From the degenerate PCR screening onto the fosmid libraries of an istamycin aminoglycoside-producing
S. tenjimariensis ATCC 31603, a total of four positive clones were selected then thoroughly sequenced: two were found to be involved in the istamycin aminoglycoside biosynthesis by sequence alignment using BLAST online search tool (
http://www.ncbi.nih.gov/BLAST; accessed on 1 July 2022); the remaining two surely encode family I GT; however; they are supposed to act on different kinds of aglycones. Hence, an orf44 (designated with NdeI and XhoI cut) was selected and amplified by PCR, and the amplified ORF of interest (annotated as
stSPGT) was 1176 base pairs in length, which encodes a predicted 41.78-kDa protein (deposited as GenBank Accession No. MT770755). In silico BLAST analysis of the deduced protein revealed dominant homology with GTs originating from the soil dwelling Actinomycete (including
Micromonospora,
Streptomyces,
Actinoplanes and
Frankia species). In addition, the presence of a significant conserved motif (as nucleotide-sugar donor binding domain) in the
C-terminal region indicates that the designated orf44 product StSPGT is a member of the family I GTs.
3.1. Enzymatic Biosynthesis of Simple Phenolic Glycosides Using Recombinant StSPGT
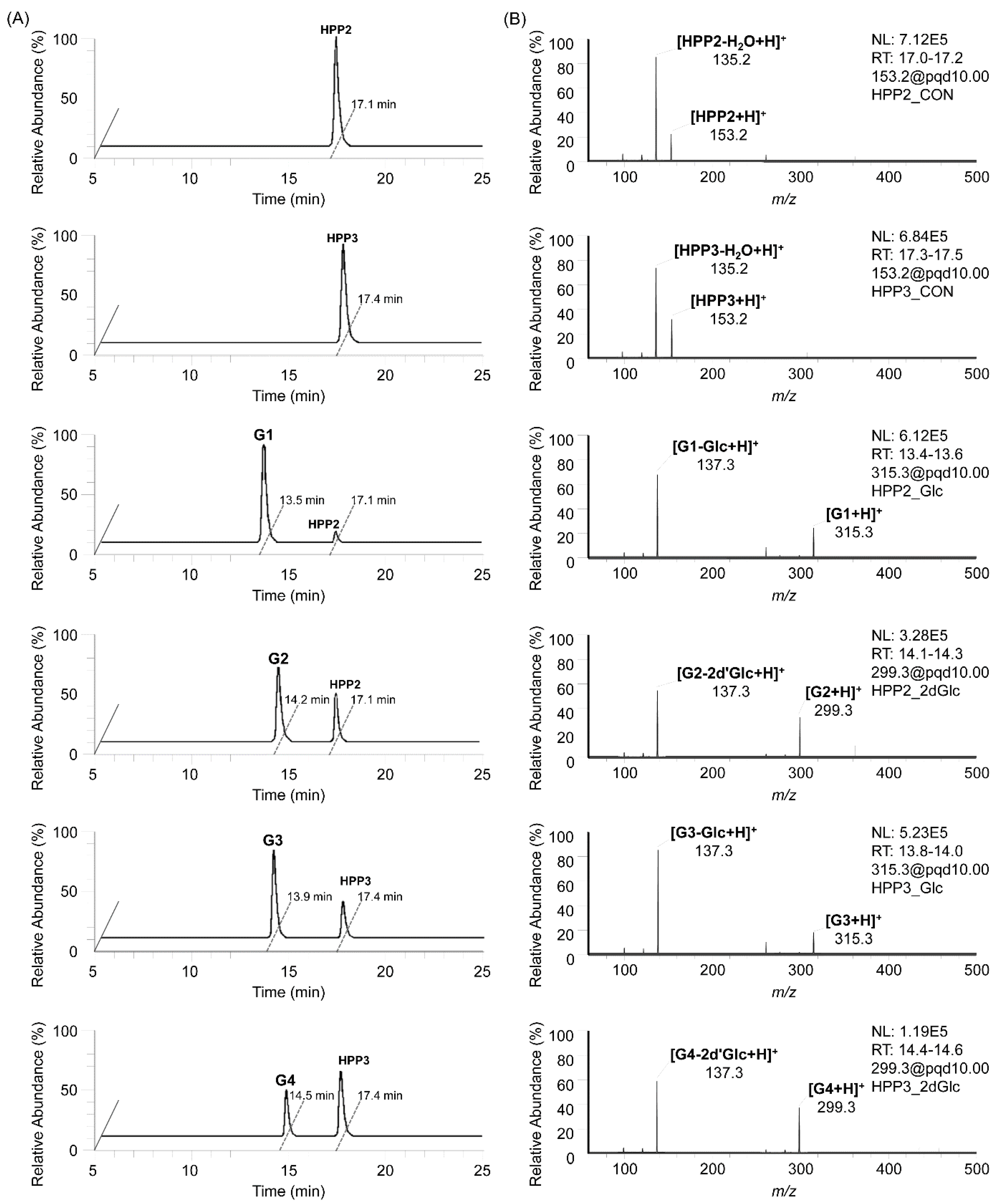
The in vitro enzymatic formation of the corresponding SP glycosides from SP aglycones, together with glycosyl donors, was traced using HPLC-MS/MS analyses, and their MS fragmentation patterns (pairing of mass transitions specific to glycone molecular ion and its dominant aglycone ion corresponding to the loss of a glycosyl moiety) characteristic of the SP glycosides clearly indicated the attachment of glycosyl moieties onto the SP glycosyl acceptors, representing them as a series of SP-O-glycosides (
