
Ambivalent Roles of Oxidative Stress in Triangular Relationships among Arthropod Vectors, Pathogens and Hosts

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. ROS Generation and Oxidative Stress in Arthropods
2.1. Blood Meal and the Fate of Heme within Arthropods
2.2. Other Biological Sources of ROS
3. Vector Competency and Oxidative Stress
3.1. ROS and Arthropod’s Innate Immunity
3.2. ROS after the Establishment of Infection in Hosts
3.3. ROS and Arthropod Microbiome
4. ROS and Chemical Control of Arthropods
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gubler, D.J. Vector-borne diseases. Rev. Sci. Tech. 2009, 28, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.M.; Oliveira, M.P.; Paes, M.C.; Oliveira, M.F. Modulation of mitochondrial metabolism as a biochemical trait in blood feeding organisms: The redox vampire hypothesis redux. Cell Biol. Int. 2018, 42, 683–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisuzzaman; Islam, M.K.; Alim, M.A.; Miyoshi, T.; Hatta, T.; Yamaji, K.; Matsumoto, Y.; Fujisaki, K.; Tsuji, N. Longistatin, a plasminogen activator, is key to the availability of blood-meals for ixodid ticks. PLoS Pathog. 2011, 7, e1001312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisuzzaman; Islam, M.K.; Alim, M.A.; Miyoshi, T.; Hatta, T.; Yamaji, K.; Matsumoto, Y.; Fujisaki, K.; Tsuji, N. Longistatin is an unconventional serine protease and induces protective immunity against tick infestation. Mol. Biochem. Parasitol. 2012, 182, 45–53. [Google Scholar] [CrossRef]
- Anisuzzaman; Hatta, T.; Miyoshi, T.; Matsubayashi, M.; Islam, M.K.; Alim, M.A.; Anas, M.A.; Hasan, M.M.; Matsumoto, Y.; Yamamoto, Y.; et al. Longistatin in tick saliva blocks advanced glycation end-product receptor activation. J. Clin. Investig. 2014, 124, 4429–4444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alim, M.A.; Tsuji, N.; Miyoshi, T.; Islam, M.K.; Hatta, T.; Fujisaki, K. Legumains from the hard tick Haemaphysalis longicornis play modulatory roles in blood feeding and gut cellular remodelling and impact on embryogenesis. Int. J. Parasitol. 2009, 39, 97–107. [Google Scholar] [CrossRef]
- Hatta, T.; Tsuji, N.; Miyoshi, T.; Islam, M.K.; Alim, M.A.; Yamaji, K.; Anisuzzaman; Fujisaki, K. Leucine aminopeptidase, HlLAP, from the ixodid tick Haemaphysalis longicornis, plays vital roles in the development of oocytes. Parasitol. Int. 2010, 59, 286–289. [Google Scholar] [CrossRef]
- Graça-Souza, A.V.; Maya-Monteiro, C.; Paiva-Silva, G.O.; Braz, G.R.C.; Paes, M.C.; Sorgine, M.H.F.; Oliveira, M.F.; Oliveira, P.L. Adaptations against heme toxicity in blood-feeding arthropods. Insect Biochem. Mol. Biol. 2006, 36, 322–335. [Google Scholar] [CrossRef]
- Anisuzzaman; Islam, M.K.; Miyoshi, T.; Alim, M.A.; Hatta, T.; Yamaji, K.; Matsumoto, Y.; Fujisaki, K.; Tsuji, N. Longistatin, a novel EF-hand protein from the ixodid tick Haemaphysalis longicornis, is required for acquisition of host blood-meals. Int. J. Parasitol. 2010, 40, 721–729. [Google Scholar] [CrossRef]
- GBD 2017 DALYs and HALE Collaborators Global. Regional, and national disability-adjusted life-years (DALYs) for 359 diseases and injuries and healthy life expectancy (HALE) for 195 countries and territories, 1990-2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1859–1922. [Google Scholar] [CrossRef] [Green Version]
