAAGATACGATGATTCACA | TACTGAGCAGCTGGCGATCTC | XM_013981626.2 | 58 | 50 |
| ASC | CCAAGCCAGCTGGAATCAA | TGCAGTGCTGGTTTGTTGTCT | MK302492.1 | 58 | 58 |
| GSDMD | GGCAGCGTCATTGCATTTC | TGAAGGTTCGCTGCTTCTTGT | XM_021090504.1 | 58 | 91 |
| NLRP3 | TCCCCTGGTCTGCTGGATT | ACTCTTGCCGCTATCCATCTG | NM_001256770.2 | 58 | 61 |
| Caspase-1 | CGACCCCCACCTTGCA | AAGGCATTTTCCAGATTGTGAAC | NM_214162.1 | 58 | 61 |
| Caspase-3 | CCGGAATGGCATGTCGAT | TGAAGGTCTCCCTGAGATTTGC | NM_214131.1 | 58 | 60 |
| Bax | AAGCGCATTGGAGATGAACTG | CACGGCTGCGATCATCCT | XM_003127290.5 | 58 | 59 |
| BCL2 | CCAGCATGCGGCCTCTAT | GACTGAGCAGCGCCTTCAG | AB271960.1 | 58 | 57 |
| β-actin | TGCGGGACATCAAGGAGAA | GCCATCTCCTGCTCGAAGTC | DQ452569.1 | 58 | 59 |
| Item | Treatment 1 | SEM | p-Value 2 | |||||
|---|---|---|---|---|---|---|---|---|
| CONTR | LPS | LPS + 0.2% Trp | LPS + 0.4% Trp | CONTR vs. LPS | Linear | Quadratic | ||
| Villus height, μm | 401.51 | 193.11 | 320.62 | 341.93 | 19.83 | <0.001 | 0.001 | 0.108 |
| Villus area, μm2 | 237,721.19 | 101,927.17 | 177,673.28 | 196,176.90 | 14,306.24 | 0.001 | 0.004 | 0.258 |
| Crypt depth, μm | 254.97 | 274.17 | 212.78 | 191.96 | 11.60 | 0.606 | 0.002 | 0.297 |
| VCR | 1.76 | 0.77 | 1.63 | 1.92 | 0.12 | 0.001 | <0.001 | 0.165 |
| Item | Treatment 1 | SEM | p-Value 2 | |||||
|---|---|---|---|---|---|---|---|---|
| CONTR | LPS | LPS + 0.2% Trp | LPS + 0.4% Trp | CONTR vs. LPS | Linear | Quadratic | ||
| MDA, nmol/mg prot | 0.49 | 1.01 | 0.62 | 0.81 | 0.05 | <0.001 | 0.069 | 0.004 |
| T-SOD, U/mg prot | 75.51 | 51.21 | 67.34 | 64.37 | 2.72 | 0.015 | 0.008 | 0.022 |
| CAT, U/mg prot | 9.24 | 6.02 | 8.45 | 8.64 | 0.34 | 0.007 | <0.001 | 0.015 |
| GSH-Px, U/mg prot | 16.48 | 2.87 | 16.30 | 9.93 | 1.21 | <0.001 | <0.001 | <0.001 |
| H2O2, U/mg prot | 0.48 | 0.88 | 0.47 | 0.59 | 0.04 | <0.001 | <0.001 | <0.001 |
| ROS, pg/mg prot | 0.49 | 1.01 | 0.62 | 0.81 | 1.97 | <0.001 | <0.001 | <0.001 |
| Item | Treatment 1 | SEM | p-Value 2 | |||||
|---|---|---|---|---|---|---|---|---|
| CONTR | LPS | LPS + 0.2% Trp | LPS + 0.4% Trp | CONTR vs. LPS | Linear | Quadratic | ||
| TNF-α, pg/mg prot | 33.15 | 64.52 | 35.19 | 41.92 | 2.99 | <0.001 | <0.001 | <0.001 |
| IL-1β, pg/mg prot | 123.98 | 179.53 | 93.65 | 110.62 | 7.43 | <0.001 | <0.001 | <0.001 |
| IL-6, pg/mg prot | 172.47 | 257.39 | 135.52 | 171.16 | 10.33 | <0.001 | <0.001 | <0.001 |
| IL-8, pg/mg prot | 12.25 | 18.43 | 8.79 | 10.17 | 0.86 | <0.001 | <0.001 | <0.001 |
| IL-10, pg/mg prot | 76.14 | 49.78 | 67.38 | 95.13 | 3.91 | <0.001 | <0.001 | 0.150 |
| IDO, pg/mg prot | 9.20 | 12.11 | 6.72 | 10.39 | 0.48 | <0.001 | 0.008 | <0.001 |
| IFN-γ, pg/mg prot | 14.76 | 21.07 | 11.36 | 18.39 | 0.89 | <0.001 | 0.019 | <0.001 |
| Item | Treatment 1 | SEM | p-Value 2 | |||||
|---|---|---|---|---|---|---|---|---|
| LPS | CONTR | LPS + 0.2% Trp | LPS + 0.4% Trp | CONTR vs. LPS | Linear | Quadratic | ||
| TNF-α | 1.00 | 0.17 | 0.16 | 0.13 | 0.08 | <0.001 | <0.001 | <0.001 |
