
Physiological Performance, Antioxidant and Immune Status, Columnaris Resistance, and Growth of Nile Tilapia That Received Alchemilla vulgaris-Supplemented Diets

 ,
, 
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tested Compounds and Chemicals
2.2. Experimental Fish
2.3. Diet Formulation
2.4. Growth Performance Assessment
weight gain percentage = [(Final average body weight-Initial average body weight)/Initial average body weight] × 100
where weight = total weight of fish (g) and L = length of fish (cm), which is measured from the tip of the snout to the end of the middle caudal fin.
2.5. Blood and Tissue Sampling
2.6. Estimation of Hematological Parameters
2.7. Biochemical Analysis
2.7.1. Hepatorenal Function Indicators
2.7.2. Immune System Response
2.7.3. Oxidant/Antioxidant Stress Biomarkers
2.8. Pathogenic Bacteria and Immersion Challenge Test
2.9. Statistical Analysis
3. Results
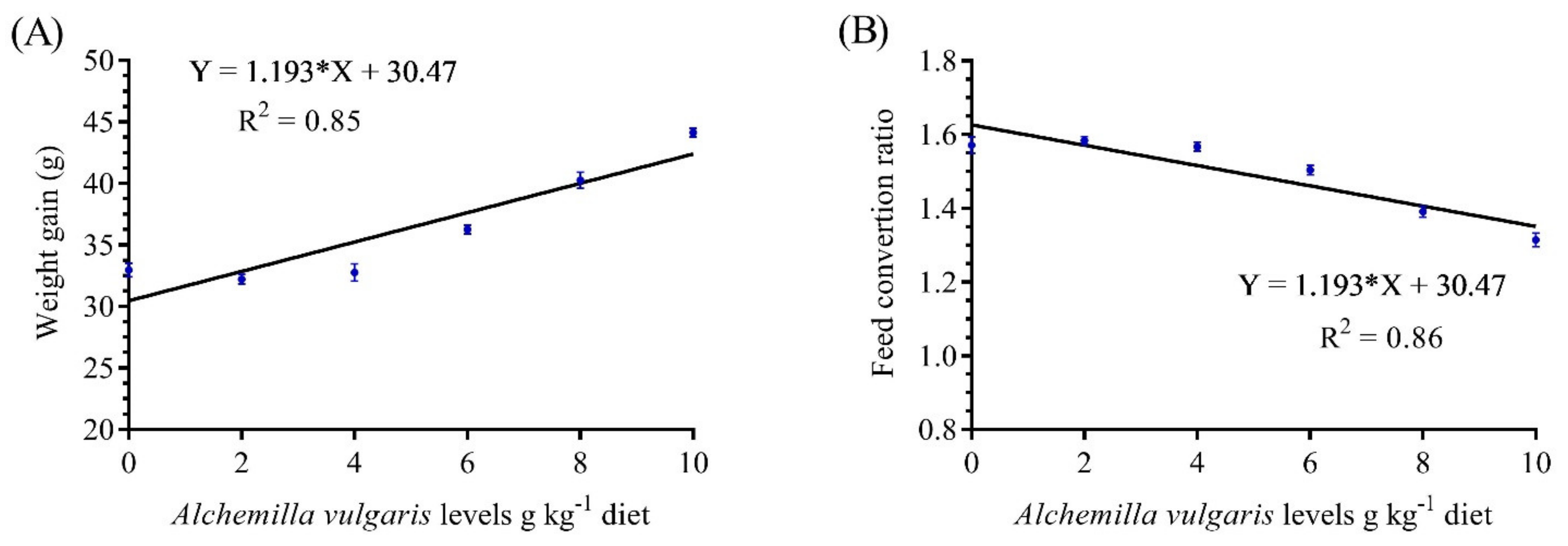
3.1. Growth Performance and Feed Utilization
3.2. Hematological Indices
3.3. Hepatorenal Function Indicators
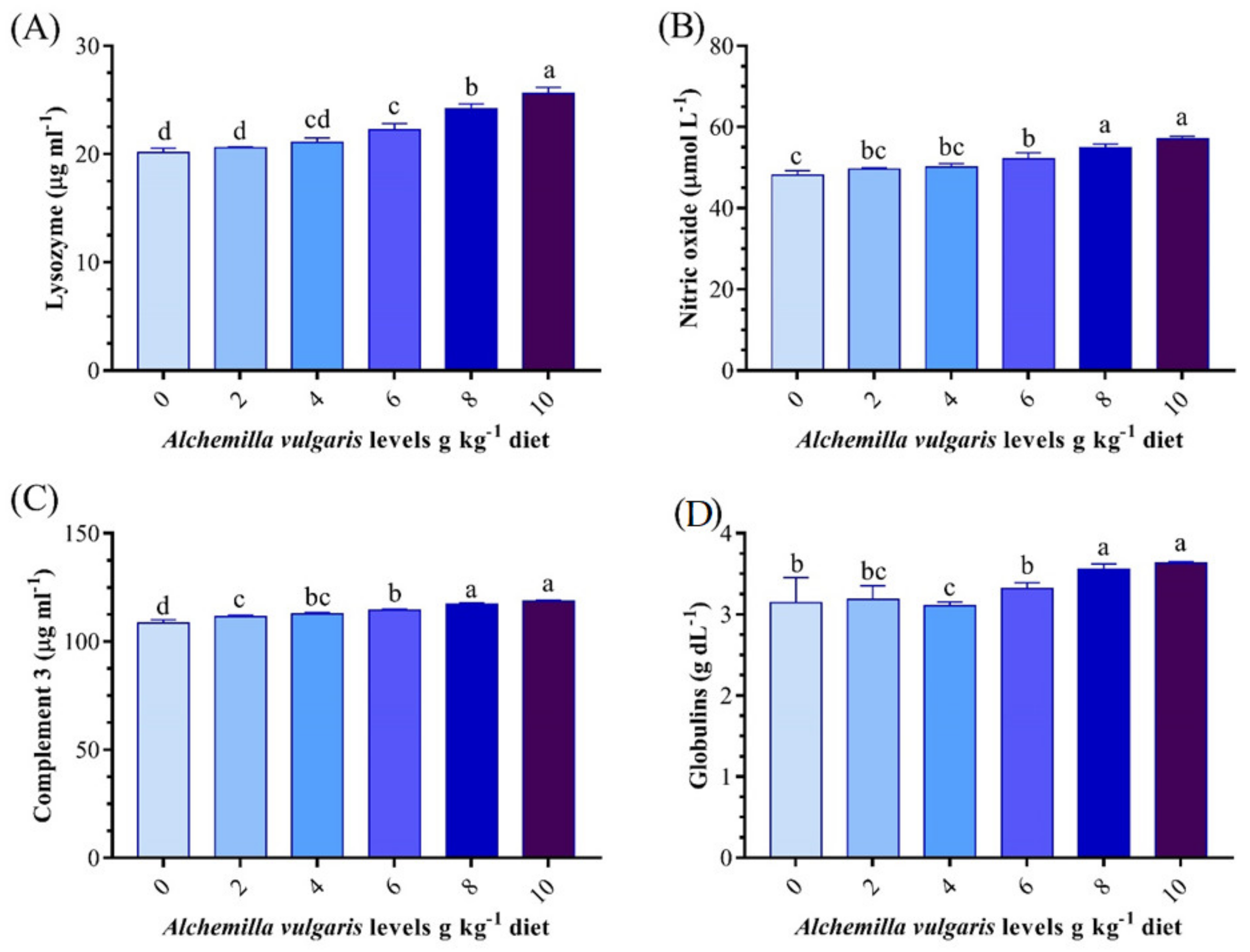
3.4. Innate Immune Response
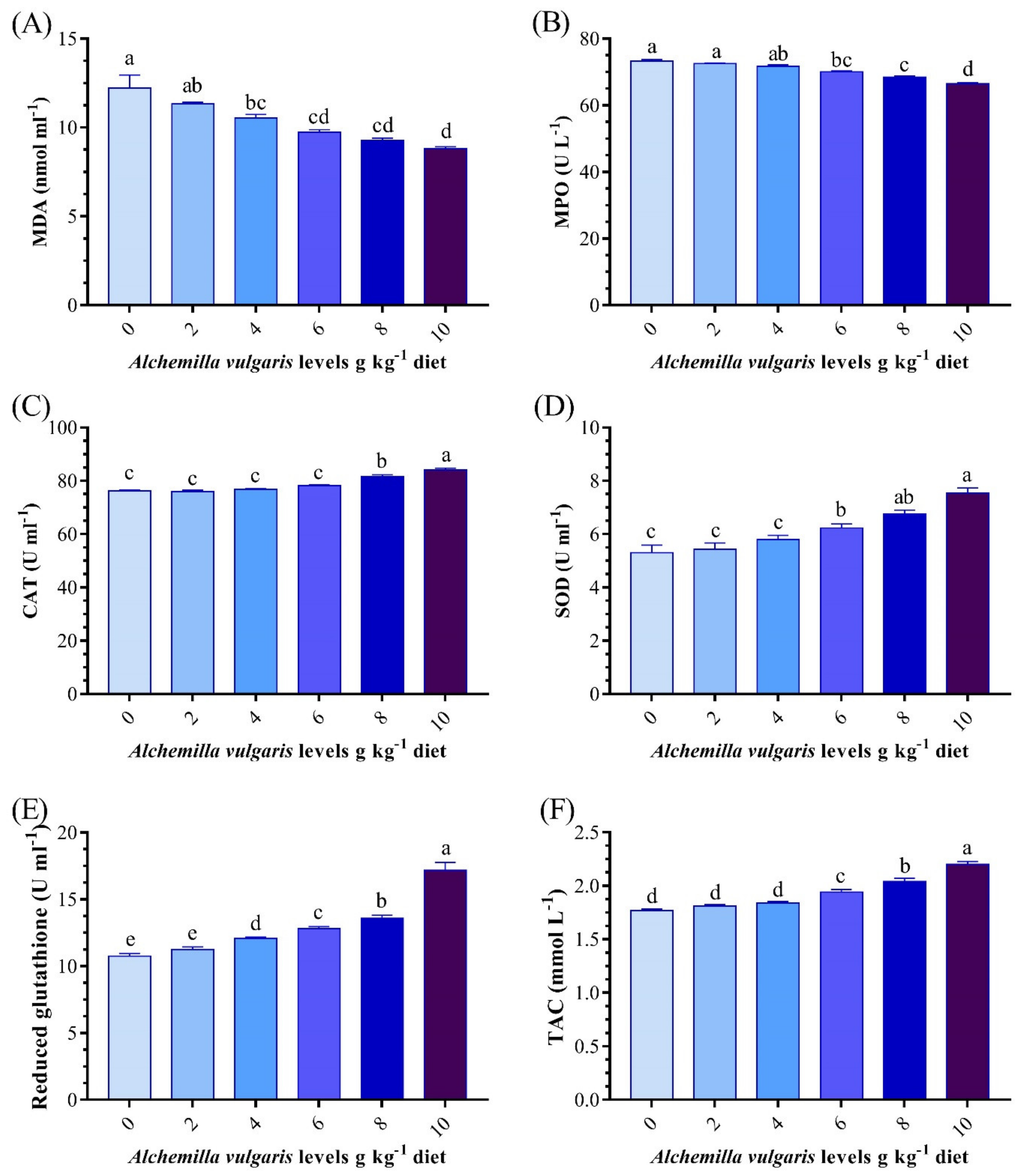
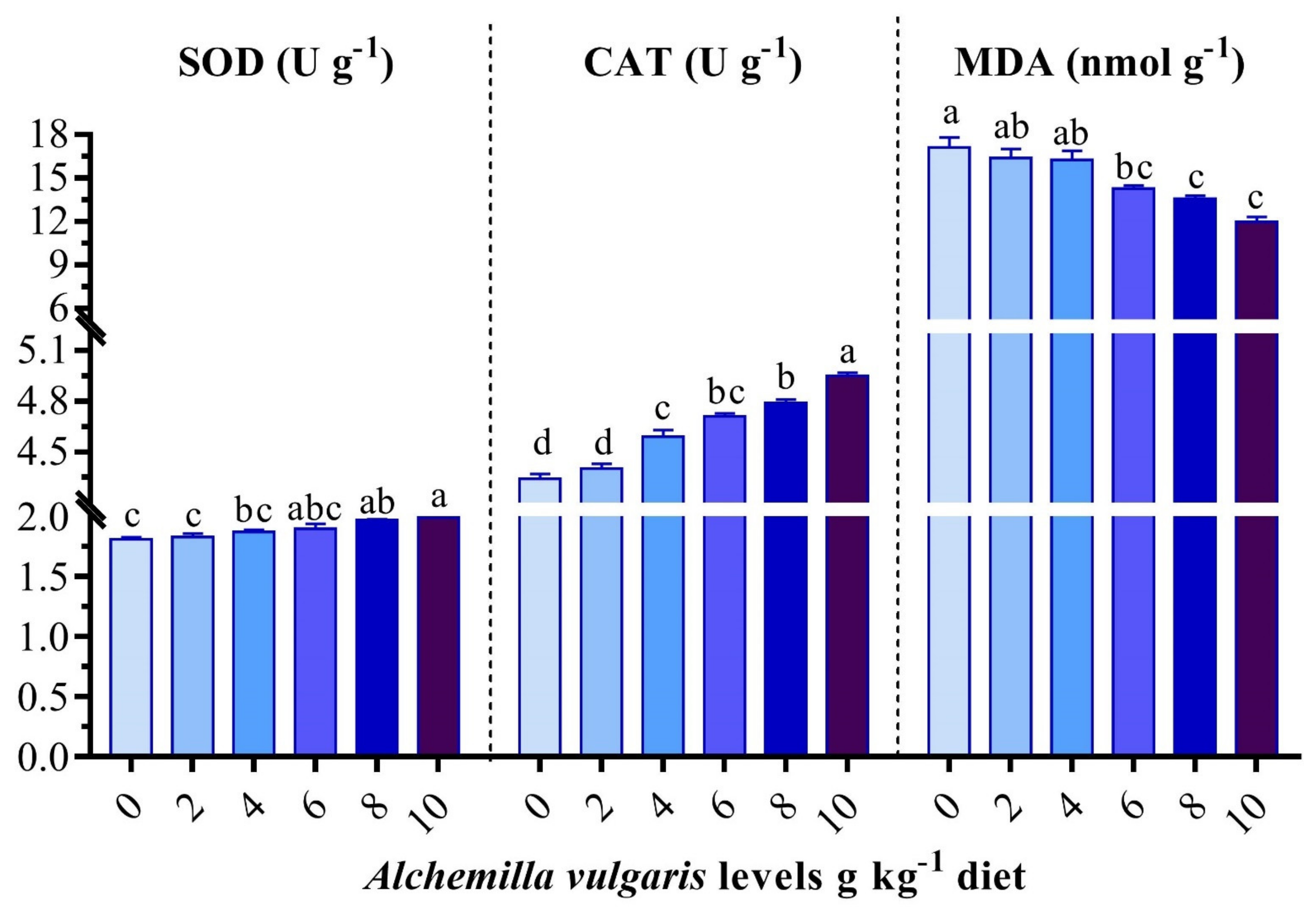
3.5. Antioxidant and Oxidative Stress Biomarkers
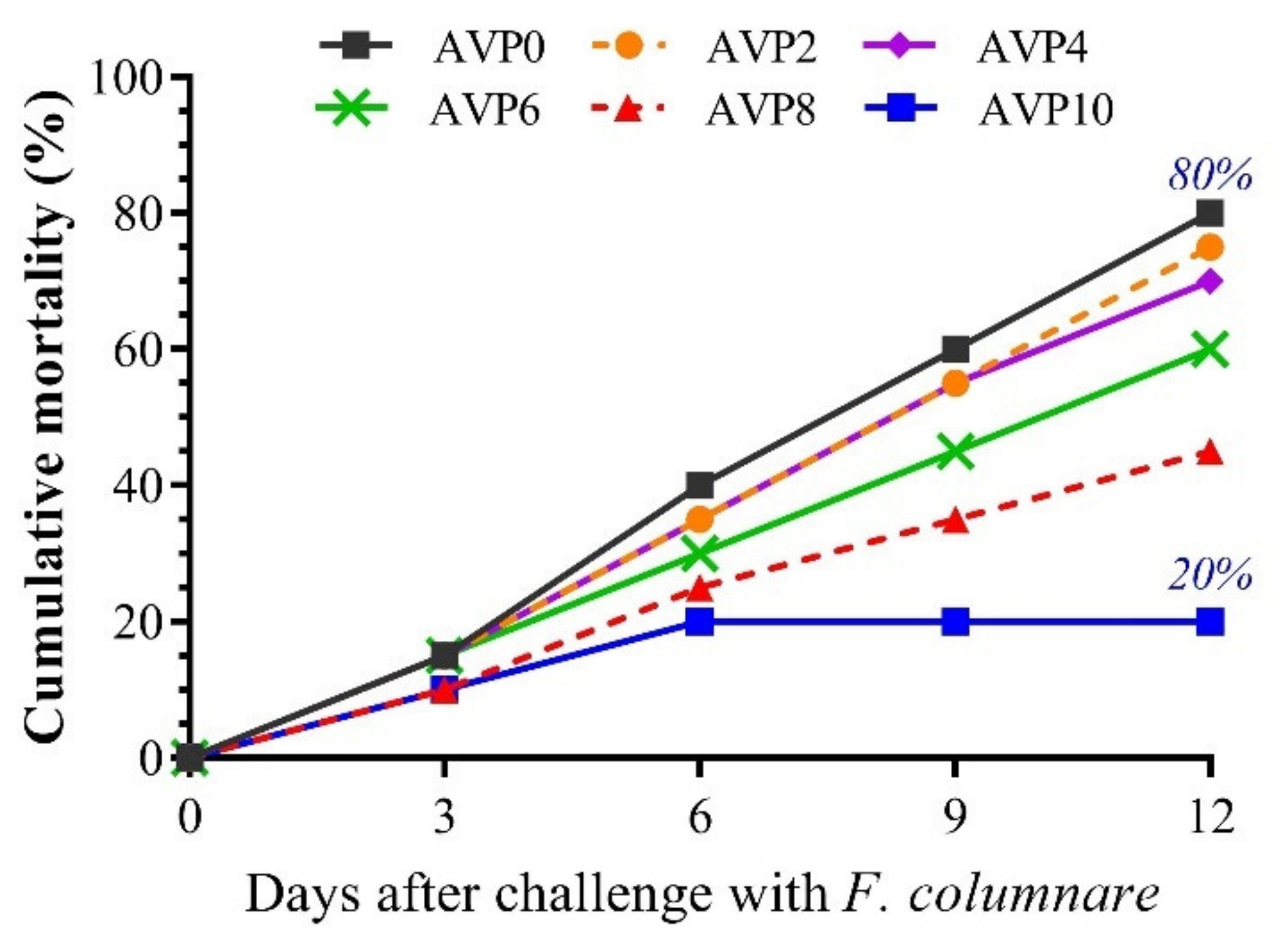
3.6. Fish Resistance against Challenge with Flavobacterium columnare
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tilami, S.K.; Sampels, S. Nutritional value of fish: Lipids, proteins, vitamins, and minerals. Rev. Fish. Sci. Aquac. 2018, 26, 243–253. [Google Scholar] [CrossRef]
- Mansour, A.T.; Ashour, M.; Alprol, A.E.; Alsaqufi, A.S. Aquatic Plants and Aquatic Animals in the Context of Sustainability: Cultivation Techniques, Integration, and Blue Revolution. Sustainability 2022, 14, 3257. [Google Scholar] [CrossRef]
- El-Sayed, A.-F.M. Tilapia Culture, 2nd ed.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Tartor, Y.; Taha, M.; Mahboub, H.; El Ghamery, M. Yeast species associated with diseased fish: Occurrence, identification, experimental challenges and antifungal susceptibility testing. Aquaculture 2018, 488, 134–144. [Google Scholar] [CrossRef]
- Rahman, A.N.A.; Mansour, D.A.; Abd El-Rahman, G.I.; Elseddawy, N.M.; Zaglool, A.W.; Khamis, T.; Mahmoud, S.F.; Mahboub, H.H. Imidacloprid toxicity in Clarias gariepinus: Protective role of dietary Hyphaene thebaica against biochemical and histopathological disruption, oxidative stress, immune genes expressions, and Aeromonas sobria infection. Aquaculture 2022, 555, 738170. [Google Scholar] [CrossRef]
- Mahboub, H.H.; Faggio, C.; Hendam, B.M.; Algharib, S.A.; Alkafafy, M.; Hashem, M.A.; Mahmoud, Y.K.; Khamis, T.; Abdel-Ghany, H.M.; Masoud, S.R. Immune-antioxidant trait, Aeromonas veronii resistance, growth, intestinal architecture, and splenic cytokines expression of Cyprinus carpio fed Prunus armeniaca kernel-enriched diets. Fish Shellfish Immunol. 2022, 124, 182–191. [Google Scholar] [CrossRef]
- Mahboub, H.H.; Elsheshtawy, H.M.; Sheraiba, N.I.; Fahmy, E.M.; Mohamed, E.A.; Abdelnaeim, N.S.; Mohamed, D.I.; Ismail, T.A.; Ahmed, S.A. Dietary black cumin (Nigella sativa) improved hemato-biochemical, oxidative stress, gene expression, and immunological response of Nile tilapia (Oreochromis niloticus) infected by Burkholderia cepacia. Aquacult. Rep. 2022, 22, 100943. [Google Scholar] [CrossRef]