- Dantas-Torres, F.; Otranto, D. Best Practices for Preventing Vector-Borne Diseases in Dogs and Humans. Trends Parasitol. 2016, 32, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Whiten, S.R.; Eggleston, H.; Adelman, Z.N. Ironing out the Details: Exploring the Role of Iron and Heme in Blood-Sucking Arthropods. Front. Physiol. 2017, 8, 1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galay, R.L.; Umemiya-Shirafuji, R.; Mochizuki, M.; Fujisaki, K.; Tanaka, T. Iron metabolism in hard ticks (Acari: Ixodidae): The antidote to their toxic diet. Parasitol. Int. 2015, 64, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Toh, S.Q.; Glanfield, A.; Gobert, G.N.; Jones, M.K. Heme and blood-feeding parasites: Friends or foes? Parasit. Vectors 2010, 3, 108. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, E.P.; Talactac, M.R.; Fujisaki, K.; Tanaka, T. The case for oxidative stress molecule involvement in the tick-pathogen interactions -an omics approach. Dev. Comp. Immunol. 2019, 100, 103409. [Google Scholar] [CrossRef]
- Chaitanya, R.K.; Shashank, K.; Sridevi, P. Oxidative stress in invertebrate systems. In Free Radicals and Diseases; Ahmad, R., Ed.; InTech: London, UK, 2016. [Google Scholar]
- Sabadin, G.A.; Xavier, M.A.; Vaz, I.D.S., Jr. Control of redox homeostasis in tick blood feeding. Acta Sci. Vet. 2019, 47, 1678. [Google Scholar] [CrossRef] [Green Version]
- Fraga, A.; Moraes, J.; da Silva, J.R.; Costa, E.P.; Menezes, J.; da Silva Vaz, I.; Logullo, C.; da Fonseca, R.N.; Campos, E. Inorganic polyphosphates regulate hexokinase activity and reactive oxygen species generation in mitochondria of Rhipicephalus (Boophilus) microplus embryo. Int. J. Biol. Sci. 2013, 9, 842–852. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.A. Traffic of the tick embryo basic protein during embryogenesis of the camel tick Hyalomma dromedarii (Acari: Ixodidae). Exp. Appl. Acarol. 1998, 22, 481–495. [Google Scholar] [CrossRef]
- Santos, V.T.; Ribeiro, L.; Fraga, A.; de Barros, C.M.; Campos, E.; Moraes, J.; Fontenele, M.R.; Araújo, H.M.; Feitosa, N.M.; Logullo, C.; et al. The embryogenesis of the tick Rhipicephalus (Boophilus) microplus: The establishment of a new chelicerate model system. Genesis 2013, 51, 803–818. [Google Scholar] [CrossRef]
- Frahm, S.; Anisuzzaman, A.; Prodjinotho, U.F.; Vejzagić, N.; Verschoor, A.; Prazeres da Costa, C. A novel cell-free method to culture Schistosoma mansoni from cercariae to juvenile worm stages for in vitro drug testing. PLoS Negl. Trop. Dis. 2019, 13, e0006590. [Google Scholar] [CrossRef] [Green Version]
- Sterkel, M.; Oliveira, J.H.M.; Bottino-Rojas, V.; Paiva-Silva, G.O.; Oliveira, P.L. The Dose Makes the Poison: Nutritional Overload Determines the Life Traits of Blood-Feeding Arthropods. Trends Parasitol. 2017, 33, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Hatta, T.; Tsuji, N.; Miyoshi, T.; Alim, M.A.; Islam, M.K.; Fujisaki, K. Leucine aminopeptidase in the ixodid tick Haemaphysalis longicornis: Endogenous expression profiles in midgut. J. Vet. Med. Sci. 2009, 71, 589–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyoshi, T.; Tsuji, N.; Islam, M.K.; Alim, M.A.; Hatta, T.; Huang, X.; Fujisaki, K. A set of serine proteinase paralogs are required for blood-digestion in the ixodid tick Haemaphysalis longicornis. Parasitol. Int. 2008, 57, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Alim, A.M.; Tsuji, N.; Miyoshi, T.; Khyrul