| IL-1β | 1.00 | 0.55 | 0.51 | 0.59 | 0.05 | <0.001 | 0.002 | 0.007 |
| IL-6 | 1.00 | 0.75 | 0.76 | 0.79 | 0.03 | 0.001 | 0.002 | 0.01 |
| IL-8 | 1.00 | 0.65 | 0.29 | 0.31 | 0.06 | <0.001 | <0.001 | <0.001 |
| IL-18 | 1.00 | 0.34 | 0.35 | 0.37 | 0.06 | <0.001 | <0.001 | <0.001 |
| Item | Treatment 1 | SEM | p-Value 2 | |||||
|---|---|---|---|---|---|---|---|---|
| LPS | CONTR | LPS + 0.2% Trp | LPS + 0.4% Trp | CONTR vs. LPS | Linear | Quadratic | ||
| GRP78 | 1.00 | 0.61 | 0.72 | 0.79 | 0.03 | <0.001 | 0.001 | 0.001 |
| CHOP | 1.00 | 0.62 | 0.40 | 0.41 | 0.05 | <0.001 | <0.001 | <0.001 |
| ATF6 | 1.00 | 0.18 | 0.23 | 0.31 | 0.07 | <0.001 | <0.001 | <0.001 |
| ERO1α | 1.00 | 0.52 | 0.57 | 0.72 | 0.04 | 0.001 | 0.002 | <0.001 |
| ERO1β | 1.00 | 0.66 | 0.76 | 0.71 | 0.03 | <0.001 | <0.001 | 0.049 |
| Caspase-3 | 1.00 | 0.45 | 0.48 | 0.48 | 0.05 | <0.001 | <0.001 | <0.001 |
| Bax | 1.00 | 0.46 | 0.44 | 0.47 | 0.05 | <0.001 | <0.001 | <0.001 |
| BCL2 | 1.00 | 1.49 | 1.62 | 1.56 | 0.07 | 0.012 | 0.001 | 0.012 |
| ASC | 1.00 | 0.06 | 0.08 | 0.1 | 0.08 | <0.001 | <0.001 | <0.001 |
| GSDMD | 1.00 | 0.17 | 0.16 | 0.13 | 0.38 | <0.001 | <0.001 | <0.001 |
| NLRP3 | 1.00 | 0.64 | 0.68 | 0.57 | 0.04 | <0.001 | <0.001 | 0.047 |
| Caspase-1 | 1.00 | 0.16 | 0.19 | 0.25 | 0.36 | <0.001 | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Tao, J.; Lu, J.; Jia, G.; Zhao, H.; Chen, X.; Tian, G.; Cai, J.; Zhang, R.; Wang, J. Dietary Tryptophan Supplementation Improves Antioxidant Status and Alleviates Inflammation, Endoplasmic Reticulum Stress, Apoptosis, and Pyroptosis in the Intestine of Piglets after Lipopolysaccharide Challenge. Antioxidants 2022, 11, 872. https://doi.org/10.3390/antiox11050872
Liu G, Tao J, Lu J, Jia G, Zhao H, Chen X, Tian G, Cai J, Zhang R, Wang J. Dietary Tryptophan Supplementation Improves Antioxidant Status and Alleviates Inflammation, Endoplasmic Reticulum Stress, Apoptosis, and Pyroptosis in the Intestine of Piglets after Lipopolysaccharide Challenge. Antioxidants. 2022; 11(5):872. https://doi.org/10.3390/antiox11050872
Chicago/Turabian StyleLiu, Guangmang, Jingyuan Tao, Jiajia Lu, Gang Jia, Hua Zhao, Xiaoling Chen, Gang Tian, Jingyi Cai, Ruinan Zhang, and Jing Wang. 2022. "Dietary Tryptophan Supplementation Improves Antioxidant Status and Alleviates Inflammation, Endoplasmic Reticulum Stress, Apoptosis, and Pyroptosis in the Intestine of Piglets after Lipopolysaccharide Challenge" Antioxidants 11, no. 5: 872. https://doi.org/10.3390/antiox11050872
APA StyleLiu, G., Tao, J., Lu, J., Jia, G., Zhao, H., Chen, X., Tian, G., Cai, J., Zhang, R., & Wang, J. (2022). Dietary Tryptophan Supplementation Improves Antioxidant Status and Alleviates Inflammation, Endoplasmic Reticulum Stress, Apoptosis, and Pyroptosis in the Intestine of Piglets after Lipopolysaccharide Challenge. Antioxidants, 11(5), 872. https://doi.org/10.3390/antiox11050872