- Mahboub, H.H.; Nada, H.S.; Abdel-Ghany, H.M.; Ghanem, R.; Ahmed Ismail, T.; Abdel Rahman, A.N. Detection, diagnosis, Koch’s postulate, hepatorenal and antioxidant indicators for some systemic pathogenic fungi invading the liver and kidneys of African catfish (Clarias gariepinus) in Egypt with a histopathological approach. Aquacult. Res. 2022, 53, 2670–2685. [Google Scholar] [CrossRef]
- El-Bouhy, Z.M.; Reda, R.M.; Mahboub, H.H.; Gomaa, F.N. Bioremediation effect of pomegranate peel on subchronic mercury immunotoxicity on African catfish (Clarias gariepinus). Environ. Sci. Pollut. Res. 2021, 28, 2219–2235. [Google Scholar] [CrossRef]
- Rashidian, G.; Mahboub, H.H.; Hoseinifar, S.H.; Ghafarifarsani, H.; Zare, M.; Punyatong, M.; Van Doan, H. Allium hirtifolium protects Cyprinus carpio against the detrimental responses mediated by foodborne zinc oxide nanoparticle. Aquaculture 2022, 555, 738252. [Google Scholar] [CrossRef]
- Rashidian, G.; Mahboub, H.H.; Fahim, A.; Hefny, A.A.; Prokić, M.D.; Rainis, S.; Boldaji, J.T.; Faggio, C. Mooseer (Allium hirtifolium) boosts growth, general health status, and resistance of rainbow trout (Oncorhynchus mykiss) against Streptococcus iniae infection. Fish Shellfish Immunol. 2022, 120, 360–368. [Google Scholar] [CrossRef]
- Chen, J.; Chen, D.; Li, J.; Liu, Y.; Gu, X.; Teng, X. Cadmium-induced oxidative stress and immunosuppression mediated mitochondrial apoptosis via JNK-FoxO3a-PUMA pathway in common carp (Cyprinus carpio L.) gills. Aquat. Toxicol. 2021, 233, 105775. [Google Scholar] [CrossRef]
- Sun, Q.; Liu, Y.; Teng, X.; Luan, P.; Teng, X.; Yin, X. Immunosuppression participated in complement activation-mediated inflammatory injury caused by 4-octylphenol via TLR7/IκBα/NF-κB pathway in common carp (Cyprinus carpio) gills. Aquat. Toxicol. 2022, 249, 106211. [Google Scholar] [CrossRef]
- Al-osaj, S.L. Cytotoxic Evaluation of Alchemilla Vulgaris Favonoids in Normal Blood Lymphocytes; Al-Nahrain University: Baghdad, Iraq, 2016. [Google Scholar]
- Duckstein, S.M.; Lotter, E.M.; Meyer, U.; Lindequist, U.; Stintzing, F.C. Phenolic constituents from Alchemilla vulgaris L. and Alchemilla mollis (Buser) Rothm. at different dates of harvest. Z. Für Nat. C 2012, 67, 529–540. [Google Scholar] [CrossRef] [Green Version]
- El-Hadidy, E.M.; Refat, O.G.; Halaby, M.S.; Elmetwaly, E.M.; Omar, A.A. Effect of Lion’s Foot (Alchemilla vulgaris) on Liver and Renal Functions in Rats Induced by CCl4. Food Nutr. Sci. 2018, 9, 46–63. [Google Scholar]
- Maier, M.; Oelbermann, A.-L.; Renner, M.; Weidner, E. Screening of European medicinal herbs on their tannin content-New potential tanning agents for the leather industry. Ind. Crops Prod. 2017, 99, 19–26. [Google Scholar] [CrossRef]
- Özbilgin, S.; Özbek, H.; Kirmizi, N.İ.; Öz, B.E.; Kurtul, E.; Özrenk, B.C.; Işcan, G.S.; Acikara, Ö.B. Evaluation of the antidiabetic activity of Alchemilla persica Rothm. In mice with diabetes induced by alloxan. Turk. J. Pharm. Sci. 2019, 16, 261. [Google Scholar] [CrossRef]
- Jarić, S.; Mačukanović-Jocić, M.; Djurdjević, L.; Mitrović, M.; Kostić, O.; Karadžić, B.; Pavlović, P. An ethnobotanical survey of traditionally used plants on Suva planina mountain (south-eastern Serbia). J. Ethnopharmacol. 2015, 175, 93–108. [Google Scholar] [CrossRef]
- Kuete, V. Potential of Cameroonian plants and derived products against microbial infections: A review. Planta Med. 2010, 76, 1479–1491. [Google Scholar] [CrossRef] [Green Version]