Islam, M.; Huang, X.; Motobu, M.; Fujisaki, K. Characterization of asparaginyl endopeptidase, legumain induced by blood feeding in the ixodid tick Haemaphysalis longicornis. Insect Biochem. Mol. Biol. 2007, 37, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Alim, M.A.; Tsuji, N.; Miyoshi, T.; Islam, M.K.; Huang, X.; Hatta, T.; Fujisaki, K. HlLgm2, a member of asparaginyl endopeptidases/legumains in the midgut of the ixodid tick Haemaphysalis longicornis, is involved in blood-meal digestion. J. Insect Physiol. 2008, 54, 573–585. [Google Scholar] [CrossRef]
- Perner, J.; Sobotka, R.; Sima, R.; Konvickova, J.; Sojka, D.; de Oliveira, P.L.; Hajdusek, O.; Kopacek, P. Acquisition of exogenous haem is essential for tick reproduction. Elife 2016, 5, e12318. [Google Scholar] [CrossRef]
- de Araújo, A.P.O.; Telleria, E.L.; da Dutra, J.M.F.; Júlio, R.M.; Traub-Csekö, Y.M. Disruption of the peritrophic matrix by exogenous chitinase feeding reduces fecundity in Lutzomyia longipalpis females. Mem. Inst. Oswaldo Cruz 2012, 107, 543–545. [Google Scholar] [CrossRef] [Green Version]
- Hatta, T.; Miyoshi, T.; Matsubayashi, M.; Islam, M.K.; Alim, M.A.; Anisuzzaman; Yamaji, K.; Fujisaki, K.; Tsuji, N. Semi-artificial mouse skin membrane feeding technique for adult tick, Haemaphysalis longicornis. Parasit. Vectors 2012, 5, 263. [Google Scholar] [CrossRef] [Green Version]
- Sonenshine, D.E.; Anderson, J.M. Mouthparts and Digestive System: Anatomy and Molecular biology of Feeding and Digestion. In Biology of Ticks, 2nd ed.; Sonenshine, D.E., Roe, R.M., Eds.; Oxford University Press: New York, NY, USA, 2014; Volume 1. [Google Scholar]
- Alim, M.A.; Tsuji, N.; Miyoshi, T.; Islam, M.K.; Hatta, T.; Yamaji, K.; Fujisaki, K. Developmental stage- and organ-specific expression profiles of asparaginyl endopeptidases/legumains in the ixodid tick Haemaphysalis longicornis. J. Vet. Med. Sci. 2008, 70, 1363–1366. [Google Scholar] [CrossRef] [Green Version]
- Narasimhan, S.; Sukumaran, B.; Bozdogan, U.; Thomas, V.; Liang, X.; De Ponte, K.; Marcantonio, N.; Koski, R.A.; Anderson, J.F.; Kantor, F.; et al. A tick antioxidant facilitates the Lyme disease agent’s successful migration from the mammalian host to the arthropod vector. Cell Host Microbe 2007, 2, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Narasimhan, S.; Fikrig, E. Tick microbiome: The force within. Trends Parasitol. 2015, 31, 315–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, D.R.J.; Rosa, R.M.; Moraes, J.; Campos, E.; Logullo, C.; Da Silva Vaz, I.; Masuda, A. Relationship between glutathione S-transferase, catalase, oxygen consumption, lipid peroxidation and oxidative stress in eggs and larvae of Boophilus microplus (Acarina: Ixodidae). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 146, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, E.P.; Shimazaki, K.; Niihara, H.; Umemiya-Shirafuji, R.; Fujisaki, K.; Tanaka, T. Expression analysis of glutathione S-transferases and ferritins during the embryogenesis of the tick Haemaphysalis longicornis. Heliyon 2020, 6, e03644. [Google Scholar] [CrossRef] [PubMed]
- DeJong, R.J.; Miller, L.M.; Molina-Cruz, A.; Gupta, L.; Kumar, S.; Barillas-Mury, C. Reactive oxygen species detoxification by catalase is a major determinant of fecundity in the mosquito Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2007, 104, 2121–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strode, C.; Steen, K.; Ortelli, F.; Ranson, H. Differential expression of the detoxification genes in the different life stages of the malaria vector Anopheles gambiae. Insect Mol. Biol. 2006, 15, 523–530. [Google Scholar] [CrossRef]