Figure 2); on the HPLC-MS/MS chromatograms obtained from the StSPGT reactants in which UDP-Glc and TDP-2′dGlc were supplemented as the glycosyl donor towards two SP acceptor substrates (HPP2 and HPP3), a total four peaks of glycosylated products were separately traced, thus possibly indicating the substrate-flexible attachment of glycosyl moieties onto both SP aglycones. Further in vitro enzymatic reactions were performed, using two additional common nucleotide-activated sugars (i.e., UDP-GlcA and UDP-GlcNAc) as glycosyl donor substrates of StSPGT to examine whether varied glycosylation occurs or not. However, throughout the overnight reaction, we were not able to trace any targeted SP glycosides as products in the reactant, thus indicating the StSPGT’s limited substrate specificity for the attachment of both Glc and 2′dGlc moieties onto the SP acceptor substrate. The ratios of corresponding SP glycoside to SP aglycone in the reactants were averaged to 97% glycoside
G1 (retention time at 13.5 min) from HPP2 with UDP-Glc, 62% glycoside
G2 (retention time at 14.2 min) from HPP2 with TDP-2′dGlc, 71% glycoside
G3 (retention time at 13.9 min) from HPP3 with UDP-Glc and 44% glycoside
G4 (retention time at 14.5 min) from HPP3 with TDP-2′dGlc (
Figure 2A). Three different sets of mass transitions specific to both SPs and their corresponding glycosides were certainly acquired to detect the transition of the protonated precursor ion to the dominant product ion (i.e., 153.2 [M+H]
+ > 135.2 [M-H
2O+H]
+ as a dehydrated product ion equivalent for both HPP2 and HPP3 aglycones; 315.3 > 137.3 [M-Glc+H]
+ as an aglycone product ion equivalent for glycosides G1 and G3; 299.3 > 137.3 [M-2′dGlc+H]
+ as an aglycone product ion equivalent for both glycosides G2 and G4) (
Figure 2B). Indeed, the recombinant StSPGT seems to recruit HPP2 as a favorable acceptor substrate prior to HPP3, thus revealing its catalytic preference (together with acceptor promiscuity) for the transfer of a glycosyl moiety (glucose or 2′-deoxyglucose) towards SPs. The reverse, namely with respect to the glycosyl donors, also represents that the enzyme is likely to prefer UDP-Glc to TDP-2′dGlc; 97 and 71% conversions from UDP-Glc-supplemented reactions vs. 62 and 44% from TDP-2′dGlc-supplemented ones.