- Mazurkova, N.; Protsenko, M.; Filippova, E.; Kukushkina, T.; Vysochina, G.; Lobanova, I.; Mazurkov, O.Y.; Shishkina, L.; Agafonov, A. Investigation of the antiviral activity of experimental samples obtained from the grass and roots of Alchemilla vulgaris L. against vaccinia virus and ectromelia virus. Drug Dev. Regist. 2019, 8, 9–15. [Google Scholar] [CrossRef]
- Boroja, T.; Mihailović, V.; Katanić, J.; Pan, S.-P.; Nikles, S.; Imbimbo, P.; Monti, D.; Stanković, N.; Stanković, M.; Bauer, R. The biological activities of roots and aerial parts of Alchemilla vulgaris L. S. Afr. J. Bot. 2018, 116, 175–184. [Google Scholar] [CrossRef]
- Figueiredo, H.; Klesius, P.; Arias, C.; Evans, J.; Shoemaker, C.; Pereira, D., Jr.; Peixoto, M. Isolation and characterization of strains of Flavobacterium columnare from Brazil. J. Fish. Dis. 2005, 28, 199–204. [Google Scholar] [CrossRef]
- Dong, H.; Senapin, S.; LaFrentz, B.; Rodkhum, C. Virulence assay of rhizoid and non-rhizoid morphotypes of Flavobacterium columnare in red tilapia, Oreochromis sp., fry. J. Fish. Dis. 2016, 39, 649–655. [Google Scholar] [CrossRef]
- Groff, J.M.; Lapatra, S.E. Infectious diseases impacting the commercial culture of salmonids. J. Appl. Aquacult. 2000, 10, 17–90. [Google Scholar] [CrossRef]
- Elgendy, M.Y.; Abdelsalam, M.; Mohamed, S.A.; Ali, S.E. Molecular characterization, virulence profiling, antibiotic susceptibility, and scanning electron microscopy of Flavobacterium columnare isolates retrieved from Nile tilapia (Oreochromis niloticus). Aquacult. Int. 2022, 30, 845–862. [Google Scholar] [CrossRef]
- El-Houseiny, W.; Khalil, A.A.; Abd-Elhakim, Y.M.; Badr, H.A. The potential role of turmeric and black pepper powder diet supplements in reversing cadmium-induced growth retardation, ATP depletion, hepatorenal damage, and testicular toxicity in Clarias gariepinus. Aquaculture 2019, 510, 109–121. [Google Scholar] [CrossRef]
- Khalil, A.A.; Abd-Elhakim, Y.M.; Said, E.N.; Moselhy, A.A.; Abu-Elsaoud, A.M.; El-Houseiny, W. Milk thistle and co-enzyme Q10 fortified diets lessen the nickel chloride-induced neurotoxic and neurobehavioral impairments in Oreochromis niloticus via regulating the oxidative stress response, acetylcholinesterase activity, and brain nickel content. Aquaculture 2022, 553, 738102. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analysis of Chemist: Washington, DC, USA, 2000. [Google Scholar]
- Feldman, B.V.; Zinkl, J.G.; Jain, N.C.; Schalm, O.W. Schalm’s Veterinary Hematology, 5th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2000. [Google Scholar]
- Bain, B.J.; Bates, I.; Laffan, M.A. Dacie and Lewis Practical Haematology E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Burtis, C.A.; Ashwood, E.R. Tietz Textbook of Clinical Chemistry; American Assosiation for Clinical Chemistry: Washington, DC, USA, 1994. [Google Scholar]
- Wenger, C.; Kaplan, A.; Rubaltelli, F.; Hammerman, C. Alkaline phosphatase. Clin. Chem. 1984, 62, 1094–1098. [Google Scholar]
- Murray, R.; Kaplan, A. Aspartate Aminotransferase; CV Mosby Company: Maryland Heights, MO, USA, 1984; pp. 1105–1108. [Google Scholar]
- Neely, W.; Phillipson, J. Automated enzymatic method for determining ammonia in plasma with 14 day stability. Clin. Chem. 1988, 34, 1868–1869. [Google Scholar] [CrossRef]
- Martinen, K. Improved micro-method for determination of serum bilirubin. Clin. Chim. Acta 1966, 13, 161–170. [Google Scholar] [CrossRef]