- Reeves, M.A.; Hoffmann, P.R. The human selenoproteome: Recent insights into functions and regulation. Cell Mol. Life Sci. 2009, 66, 2457–2478. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.S.; Oliveira, P.L.; Barja-Fidalgo, C.; Daffre, S. Production of reactive oxygen species by hemocytes from the cattle tick Boophilus microplus. Exp. Parasitol. 2001, 99, 66–72. [Google Scholar] [CrossRef]
- Kumar, S.; Christophides, G.K.; Cantera, R.; Charles, B.; Han, Y.S.; Meister, S.; Dimopoulos, G.; Kafatos, F.C.; Barillas-Mury, C. The role of reactive oxygen species on Plasmodium melanotic encapsulation in Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2003, 100, 14139–14144. [Google Scholar] [CrossRef] [Green Version]
- Surachetpong, W.; Pakpour, N.; Cheung, K.W.; Luckhart, S. Reactive oxygen species-dependent cell signaling regulates the mosquito immune response to Plasmodium falciparum. Antioxid. Redox Signal. 2011, 14, 943–955. [Google Scholar] [CrossRef] [Green Version]
- Corby-Harris, V.; Drexler, A.; Watkins de Jong, L.; Antonova, Y.; Pakpour, N.; Ziegler, R.; Ramberg, F.; Lewis, E.E.; Brown, J.M.; Luckhart, S.; et al. Activation of Akt signaling reduces the prevalence and intensity of malaria parasite infection and lifespan in Anopheles stephensi mosquitoes. PLoS Pathog. 2010, 6, e1001003. [Google Scholar] [CrossRef]
- Budachetri, K.; Kumar, D.; Crispell, G.; Beck, C.; Dasch, G.; Karim, S. The tick endosymbiont Candidatus Midichloria mitochondrii and selenoproteins are essential for the growth of Rickettsia parkeri in the Gulf Coast tick vector. Microbiome 2018, 6, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otali, D.; Novak, R.J.; Wan, W.; Bu, S.; Moellering, D.R.; De Luca, M. Increased production of mitochondrial reactive oxygen species and reduced adult life span in an insecticide-resistant strain of Anopheles gambiae. Bull. Entomol. Res. 2014, 104, 323–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajan, V.; Puthur, R. Activation of the Oxidative Stress in Culex quinquefasciatus by the Augmented Production of Reactive Oxygen Species (ROS) in response to Stachytarpheta jamaicensis Exposure. J. Commun. Dis. 2021, 53, 43–51. [Google Scholar]
- Beerntsen, B.T.; James, A.A.; Christensen, B.M. Genetics of mosquito vector competence. Microbiol. Mol. Biol. Rev. 2000, 64, 115–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisuzzaman; Islam, M.K.; Alim, M.A.; Tsuji, N. Longistatin, an EF-hand Ca2+-binding protein from vector tick: Identification, purification, and characterization. Methods Mol. Biol. 2013, 963, 127–146. [Google Scholar]
- Anisuzzaman; Khyrul Islam, M.; Abdul Alim, M.; Miyoshi, T.; Hatta, T.; Yamaji, K.; Matsumoto, Y.; Fujisaki, K.; Tsuji, N. Longistatin, a novel plasminogen activator from vector ticks, is resistant to plasminogen activator inhibitor-1. Biochem. Biophys. Res. Commun. 2011, 413, 599–604. [Google Scholar] [CrossRef]
- de la Fuente, J.; Antunes, S.; Bonnet, S.; Cabezas-Cruz, A.; Domingos, A.G.; Estrada-Peña, A.; Johnson, N.; Kocan, K.M.; Mansfield, K.L.; Nijhof, A.M.; et al. Tick-Pathogen Interactions and Vector Competence: Identification of Molecular Drivers for Tick-Borne Diseases. Front. Cell Infect. Microbiol. 2017, 7, 114. [Google Scholar] [CrossRef] [Green Version]