3.2. Structural Elucidation of New Simple Phenolic Glycosides
In order to obtain and purify enough quantities of SP glycosides for the complete structural elucidation of the glycosyl decoration position onto these glycoside products and the anomeric configuration of attached glycosyl moieties, a normal-phase silica cartridge built in the CombiFlash Rf MPLC system (Teledyne ISCO, Lincoln, NE, USA) was employed. After multiple scale-up reactions followed by MPLC chromatographic separation, a total of four different SP glycosides were obtained as white powder: glycoside
G1 (9.7 mg), glycoside
G2 (6.3 mg), glycoside
G3 (6.8 mg) and glycoside
G4 (3.9 mg). Both HR-MS and NMR instrumental analyses were further performed for the structural elucidation of these SP glycosides, and their structural features were finally described (see
Supplementary Tables S1–S4): (i) The
13C-NMR spectra of glycosides displayed nine aglycone carbon signals and six carbon signals (as glycosyl group) clearly, and the appearance of proton peaks representing hexosyl moiety within the range of
δH 2.11 to 5.05 confirmed that each glycoside contains a glycosyl moiety attached to HPP2 or HPP3 aglycone. (ii) In the
1H-NMR spectra, all the glycoside products (
G1 to
G4) displayed common anomeric proton signals between
δH 4.95 to 5.05 (equivalent to the equatorial resonance instead of up fielded-axial one), and the typical observation of noticeable up-field
13C glycosylation shifts at each C2-/C3-carbon revealed that a glycosyl group was attached to a free hydroxyl function of the propanoyl moiety, instead being attached to an C3′-hydroxyl function adjacent to phenyl one. (iii) A small coupling constant (J) between 2.6 to 2.9 Hz found at the anomeric proton of all glycosides clearly indicated the introduction of each glycosyl moiety with an α-type glycosidic bond. (iv) The maintenance of proton signals characteristic of each glycosyl moiety (i.e., additional methylene signal [
δH 2.25 and 2.13/
δC 38.1] at C2″-position of glycosides
G2 and
G4) confirmed the glycosyl moieties derived from the glycosyl donors were intactly transferred onto the aglycone without further modification (
Supplementary Tables S1–S4). Therefore, the enzymatic products
G1 to
G4 were unequivocally assigned as hydroxyphenyl-2-propanoyl-
O-α-glucoside (HPP2G), hydroxyphenyl-2-propanoyl-
O-α-2″-deoxyglucoside (HPP2DG), hydroxyphenyl-3-propanoyl-
O-α-glucoside (HPP3G) and hydroxyphenyl-3-propanoyl-
O-α-2″-deoxyglucoside (HPP3DG) respectively, and all were even previously unprecedented SP glycosides (
Figure 3). Accordingly, the recombinant StSPGT has been shown to attach a series of glycosyl donors with strict regiospecificity at the C2- or C3-hydroxy function present at the propanoyl moiety onto the designated SP aglycone.
3.3. Kinetic Features of Recombinant StSPGT
The optimum pH and temperature for the glycosyl-transferring reaction of recombinant StSPGT were investigated using HPP2 and UDP-Glc as an acceptor and a donor substrate, respectively. The enzyme is likely to be stable within the pH range of 5.5 to 8.0 (optimum set at pH 7.4), and the maximum activity was found at 30 °C (data not shown). To learn more details about the enzymatic function of a recombinant StSPGT, we investigated and determined its bio-catalytic properties using two nucleotide sugars as the glycosyl donor substrates; the production velocities of the corresponding glycosides were plotted against the varied glycosyl donor concentrations, instead of with fixed acceptor concentrations. The
Km and
kcat values for UDP-Glc donor substrate of the enzyme was averaged to be 0.16 mM and 19.41 min
−1, respectively, for HPP2 and 0.22 mM and 16.87 min
−1, respectively, for HPP3 (
Table 1 and
Supplementary Figure S1). The
Km values of recombinant StSPGT for the Glc-transferring activity onto HPP2 and HPP3 were both rigid within the range of 0.16 to 0.22 mM and comparable; however, they were noticeably lower than those determined for another 2′dGlc-transferring substrate. Indeed, the enzyme displayed the lowest
Km value (0.16 mM) for the glucosylation of HPP2 compared to those for HPP3 (0.22 mM), displaying StSPGT’s preference for HPP2 as the acceptor substrate over HPP3. These apparent