- Rashidian, G.; Shahin, K.; Elshopakey, G.E.; Mahboub, H.H.; Fahim, A.; Elabd, H.; Prokić, M.D.; Faggio, C. The Dietary Effects of Nutmeg (Myristica fragrans) Extract on Growth, Hematological Parameters, Immunity, Antioxidant Status, and Disease Resistance of Common Carp (Cyprinus carpio) against Aeromonas hydrophila. J. Mar. Sci. Eng. 2022, 10, 325. [Google Scholar] [CrossRef]
- Ellis, A.E. Lysozyme assays. Tech. Fish Immunol. 1990, 1, 101–103. [Google Scholar]
- Montgomery, H.; Dymock, J.F. The determination of nitrite in water. Analyst 1961, 86, 414–416. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzymology. 1984, 105, 121–126. [Google Scholar]
- Nishikimi, M.; Rao, N.A.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Beutler, E. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar]
- Uchiyama, M.; Mihara, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef]
- Kumari, J.; Sahoo, P. Effects of cyclophosphamide on the immune system and disease resistance of Asian catfish Clarias batrachus. Fish Shellfish Immunol. 2005, 19, 307–316. [Google Scholar] [CrossRef]
- Griffin, B. A simple procedure for identification of Cytophaga columnaris. J. Aquat. Anim. Health 1992, 4, 63–66. [Google Scholar] [CrossRef]
- Farmer, B.D. Improved Methods for the Isolation and Characterization of Flavobacterium Columnare; Louisiana State University: Baton Rouge, LA, USA, 2004. [Google Scholar]
- García-Beltrán, J.M.; Mansour, A.T.; Alsaqufi, A.S.; Ali, H.M.; Esteban, M.Á. Effects of aqueous and ethanolic leaf extracts from drumstick tree (Moringa oleifera) on gilthead seabream (Sparus aurata L.) leucocytes, and their cytotoxic, antitumor, bactericidal and antioxidant activities. Fish Shellfish Immunol. 2020, 106, 44–55. [Google Scholar] [CrossRef]
- Köseman, A.; Akdemir, F.; Üremiş, N.; Şeker, İ.; Türköz, Y. Effects of Alchemilla vulgaris on growth performance, carcass characteristics and some biochemical parameters of heat stressed broilers. J. Hell. Vet. Med. Soc. 2020, 71, 2491–2498. [Google Scholar] [CrossRef]
- Akdemir, F.; Köseman, A.; Şeker, I. Alchemilla vulgaris effects on egg production and quality expressed by heatstressed quail during the late laying period. S. Afr. J. Anim. Sci. 2019, 49, 857–868. [Google Scholar] [CrossRef]
- Samah, S.; Abdullah, K.; Ream, N. Phytochemical screening of Alchemilla vulgaris, Sophora Japonica, Crataegus azarolus, and their inhibitory activity on lipase and α-amylase. Int. J. Appl. Sci. Res. 2018, 6, 1–21. [Google Scholar]
- Van Doan, H.; Hoseinifar, S.H.; Sringarm, K.; Jaturasitha, S.; Khamlor, T.; Dawood, M.A.; Esteban, M.Á.; Soltani, M.; Musthafa, M.S. Effects of elephant’s foot (Elephantopus scaber) extract on growth performance, immune response, and disease resistance of nile tilapia (Oreochromis niloticus) fingerlings. Fish Shellfish Immunol. 2019, 93, 328–335. [Google Scholar] [CrossRef]
- Peng, K.; Wang, G.; Zhao, H.; Wang, Y.; Mo, W.; Wu, H.; Huang, Y. Effect of high level of carbohydrate and supplementation of condensed tannins on growth performance, serum metabolites, antioxidant and immune response, and hepatic glycometabolism gene expression of Lateolabrax japonicus. Aquacult. Rep. 2020, 18, 100515. [Google Scholar] [CrossRef]
- Peng, K.; Zhao, H.; Wang, G.; Chen, B.; Mo, W.; Huang, Y. Effect of condensed tannins on growth performance, intestinal immune capacity and bacterial microbiomes of Lateolabrax japonicus. Aquacult. Res. 2021, 52, 5321–5331. [Google Scholar] [CrossRef]
- Li, M.; Feng, L.; Jiang, W.-D.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Zhou, X.-Q. Condensed tannins decreased the growth performance and impaired intestinal immune function in on-growing grass carp (Ctenopharyngodon idella). Br. J. Nutr. 2020, 123, 737–755. [Google Scholar] [CrossRef]