- Hajdušek, O.; Síma, R.; Ayllón, N.; Jalovecká, M.; Perner, J.; de la Fuente, J.; Kopáček, P. Interaction of the tick immune system with transmitted pathogens. Front. Cell Infect. Microbiol. 2013, 3, 26. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, N.; Miyoshi, T.; Battsetseg, B.; Matsuo, T.; Xuan, X.; Fujisaki, K. A cysteine protease is critical for Babesia spp. transmission in Haemaphysalis ticks. PLoS Pathog. 2008, 4, e1000062. [Google Scholar] [CrossRef] [Green Version]
- Budachetri, K.; Crispell, G.; Karim, S. Amblyomma maculatum SECIS binding protein 2 and putative selenoprotein P are indispensable for pathogen replication and tick fecundity. Insect Biochem. Mol. Biol. 2017, 88, 37–47. [Google Scholar] [CrossRef]
- Ha, E.M.; Oh, C.T.; Ryu, J.H.; Bae, Y.S.; Kang, S.W.; Jang, I.H.; Brey, P.T.; Lee, W.J. An antioxidant system required for host protection against gut infection in Drosophila. Dev. Cell 2005, 8, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muralidharan, S.; Mandrekar, P. Cellular stress response and innate immune signaling: Integrating pathways in host defense and inflammation. J. Leukoc. Biol. 2013, 94, 1167–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina-Cruz, A.; DeJong, R.J.; Charles, B.; Gupta, L.; Kumar, S.; Jaramillo-Gutierrez, G.; Barillas-Mury, C. Reactive oxygen species modulate Anopheles gambiae immunity against bacteria and Plasmodium. J. Biol. Chem. 2008, 283, 3217–3223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalil, S.P.; da Rosa, R.D.; Capelli-Peixoto, J.; Pohl, P.C.; de Oliveira, P.L.; Fogaça, A.C.; Daffre, S. Immune-related redox metabolism of embryonic cells of the tick Rhipicephalus microplus (BME26) in response to infection with Anaplasma marginale. Parasit. Vectors 2017, 10, 613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashida, H.; Mimuro, H.; Ogawa, M.; Kobayashi, T.; Sanada, T.; Kim, M.; Sasakawa, C. Cell death and infection: A double-edged sword for host and pathogen survival. J. Cell Biol. 2011, 195, 931–942. [Google Scholar] [CrossRef] [Green Version]
- Villar, M.; Ayllón, N.; Alberdi, P.; Moreno, A.; Moreno, M.; Tobes, R.; Mateos-Hernández, L.; Weisheit, S.; Bell-Sakyi, L.; de la Fuente, J. Integrated Metabolomics, Transcriptomics and Proteomics Identifies Metabolic Pathways Affected by Anaplasma phagocytophilum Infection in Tick Cells. Mol. Cell Proteom. 2015, 14, 3154–3172. [Google Scholar] [CrossRef] [Green Version]
- Villar, M.; Popara, M.; Bonzón-Kulichenko, E.; Ayllón, N.; Vázquez, J.; de la Fuente, J. Characterization of the tick-pathogen interface by quantitative proteomics. Ticks Tick Borne Dis. 2012, 3, 154–158. [Google Scholar] [CrossRef]
- Chen, T.H.; Wu, Y.J.; Hou, J.N.; Chiang, Y.H.; Cheng, C.C.; Sifiyatun, E.; Chiu, C.H.; Wang, L.C.; Chen, W.J. A novel p53 paralogue mediates antioxidant defense of mosquito cells to survive dengue virus replication. Virology 2018, 519, 156–169. [Google Scholar] [CrossRef]