Km values appear to be comparable to those reported for other microbial- or plant-origin simple phenolic glycosyltransferases [
1,
16,
22,
28]. Depending upon the donor substrates, several fold differences were found in the
Km values; 0.16 and 0.22 mM when a Glc moiety was incorporated into each aglycone vs. 0.41 and 0.50 mM when a 2′dGlc-moiety was. In particular, the glycosyl donor supplement, especially UDP-Glc onto StSPGT’s enzymatic reactions with HPP2 aglycone, gave the highest turnover number (
kcat = 19.41 min
−1) (
Table 1 and
Supplementary Figure S1). However, the incorporation of another glycosyl donor (TDP-2′dGlc) into the reactions resulted in more remarkably increased
Km and decreased
kcat values, hence clearly demonstrating UDP-Glc as a favorable glycosyl donor substrate. The thorough kinetic analyses revealed that the incorporation of both Glc and 2′dGlc moieties onto SP acceptors can be bio-catalyzed by StSPGT, but the latter had catalytic efficiencies only one-fourth as high as for the UDP-Glc donor, as judged by
kcat/
Km values (
Table 1). Indeed, the
kcat values obtained from UDP-Glc towards HPP2 (19.41 min
−1) and HPP3 (16.87 min
−1) were considerably high, thereby indicating StSPGT’s higher catalytic efficiency (107.83 min
−1 mM
−1 for HPP2 and 76.68 min
−1 mM
−1 for HPP3) compared with that derived from another glycosyl donor; it is obvious that StSPGT can utilize two different nucleotide sugars as glycosyl donors as well as both SP aglycones as glycosyl acceptors, undoubtedly indicating its promiscuity towards pairs of substrates. Based on the kinetic data obtained (
Table 1) and the structural features of the SP aglycones (
Figure 1 and
Figure 3), we were able to speculate on the relationship between the substrate structure and the enzyme’s catalytic activity. This region-specific StSPGT can distinguish, albeit with different substrate spectra and catalytic efficiencies, the positional difference of a hydroxyl function (as C2- or C3-hydroxy) attached onto the propanoyl moiety of each HPP2 and HPP3, and therefore the three-dimensional structure of StSPGT might provide valuable insights into the plausible protein–ligand (or catalytic site-ligand) interactions.
3.4. Antioxidant, Anti-Melanogenic and Anti-Wrinkle Properties
After elucidating the structural features of the SP glycosides obtained, their biological activities (including cytotoxicity, antioxidant, anti-melanogenic and anti-aging activities) were evaluated by comparison with those of both parental aglycones and the positive controls (i.e., L-ascorbic acid, kojic acid, hydroquinone, α-arbutin and oleanolic acid). The treatment concentrations of SP aglycones and their glycosides for the bioassays were selected in advance by monitoring their cytotoxicity against the B16F10 murine melanoma cell line using α-arbutin as the reference. Cell viability was measured after 48 h treatments with different concentrations (0.5, 1.0, 5.0, 10 and 20 mM) and is summarized in
Figure 4A (overall depicted in
Supplementary Figure S2); a reference compound, α-arbutin, displays a slight change (less than 5% of the control) of cell viability only at the highest 20 mM (equivalent to 0.6% content) concentration. In 2015, the European Scientific Committee on Consumer Safety issued a report on the safety of α-arbutin; its cosmetic use is safe for consumers, with concentrations up to 0.5% in skin-care products [
15]. Previous publication has also revealed that α-arbutin has no cytotoxicity for the B16F10 cells if the concentrations are less than 5.0 mM concentrations [
29], thus accordant with the observations herein. Quite similar to the result of α-arbutin, all the glycosides also present a very high percentage of viability with increasing concentrations up to the maximum tested, whereas both aglycones exhibit significantly decreased cell viabilities at concentrations lower than 1.0 mM in B16F10 cells, thus suggesting the SP glycosylation seems to considerably alleviate the cytotoxicity of parental SP aglycones. Accordingly, the further bioassays were conducted for all test samples at concentrations less than the 1.0 mM level.