- El-Houseiny, W.; Abd El-Hakim, Y.M.; Metwally, M.M.; Ghfar, S.S.A.; Khalil, A.A. The single or combined Silybum marianum and co-enzyme Q10 role in alleviating fluoride-induced impaired growth, immune suppression, oxidative stress, histological alterations, and reduced resistance to Aeromonas sobria in African catfish (Clarias gariepinus). Aquaculture 2022, 548, 737693. [Google Scholar]
- Mansour, A.T.; Hamed, H.S.; El-Beltagi, H.S.; Mohamed, W.F. Modulatory Effect of Papaya Extract against Chlorpyrifos-Induced Oxidative Stress, Immune Suppression, Endocrine Disruption, and DNA Damage in Female Clarias gariepinus. Int. J. Environ. Res. Public 2022, 19, 4640. [Google Scholar] [CrossRef]
- Sallam, A.E.; Mansour, A.T.; Srour, T.M.; Goda, A.M.A. Effects of different carotenoid supplementation sources with or without sodium taurocholate on growth, feed utilization, carotenoid content and antioxidant status in fry of the European seabass, Dicentrarchus Labrax. Aquacult. Res. 2017, 48, 3848–3858. [Google Scholar] [CrossRef]
- Köseman, A.; Akdemir, F.; Üremiş, N.; Şeker, İ.; Türköz, Y. Effects of Alchemilla vulgaris on haematology and antioxidant status of heatstressed quails during the late laying period. J. Hell. Vet. Med. Soc. 2021, 72, 3463–3472. [Google Scholar] [CrossRef]
- Motamedi-Tehrani, J.; Ebrahimi-Dorcheh, E.; Malekpouri, P.; Goli, S. Liver alteration and hematological and serum biochemical responses of common carp, Cyprinus carpio Linnaeus, 1758, following long-term feeding of pistachio (Pistacia vera) green hull extract as a source of natural phenol. J. Appl. Ichthyol. 2016, 32, 906–912. [Google Scholar] [CrossRef]
- Ozbek, H.; Acikara, O.B.; Keskin, I.; Kirmizi, N.I.; Ozbilgin, S.; Oz, B.E.; Kurtul, E.; Ozrenk, B.C.; Tekin, M.; Saltan, G. Evaluation of hepatoprotective and antidiabetic activity of Alchemilla mollis. Biomed. Pharmacother. 2017, 86, 172–176. [Google Scholar] [CrossRef]
- Li, M.; Zhu, X.; Tian, J.; Liu, M.; Wang, G. Dietary flavonoids from Allium mongolicum Regel promotes growth, improves immune, antioxidant status, immune-related signaling molecules and disease resistance in juvenile northern snakehead fish (Channa argus). Aquaculture 2019, 501, 473–481. [Google Scholar] [CrossRef]
- Emeish, W.F.; Al-Amgad, Z.; Ahmed, H. Antioxidant, immunostimulant and renal protective activities of tri-herbal combination in African Sharptooth Catfish, Clarias gariepinus. J. Vet. Med. Res. 2018, 25, 213–229. [Google Scholar] [CrossRef]
- Zhao, L.; Yuan, B.-D.; Zhao, J.-L.; Jiang, N.; Zhang, A.-Z.; Wang, G.-Q.; Li, M.-Y. Amelioration of hexavalent chromium-induced bioaccumulation, oxidative stress, tight junction proteins and immune-related signaling factors by Allium mongolicum Regel flavonoids in Ctenopharyngodon idella. Fish Shellfish Immunol. 2020, 106, 993–1003. [Google Scholar] [CrossRef]
- Shilova, I.; Suslov, N.; Samylina, I.; Baeva, V.; Lazareva, N.; Mazin, E. Neuroprotective properties of common lady’s mantle infusion. Pharm. Chem. J. 2020, 53, 1059–1062. [Google Scholar] [CrossRef]
- Vlaisavljević, S.; Jelača, S.; Zengin, G.; Mimica-Dukić, N.; Berežni, S.; Miljić, M.; Stevanović, Z.D. Alchemilla vulgaris agg. (Lady’s mantle) from central Balkan: Antioxidant, anticancer and enzyme inhibition properties. RSC Adv. 2019, 9, 37474–37483. [Google Scholar] [CrossRef] [Green Version]