- Chen, T.H.; Lo, Y.P.; Yang, C.F.; Chen, W.J. Additive protection by antioxidant and apoptosis-inhibiting effects on mosquito cells with dengue 2 virus infection. PLoS Negl. Trop. Dis. 2012, 6, e1613. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.H.; Chiang, Y.H.; Hou, J.N.; Cheng, C.C.; Sofiyatun, E.; Chiu, C.H.; Chen, W.J. XBP1-Mediated BiP/GRP78 Upregulation Copes with Oxidative Stress in Mosquito Cells during Dengue 2 Virus Infection. Biomed Res. Int. 2017, 2017, 3519158. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, J.H.M.; Talyuli, O.A.C.; Goncalves, R.L.S.; Paiva-Silva, G.O.; Sorgine, M.H.F.; Alvarenga, P.H.; Oliveira, P.L. Catalase protects Aedes aegypti from oxidative stress and increases midgut infection prevalence of dengue but not Zika. PLoS Negl. Trop. Dis. 2017, 11, e0005525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.H.; Tang, P.; Yang, C.F.; Kao, L.H.; Lo, Y.P.; Chuang, C.K.; Shih, Y.T.; Chen, W.J. Antioxidant defense is one of the mechanisms by which mosquito cells survive dengue 2 viral infection. Virology 2011, 410, 410–417. [Google Scholar] [CrossRef] [Green Version]
- Basu, M.; Courtney, S.C.; Brinton, M.A. Arsenite-induced stress granule formation is inhibited by elevated levels of reduced glutathione in West Nile virus-infected cells. PLoS Pathog. 2017, 13, e1006240. [Google Scholar] [CrossRef] [Green Version]
- Kuzmenko, Y.V.; Smirnova, O.A.; Ivanov, A.V.; Starodubova, E.S.; Karpov, V.L. Nonstructural Protein 1 of Tick-Borne Encephalitis Virus Induces Oxidative Stress and Activates Antioxidant Defense by the Nrf2/ARE Pathway. Intervirology 2016, 59, 111–117. [Google Scholar] [CrossRef]
- Hernandez, E.P.; Talactac, M.R.; Vitor, R.J.S.; Yoshii, K.; Tanaka, T. An Ixodes scapularis glutathione S-transferase plays a role in cell survival and viability during Langat virus infection of a tick cell line. Acta Trop. 2021, 214, 105763. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, J.M.; Perera, R.; Roumani, A.M.; Hedrick, V.E.; Inerowicz, H.D.; Hill, C.A.; Kuhn, R.J. Changes in the Proteome of Langat-Infected Ixodes scapularis ISE6 Cells: Metabolic Pathways Associated with Flavivirus Infection. PLoS Negl. Trop. Dis. 2016, 10, e0004180. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, J.M.; Gulia-Nuss, M.; Kuhn, R.J.; Hill, C.A. RNAi reveals proteins for metabolism and protein processing associated with Langat virus infection in Ixodes scapularis (black-legged tick) ISE6 cells. Parasit. Vectors 2017, 10, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulda, S.; Gorman, A.M.; Hori, O.; Samali, A. Cellular stress responses: Cell survival and cell death. Int. J. Cell Biol. 2010, 2010, 214074. [Google Scholar] [CrossRef] [Green Version]
- Busby, A.T.; Ayllón, N.; Kocan, K.M.; Blouin, E.F.; de la Fuente, G.; Galindo, R.C.; Villar, M.; de la Fuente, J. Expression of heat shock proteins and subolesin affects stress responses, Anaplasma phagocytophilum infection and questing behaviour in the tick, Ixodes scapularis. Med. Vet. Entomol. 2012, 26, 92–102. [Google Scholar] [CrossRef]
- Villar, M.; Ayllón, N.; Busby, A.T.; Galindo, R.C.; Blouin, E.F.; Kocan, K.M.; Bonzón-Kulichenko, E.; Zivkovic, Z.; Almazán, C.; Torina, A.; et al. Expression of Heat Shock and Other Stress Response Proteins in Ticks and Cultured Tick Cells in Response to Anaplasma spp. Infection and Heat Shock. Int. J. Proteom. 2010, 2010, 657261. [Google Scholar] [CrossRef] [Green Version]
- Alberdi, P.; Cabezas-Cruz, A.; Prados, P.E.; Rayo, M.V.; Artigas-Jerónimo, S.; de la Fuente, J. The redox metabolic pathways function to limit Anaplasma phagocytophilum infection and multiplication while preserving fitness in tick vector cells. Sci. Rep. 2019, 9, 13236. [Google Scholar] [CrossRef] [PubMed]