The in vitro antioxidant activity of SP aglycones and their corresponding glycosides was evaluated through their DPPH-driven radical scavenging abilities in comparison with a well-known antioxidant, L-ascorbic acid. The IC
50 values obtained from a total of four kinds of glycosides were examined to be within 22.4 to 24.1 μM, comparable to that (23.0 μM) obtained from the positive control L-ascorbic acid. In the latest publication, the IC
50 value of L-ascorbic acid was evaluated at 22.0 μM through its DPPH radical scavenging abilities, very similar to the foregoing results [
30]. On the other hand, two SP aglycones provided significantly lower IC
50 values (18.5 μM for HPP2 and 19.3 μM for HPP3, respectively) than the positive control, indicating a substantial radical scavenging capacity (
Figure 4B). Therefore, compared with L-ascorbic acid, the series of SP glycosides obtained still showed comparable DPPH radical scavenging ability, hence implying a considerable antioxidant activity remained in the SP glycosides of interest. Although the mean IC
50 values of HPP2G and HPP2DG appear to be a little lower than those of HPP3G and HPP3DG, respectively, there was, however, no statistically significant difference found within (also between) these glycoside groups. Next, we implemented a DCFDA cell-based ROS detection assay [
31] to determine whether the SP aglycones and their glycoside analogs function in vivo as antioxidants against intracellular ROS synthesis in the above-described B16F10 cells or not. The TBHP-induced cells were further incubated with different concentrations (5.0, 10 and 20 μM) of SPs and their glycosides for varied time periods (10, 30 min and 1 h). As shown in
Figure 4C, the TBHP-exposure induced an approximately 1.8-fold increase in the intracellular ROS level compared with the basal ROS level.
As expected, treatments of both SP aglycones and their glycosides were likely to attenuate the TBHP-mediated ROS level evenly in a concentration-dependent manner. In addition, their treatments at fixed 20 μM with extended time periods (up to 1 h) also led to the reduced intracellular ROS levels in a time-dependent manner (
Supplementary Figure S3). These results clearly demonstrated that the SPs and their glycosides possess antioxidant activity, in a time- and concentration-dependent manner, by scavenging DPPH radicals as well as the intracellular ROS. Interestingly, even though the results obtained from cell-based ROS assay are almost consistent with those from the DPPH assay, HPP2-type glycosides appeared to act as more potent ROS-scavenging antioxidants than HPP3-type glycosides (
Figure 4C); the maximal ROS-scavenging activities of HPP2G and HPP2DG (52.0 and 54.3%, respectively) were notably higher than those of HPP3G and HPP3DG (48.9 and 45.7%, respectively). Although their ROS-scavenging activities were entirely inferior to the parental aglycones, the positional glycosylation at C2- or C3-hydroxyl function onto their propanoyl moieties resulted in the distinctive ROS-scavenging results. After all, the conformational changes caused by the glycosyl attachment (or shielding) of the free C3-hydroxy group (not at C2-position) tend to weaken ROS-scavenging antioxidant potentials.
The comparative tyrosinase inhibitory activities of SPs and SP glycosides were evaluated and expressed as IC
50 values; kojic acid (one of the famous skin-whitening agents) was used as a positive control and hydroquinone (also known as a skin-depigmenting agent with the strongest tyrosinase inhibitory activity) was used as a comparison for in vitro tyrosinase inhibition [
32]. As illustrated in
Figure 4D, the IC
50 value of positive control kojic acid was evaluated as 76.5 μM; kojic acid has previously been proven to have stronger tyrosinase inhibition ability than many other commercial inhibitors (including α-arbutin and L-ascorbic acid). Among the compounds tested, the most potent inhibition was shown by hydroquinone (10.3 μM), followed by HPP2, HPP2 glycosides, HPP3 and HPP3 glycosides in that order. Indeed, the IC
50 values of HPP2 and HPP3 aglycone were determined to be 11.6 and 23.4 μM, respectively, indicating about 6.6-fold and 3.3-fold higher tyrosinase inhibitory effects, respectively, than kojic acid. In particular, an HPP2 aglycone was the most potent tyrosinase inhibitor, of which IC