- Tadić, V.M.; Krgović, N.; Žugić, A. Lady’s mantle (Alchemilla vulgaris L., Rosaceae): A review of traditional uses, phytochemical profile, and biological properties. Lek. Sirovine 2020, 40, 66–74. [Google Scholar] [CrossRef]
- Jain, S.; Yadav, A.; Gothalwal, R. Assessment of total phenolic, flavonoid content and in vitro antioxidant properties of Alchemillia vulgaris (lady’s mantle). J. Adv. Sci. Res. 2021, 12, 205–209. [Google Scholar] [CrossRef]
- Lizárraga-Velázquez, C.E.; Hernández, C.; González-Aguilar, G.A.; Heredia, J.B. Effect of dietary intake of phenolic compounds from mango peel extract on growth, lipid peroxidation and antioxidant enzyme activities in zebrafish (Danio rerio). Lat. Am. J. Aquat. Res. 2019, 47, 602–611. [Google Scholar] [CrossRef] [Green Version]
- Mansour, A.T.; Espinosa, C.; García-Beltrán, J.M.; Miao, L.; Francisco, D.C.C.; Alsaqufi, A.S.; Esteban, M.Á. Dietary supplementation of drumstick tree, Moringa oleifera, improves mucosal immune response in skin and gills of seabream, Sparus aurata, and attenuates the effect of hydrogen peroxide exposure. Fish Physiol. Bioch. 2020, 46, 981–996. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.T.; El-Feky, M.M.; El-Beltagi, H.S.; Sallam, A.E. Synergism of Dietary Co-Supplementation with Lutein and Bile Salts Improved the Growth Performance, Carotenoid Content, Antioxidant Capacity, Lipid Metabolism, and Lipase Activity of the Marbled Spinefoot Rabbitfish, Siganus rivulatus. Animals 2020, 10, 1643. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.T.; Miao, L.; Espinosa, C.; García-Beltrán, J.M.; Francisco, D.C.C.; Esteban, M.Á. Effects of dietary inclusion of Moringa oleifera leaves on growth and some systemic and mucosal immune parameters of seabream. Fish Physiol. Bioch. 2018, 44, 1223–1240. [Google Scholar] [CrossRef] [PubMed]
- Kiselova, Y.; Ivanova, D.; Chervenkov, T.; Gerova, D.; Galunska, B.; Yankova, T. Correlation between the in vitro antioxidant activity and polyphenol content of aqueous extracts from Bulgarian herbs. Phytother. Res. 2006, 4, 961–965. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Nada, H.S.; Elsheshtawy, H.M.; Ibrahim, S.M.; Fahmy, E.M.; Khedr, M.H.; Moustafa, S.M.; Ismail, T.A.; Gesriha, S.; Assayed, M.E. Comparative antitoxic potency of honey and natamycin-supplemented diets against aflatoxicosis and their influences on growth, serum biochemistry, immunohistochemistry, and residual deposition in Nile tilapia (Oreochromis niloticus). Aquaculture 2022, 551, 737934. [Google Scholar] [CrossRef]
- Mansour, A.T.; Esteban, M.Á. Effects of carbon sources and plant protein levels in a biofloc system on growth performance, and the immune and antioxidant status of Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2017, 64, 202–209. [Google Scholar] [CrossRef]
- Kostadinović, L.; Popović, S.; Pelić, D.L.; Čabarkapa, I.; Đuragić, O.; Lević, J. Medicinal plants as natural alternative to coccidial synthetic drugs in broiler chicken production. J. Agron. 2019, 5, 325–334. [Google Scholar]
- El Mesallamy, A.M.; El-Marakby, H.I.; Souleman, A.M.; Abd El-Naby, F.S. Evaluation of phenolic extract of licorice roots in diets of Nile tilapia (Oreochromis niloticus). Egypt. Pharm. J. 2015, 14, 117. [Google Scholar]
- Secombes, C.; Wang, T. The Innate and Adaptive Immune System of Fish. In Infectious Disease in Aquaculture; Elsevier: Amsterdam, The Netherlands, 2012; pp. 3–68. [Google Scholar]