- Kocan, K.M.; Zivkovic, Z.; Blouin, E.F.; Naranjo, V.; Almazán, C.; Mitra, R.; de la Fuente, J. Silencing of genes involved in Anaplasma marginale-tick interactions affects the pathogen developmental cycle in Dermacentor variabilis. BMC Dev. Biol. 2009, 9, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahlon, A.; Ojogun, N.; Ragland, S.A.; Seidman, D.; Troese, M.J.; Ottens, A.K.; Mastronunzio, J.E.; Truchan, H.K.; Walker, N.J.; Borjesson, D.L.; et al. Anaplasma phagocytophilum Asp14 is an invasin that interacts with mammalian host cells via its C terminus to facilitate infection. Infect. Immun. 2013, 81, 65–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, L.V.; Gordon, J.I. Commensal host-bacterial relationships in the gut. Science 2001, 292, 1115–1118. [Google Scholar] [CrossRef] [PubMed]
- Kinross, J.M.; Darzi, A.W.; Nicholson, J.K. Gut microbiome-host interactions in health and disease. Genome Med. 2011, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Budachetri, K.; Browning, R.E.; Adamson, S.W.; Dowd, S.E.; Chao, C.-C.; Ching, W.M.; Karim, S. An insight into the microbiome of the Amblyomma maculatum (Acari: Ixodidae). J. Med. Entomol. 2014, 51, 119–129. [Google Scholar] [CrossRef]
- Cirimotich, C.M.; Ramirez, J.L.; Dimopoulos, G. Native microbiota shape insect vector competence for human pathogens. Cell Host Microbe 2011, 10, 307–310. [Google Scholar] [CrossRef] [Green Version]
- Cirimotich, C.M.; Dong, Y.; Clayton, A.M.; Sandiford, S.L.; Souza-Neto, J.A.; Mulenga, M.; Dimopoulos, G. Natural microbe-mediated refractoriness to Plasmodium infection in Anopheles gambiae. Science 2011, 332, 855–858. [Google Scholar] [CrossRef] [Green Version]
- Crispell, G.; Budachetri, K.; Karim, S. Rickettsia parkeri colonization in Amblyomma maculatum: The role of superoxide dismutases. Parasit. Vectors 2016, 9, 291. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Budachetri, K.; Meyers, V.C.; Karim, S. Assessment of tick antioxidant responses to exogenous oxidative stressors and insight into the role of catalase in the reproductive fitness of the Gulf Coast tick, Amblyomma maculatum. Insect Mol. Biol. 2016, 25, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.J. Bacterial-modulated signaling pathways in gut homeostasis. Sci. Signal. 2008, 1, pe24. [Google Scholar] [CrossRef] [PubMed]
- Champion, C.J.; Xu, J. The impact of metagenomic interplay on the mosquito redox homeostasis. Free Radic. Biol. Med. 2017, 105, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Romoli, O.; Gendrin, M. The tripartite interactions between the mosquito, its microbiota and Plasmodium. Parasit. Vectors 2018, 11, 200. [Google Scholar] [CrossRef] [Green Version]
- Baron, S.; Barrero, R.A.; Black, M.; Bellgard, M.I.; van Dalen, E.M.S.; Fourie, J.; Maritz-Olivier, C. Differentially expressed genes in response to amitraz treatment suggests a proposed model of resistance to amitraz in R. decoloratus ticks. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Li, T.; Zhang, L.; Gao, X.; Liu, N. Permethrin induction of multiple cytochrome P450 genes in insecticide resistant mosquitoes, Culex quinquefasciatus. Int. J. Biol. Sci. 2013, 9, 863–871. [Google Scholar] [CrossRef] [Green Version]