50 value seems to be comparable to that of hydroquinone. Moreover, different form the above-described antioxidant results, where both SP aglycones are categorized into the same group, the tyrosinase inhibitory activities of both SP aglycones were markedly different. In case of SP glycosides, a similar inhibitory tendency described just above was still verified; IC
50 values of HPP2G and HPP2DG were averaged to be 14.8 and 13.8 μM, respectively, while those of HPP3G and HPP3DG were distinctively 29.1 and 30.9 μM, respectively (
Figure 4D). It was evident from these results that not only the existence of C2-hydroxy function onto a propanoyl moiety of the SP aglycone but also the glycosyl decoration at the C2-position are likely to be responsible for an enhanced tyrosinase inhibitory activity of the corresponding compounds. However, there was no difference in tyrosinase inhibitory activities depending on the type of glycoside (Glc or 2d′Glc). Therefore, it was certainly determined that the glycosyl attachment at the SP’s C2-hydroxyl group does not attenuate the parental activity appreciably, thus suggesting both HPP2 aglycone and the corresponding glycosides as promising tyrosinase inhibitors, applicable as skin-depigmenting dermatological agents. Further studies on molecular docking and dynamic simulation are requisite for understanding the physical and biochemical bases of HPP2 glycosides towards tyrosinase inhibition [
33]. Melanin biosynthesis is a stress response of melanocytes, which can cause melanosis, such as freckles, brown spots, melanoma, senile plaques and even melanocytoma [
15,
30]. To confirm whether the in vitro tyrosinase inhibition found in the preceding experiment was still correlated with the inhibition of the in vivo melanin biosynthesis, we investigated the inhibitory effects of the SPs and their corresponding glycosides on the α-MSH-induced melanin biosynthesis in B16F10 cells. Treatment with α-MSH induced an approximately 2.2-fold increase in the intracellular melanin content (
Figure 4E). When it came to the positive control α-arbutin, inhibition activities of α-MSH-induced intracellular melanin biosynthesis turned out to be about 20.3%, and HPP2 aglycone exhibited the highest inhibitory potential with 50.3%. Additionally, SP glycosides all were in-between, showing inhibitory activities within a range between 22.4 and 46.5%, and the HPP3 aglycone was ranked the lowest (21.7%) (
Figure 4E). Furthermore, the α-MSH-induced melanogeneses were all reversed with concentration-dependent manner (
Supplementary Figure S4). These findings were almost in harmony with the preceding tyrosinase inhibitory results, thus demonstrating that HPP2 and their glycosides (i.e., HPP2G and HPP2DG), albeit with their varied activities, are not only able to inhibit tyrosinase activity in vitro but also undoubtedly suppressed in vivo melanogenesis.
Elastase, a protease, is responsible for the breakdown of elastin in the extracellular matrix (ECM). Elastase can break down elastin along with other substrates (i.e., collagen and fibronectin). In order to decelerate the tangible impacts of skin-aging (i.e., wrinkles, freckles and sagging), the degradation of elastin substrate derived from the elastase activity should be interrupted, thereby terminating the depletion of skin elasticity. Herein, the positive control oleanolic acid displayed an IC
50 value of 24.9 μM (
Figure 4F). A previous study has also shown that the IC
50 value of oleanolic acid (still represented as a positive control) for the elastase inhibitory experiments was 11.7 μg/mL, equivalent to a molar concentration of 25.7 μM [
34]. Hence, it was found that this converted molar concentration was quite resemblant to ours (24.9 μM). In addition, the IC
50 values of both SP aglycones against the porcine elastase were averaged to be 95.3 and 99.1 μM, respectively, whereas those of HPP3G and HPP3DG were 31.5 and 30.7 μM, respectively, hence indicating that the inhibitory activities of these samples against elastase appeared to be lower than that of oleanolic acid. Contrary to this, a series of HPP2 glycosides are likely to exert the best inhibitory activity against elastase with IC
50 values (22.1 and 25.3 μM, respectively) lower than (or comparable to) that of the positive control oleanolic acid (
Figure 4F), thus clearly demonstrating these HPP2 glycosides with enhanced anti-melanogenic and antioxidant activities to be prospective anti-wrinkle cosmetic hits.









{kind=link}
{kind=link}
{kind=link}
{kind=link}