- Tie, H.-M.; Feng, L.; Jiang, W.-D.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Zhou, X.-Q. Dietary exogenous supplementation of nucleotides strengthens the disease resistance, antioxidant capacity and immunity in the gill of on-growing grass carp (Ctenopharyngodon idella) following a challenge with Flavobacterium columnare. Aquaculture 2021, 540, 736729. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Devi, G.; Van Doan, H.; Balasundaram, C.; Esteban, M.Á.; Abdel-Tawwab, M. Impact of grape pomace flour (GPF) on immunity and immune-antioxidant-anti-inflammatory genes expression in Labeo rohita against Flavobacterium columnaris. Fish Shellfish Immunol. 2021, 111, 69–82. [Google Scholar] [CrossRef]
- Tasić, S. Ethnobotany in SEE-WB countries; traditional uses of indigenous plants. Lek. Sirovine 2012, 32, 71–81. [Google Scholar]
- Choi, J.; Park, Y.-G.; Yun, M.-S.; Seol, J.-W. Effect of herbal mixture composed of Alchemilla vulgaris and Mimosa on wound healing process. Biomed. Pharmacother. 2018, 106, 326–332. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/kg) | Dietary Alchemilla vulgaris Levels (g/kg Diet) | |||||
|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | |
| Fishmeal | 110 | 110 | 110 | 110 | 110 | 110 |
| Corn flour | 330 | 330 | 330 | 330 | 330 | 330 |
| Soybean meal 44% | 290 | 290 | 290 | 290 | 290 | 290 |
| Corn gluten meal 60% | 120 | 120 | 120 | 120 | 120 | 120 |
| Wheat bran | 80 | 78 | 76 | 74 | 72 | 70 |
| Soybean oil | 20 | 20 | 20 | 20 | 20 | 20 |
| Fish oil | 20 | 20 | 20 | 20 | 20 | 20 |
| Alchemilla vulgaris powder (AVP) | 0 | 2 | 4 | 6 | 8 | 10 |
| Vitamin premix 1 | 15 | 15 | 15 | 15 | 15 | 15 |
| Mineral premix 2 | 15 | 15 | 15 | 15 | 15 | 15 |
| Total | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 |
| Chemical analysis (g/kg) | ||||||
| Crude protein (n × 6.25) | 308.8 | 309.5 | 309.7 | 310.0 | 310.3 | 310.6 |
| Crude lipids | 74.9 | 75.1 | 75.7 | 76.5 | 78.4 | 79.1 |
| Crude fiber | 52.3 | 53.4 | 53.8 | 54.1 | 54.5 | 54.9 |
| Ash | 52.2 | 53.7 | 54.5 | 56.9 | 57.9 | 58.4 |
| Nitrogen-free extract 3 | 511.8 | 508.3 | 506.3 | 502.5 | 498.9 | 497 |
| Gross energy (kcal/kg) 4 | 4556 | 4547 | 4546 | 4539 | 4544 | 4545 |
| Items | Dietary Alchemilla vulgaris Levels (g/kg Diet) | |||||
|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | |
| Final body weight (g) | 55.63 d ± 0.46 | 55.60 d ± 0.56 | 55.50 d ± 0.55 | 59.16 c ± 0.44 | 63.70 b ± 0.70 | 67.0 a ± 0.57 |
| Weight gain (g) | 32.96 d ± 0.31 | 32.20 d ± 0.23 | 32.76 d ± 40.0 | 36.26 c ± 0.20 | 40.26 b ± 0.37 | 44.13 a ± 0.20 |
| Daily weight gain (g) | 0.54 d ± 0.005 | 0.53 d ± 0.003 | 0.54 d ± 0.006 | 0.60 c ± 0.003 | 0.67 b ± 0.006 | 0.73 a ± 0.002 |
| Specific growth rate (%) | 0.65 c ± 0.018 | 0.62 c ± 0.007 | 0.64 c ± 0.017 | 0.68 bc ± 0.014 | 0.72 ab ± 0.003 | 0.77 a ± 0.015 |
| Feed intake (g) | 52.00 c ± 0.57 | 51.00 c ± 0.57 | 51.33 c ± 0.44 | 54.50 b ± 0.28 | 56.00 ab ± 0.28 | 58.00 a ± 0.57 |
| Feed conversion ratio | 1.57 a ± 0.012 | 1.58 a ± 0.006 | 1.56 a ± 0.007 | 1.50 b ± 0.007 | 1.39 c ± 0.008 | 1.31 d ± 0.010 |
| Condition factor (K) | 2.02 c ± 0.008 | 2.04 c ± 0.017 | 2.05 c ± 0.020 | 2.065 c ± 0.010 | 2.22 b ± 0.004 | 2.38 a ± 0.026 |
| Items | Dietary Alchemilla vulgaris Levels (g/kg Diet) | |||||
|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | |
| RBCs (106/mm3) | 3.78 d ± 0.008 | 3.81 cd ± 0.008 | 3.84 c ± 0.008 | 3.89 b ± 0.008 | 3.93 ab ± 0.005 | 3.96 a ± 0.017 |
| Hb (gm/dL) | 11.20 d ± 0.05 | 11.30 cd ± 0.02 | 11.34 cd ± 0.01 | 11.44 bc ± 0.02 | 11.52 b ± 0.01 | 11.80 a ± 0.04 |