- Oliver, S.V.; Brooke, B.D. The Role of Oxidative Stress in the Longevity and Insecticide Resistance Phenotype of the Major Malaria Vectors Anopheles arabiensis and Anopheles funestus. PLoS ONE 2016, 11, e0151049. [Google Scholar] [CrossRef] [Green Version]
- Champion, C.J.; Xu, J. Redox state affects fecundity and insecticide susceptibility in Anopheles gambiae. Sci. Rep. 2018, 8, 13054. [Google Scholar] [CrossRef]
- Koodalingam, A.; Deepalakshmi, R.; Ammu, M.; Rajalakshmi, A. Effects of NeemAzal on marker enzymes and hemocyte phagocytic activity of larvae and pupae of the vector mosquito Aedes aegypti. J. Asia. Pac. Entomol. 2014, 17, 175–181. [Google Scholar] [CrossRef]
- Thanigaivel, A.; Senthil-Nathan, S.; Vasantha-Srinivasan, P.; Edwin, E.S.; Ponsankar, A.; Selin-Rani, S.; Pradeepa, V.; Chellappandian, M.; Kalaivani, K.; Abdel-Megeed, A.; et al. Chemicals isolated from Justicia adhatoda Linn reduce fitness of the mosquito, Aedes aegypti L. Arch. Insect. Biochem. Physiol. 2017, 94, e21384. [Google Scholar] [CrossRef] [Green Version]
- Agwunobi, D.O.; Pei, T.; Wang, X.; Yu, Z.; Liu, J.Z. Expression profiles of glutathione S-transferases genes in semi-engorged Haemaphysalis longicornis (Acari: Ixodidae) exposed to Cymbopogon citratus essential oil. Sys. Appl. Acarol. 2020, 25, 918–930. [Google Scholar]



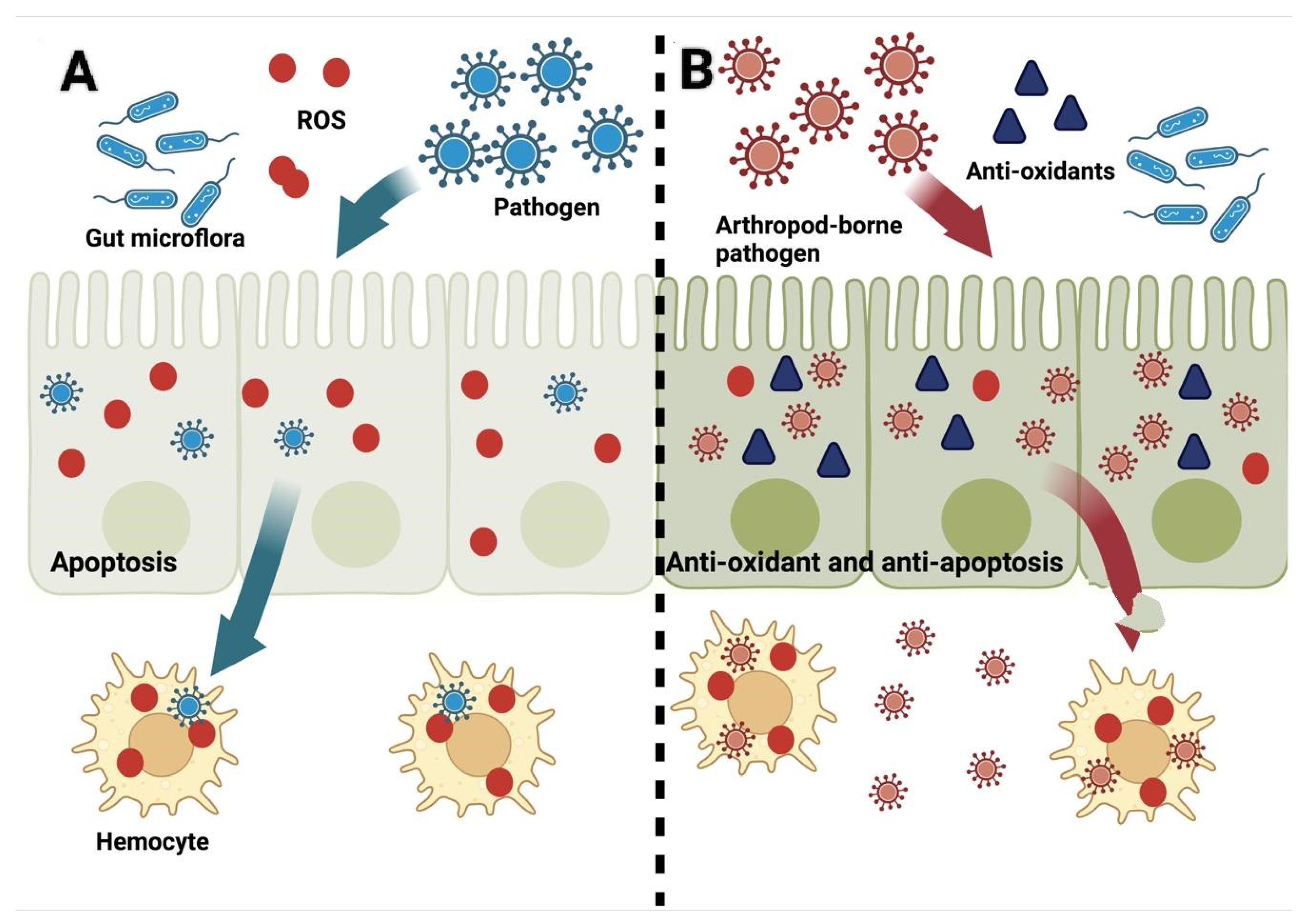{kind=link}
{kind=link}
| Biological Activity | Reference |
|---|---|
| Activity for reproduction and growth | |
| Oogenesis | [36] |
| Embryonic development | [34,35] |
| Hatching and molting | [38] |
| Larval development | [34] |
| Normal homeostasis | |
| Flight activity | [37] |
| Nervous system activity | [16] |
| Cellular activity | [38] |
| Pathogen infection | |
| Microbial killing by ROS | [39] |
| Melanocytic encapsulation | [40] |
| Immune signaling | [41,42] |
| Arthropod microbiome | |
| Enterobacter production | [43] |
| Arthropod control | |
| Metabolism of acaricide/insecticide | [16] |
| Insecticide resistance | [44] |
| Phytochemical control | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandez, E.P.; Anisuzzaman; Alim, M.A.; Kawada, H.; Kwofie, K.D.; Ladzekpo, D.; Koike, Y.; Inoue, T.; Sasaki, S.; Mikami, F.; et al. Ambivalent Roles of Oxidative Stress in Triangular Relationships among Arthropod Vectors, Pathogens and Hosts. Antioxidants 2022, 11, 1254. https://doi.org/10.3390/antiox11071254
Hernandez EP, Anisuzzaman, Alim MA, Kawada H, Kwofie KD, Ladzekpo D, Koike Y, Inoue T, Sasaki S, Mikami F, et al. Ambivalent Roles of Oxidative Stress in Triangular Relationships among Arthropod Vectors, Pathogens and Hosts. Antioxidants. 2022; 11(7):1254. https://doi.org/10.3390/antiox11071254
Chicago/Turabian StyleHernandez, Emmanuel Pacia, Anisuzzaman, Md Abdul Alim, Hayato Kawada, Kofi Dadzie Kwofie, Danielle Ladzekpo, Yuki Koike, Takahiro Inoue, Sana Sasaki, Fusako Mikami, and et al. 2022. "Ambivalent Roles of Oxidative Stress in Triangular Relationships among Arthropod Vectors, Pathogens and Hosts" Antioxidants 11, no. 7: 1254. https://doi.org/10.3390/antiox11071254
APA StyleHernandez, E. P., Anisuzzaman, Alim, M. A., Kawada, H., Kwofie, K. D., Ladzekpo, D., Koike, Y., Inoue, T., Sasaki, S., Mikami, F., Matsubayashi, M., Tanaka, T., Tsuji, N., & Hatta, T. (2022). Ambivalent Roles of Oxidative Stress in Triangular Relationships among Arthropod Vectors, Pathogens and Hosts. Antioxidants, 11(7), 1254. https://doi.org/10.3390/antiox11071254