| PCV (%) | 33.60 d ± 0.17 | 33.92 cd ± 0.08 | 34.03 cd ± 0.04 | 34.33 bc ± 0.07 | 34.57 b ± 0.04 | 35.42 a ± 0.14 |
| MCV(fl) | 88.73 ab ± 0.66 | 88.87 ab ± 0.13 | 88.46 ab ± 0.10 | 88.10 b ± 0.05 | 87.96 b ± 0.02 | 89.44 a ± 0.06 |
| MCH (%) | 29.57 ab ± 0.22 | 29.62 ab ± 0.04 | 29.48 ab ± 0.03 | 29.36 b ± 0.01 | 29.32 b ± 0.007 | 29.81 a ± 0.02 |
| WBCs (103/mm3) | 5.25 d ± 0.02 | 5.28 d ± 0.01 | 5.32 cd ± 0.01 | 5.37 bc ± 0.008 | 5.42 b ± 0.01 | 5.51 a ± 0.02 |
| Lymphocytes (103/mm3) | 2.90 c ± 0.005 | 2.90 c ± 0.008 | 2.90 c ± 0.005 | 2.92 bc ± 0.005 | 2.94 b ± 0.008 | 2.99 a ± 0.011 |
| Neutrophils (103/mm3) | 1.38 c ± 0.006 | 1.40 bc ± 0.005 | 1.41 bc ± 0.008 | 1.43 ab ± 0.012 | 1.43 ab ± 0.005 | 1.46 a ± 0.008 |
| Eosinophils (103/mm3) | 0.33 b ± 0.005 | 0.33 b ± 0.005 | 0.35 ab ± 0.008 | 0.38 a ± 0.005 | 0.38 a ± 0.005 | 0.37 a ± 0.005 |
| Monocytes (103/mm3) | 0.64 b ± 0.005 | 0.65 b ± 0.005 | 0.65 b ± 0.011 | 0.64 b ± 0.005 | 0.67 ab ± 0.005 | 0.69 a ± 0.005 |
| Items | Dietary Alchemilla vulgaris Levels (g/kg Diet) | |||||
|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | |
| Total proteins (g/dL) | 5.46 d ± 0.10 | 6.10 c ± 0.069 | 6.13 bc ± 0.04 | 6.44 b ± 0.086 | 6.81 a ± 0.060 | 6.94 a ± 0.026 |
| Albumin (g/dL) | 2.31 c ± 0.101 | 2.92 b ± 0.136 | 3.03 ab ± 0.02 | 3.12 ab ± 0.06 | 3.26 ab ± 0.02 | 3.31 a ± 0.014 |
| ALT (U/L) | 13.24 a ± 0.43 | 13.26 a ± 0.48 | 12.45 ab ± 0.29 | 11.96 ab ± 0.12 | 11.73 b ± 0.06 | 11.58 b ± 0.04 |
| AST (U/L) | 27.80 a ± 0.32 | 27.70 a ± 0.32 | 27.02 ab ± 0.07 | 26.85 ab ± 0.13 | 26.71 b ± 0.12 | 26.15 b ± 0.09 |
| ALP (IU/L) | 24.30 a ± 0.15 | 24.21 a ± 0.06 | 24.08 a ± 0.06 | 23.90 ab ± 0.09 | 23.60 b ± 0.08 | 23.09 c ± 0.06 |
| Urea (mg/dL) | 2.76 a ± 0.037 | 2.76 a ± 0.023 | 2.68 a ± 0.02 | 2.59 a ± 0.03 | 2.18 b ± 0.044 | 2.14 b ± 0.049 |
| Creatinine (mg/dL) | 0.436 a ± 0.01 | 0.44 a ± 0.005 | 0.41 ab ± 0.01 | 0.356 bc ± 0.01 | 0.323 c ± 0.01 | 0.246 d ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mansour, A.T.; Mahboub, H.H.; Elshopakey, G.E.; Aziz, E.K.; Alhajji, A.H.M.; Rayan, G.; Ghazzawy, H.S.; El-Houseiny, W. Physiological Performance, Antioxidant and Immune Status, Columnaris Resistance, and Growth of Nile Tilapia That Received Alchemilla vulgaris-Supplemented Diets. Antioxidants 2022, 11, 1494. https://doi.org/10.3390/antiox11081494
Mansour AT, Mahboub HH, Elshopakey GE, Aziz EK, Alhajji AHM, Rayan G, Ghazzawy HS, El-Houseiny W. Physiological Performance, Antioxidant and Immune Status, Columnaris Resistance, and Growth of Nile Tilapia That Received Alchemilla vulgaris-Supplemented Diets. Antioxidants. 2022; 11(8):1494. https://doi.org/10.3390/antiox11081494
Chicago/Turabian StyleMansour, Abdallah Tageldein, Heba H. Mahboub, Gehad E. Elshopakey, Enas K. Aziz, Adnan H. M. Alhajji, Gamal Rayan, Hesham S. Ghazzawy, and Walaa El-Houseiny. 2022. "Physiological Performance, Antioxidant and Immune Status, Columnaris Resistance, and Growth of Nile Tilapia That Received Alchemilla vulgaris-Supplemented Diets" Antioxidants 11, no. 8: 1494. https://doi.org/10.3390/antiox11081494
APA StyleMansour, A. T., Mahboub, H. H., Elshopakey, G. E., Aziz, E. K., Alhajji, A. H. M., Rayan, G., Ghazzawy, H. S., & El-Houseiny, W. (2022). Physiological Performance, Antioxidant and Immune Status, Columnaris Resistance, and Growth of Nile Tilapia That Received Alchemilla vulgaris-Supplemented Diets. Antioxidants, 11(8), 1494. https://doi.org/10.3390/antiox11081494