
N-Acetylcysteine Regenerates In Vivo Mercaptoalbumin

 , , ,
, , ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Methods
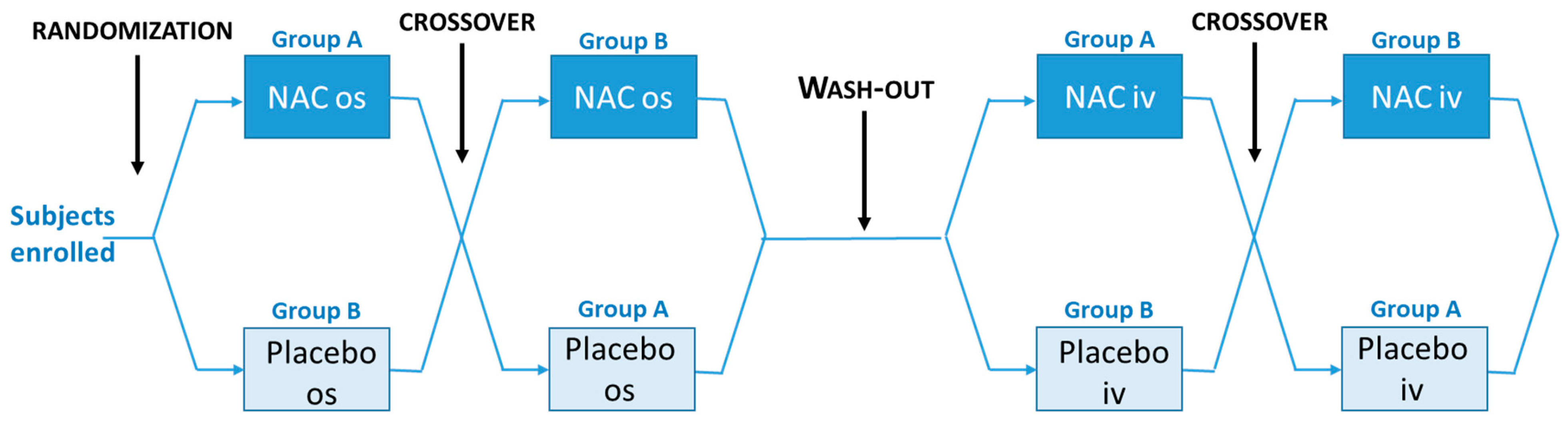
2.1. Study Design
2.2. Plasma Preparation
2.3. Albumin Analysis by Mass Spectrometry
2.4. Plasma Antioxidant Capacity Analysis (TRAP Assay)
2.5. Measurement of Glutathione
2.6. Statistical Analysis
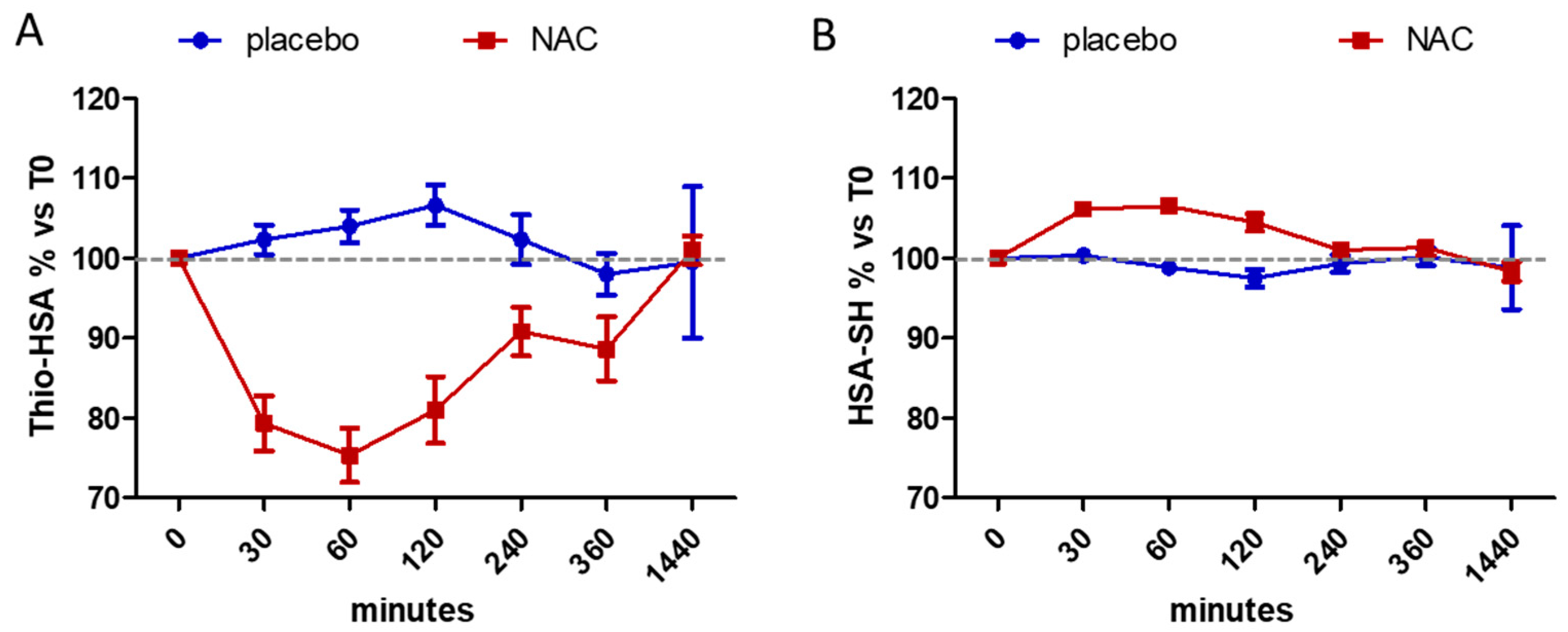
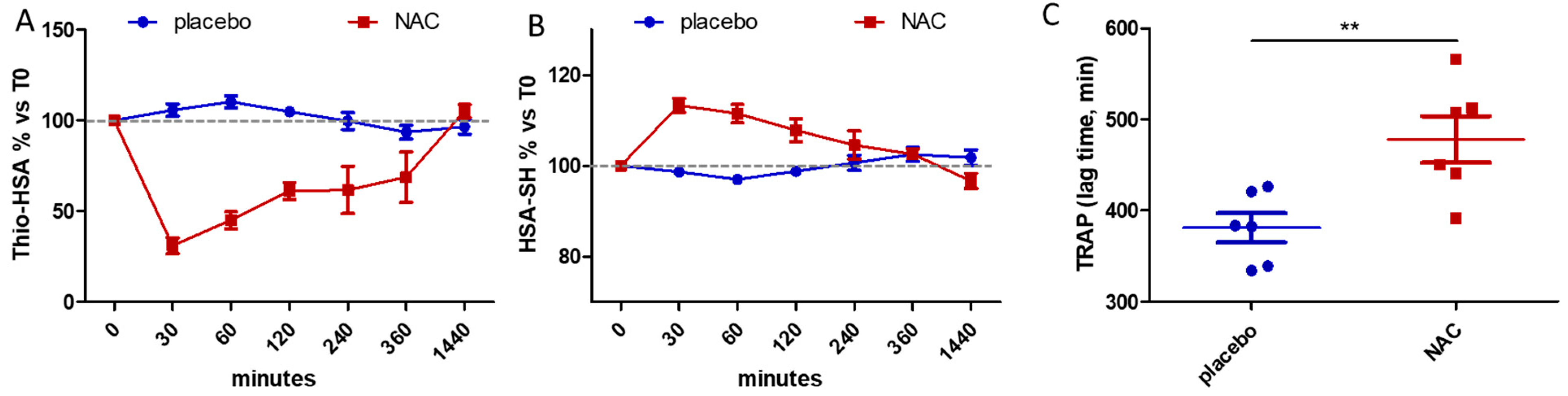
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sugio, S.; Kashima, A.; Mochizuki, S.; Noda, M.; Kobayashi, K. Crystal structure of human serum albumin at 2.5 A resolution. Protein Eng. 1999, 12, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Bonanata, J.; Turell, L.; Antmann, L.; Ferrer-Sueta, G.; Botasini, S.; Mendez, E.; Alvarez, B.; Coitino, E.L. The thiol of human serum albumin: Acidity, microenvironment and mechanistic insights on its oxidation to sulfenic acid. Free Radic. Biol. Med. 2017, 108, 952–962. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, F.; Shibata, T.; Uchida, K. A unique mechanism for thiolation of serum albumins by disulphide molecules. J. Biochem. 2020, 167, 165–171. [Google Scholar] [CrossRef]
- Zoanni, B.; Brioschi, M.; Mallia, A.; Gianazza, E.; Eligini, S.; Carini, M.; Aldini, G.; Banfi, C. Novel insights about albumin in cardiovascular diseases: Focus on heart failure. Mass Spectrom. Rev. 2021, e21743. [Google Scholar] [CrossRef] [PubMed]
- Brioschi, M.; Gianazza, E.; Mallia, A.; Zoanni, B.; Altomare, A.; Martinez Fernandez, A.; Agostoni, P.; Aldini, G.; Banfi, C. S-Thiolation Targets Albumin in Heart Failure. Antioxidants 2020, 9, 763. [Google Scholar] [CrossRef]
- Eligini, S.; Porro, B.; Aldini, G.; Colli, S.; Banfi, C. N-Acetylcysteine Inhibits Platelet Function through the Regeneration of the Non-Oxidative Form of Albumin. Antioxidants 2022, 11, 445. [Google Scholar] [CrossRef]
- Altomare, A.; Baron, G.; Brioschi, M.; Longoni, M.; Butti, R.; Valvassori, E.; Tremoli, E.; Carini, M.; Agostoni, P.; Vistoli, G.; et al. N-Acetyl-Cysteine Regenerates Albumin Cys34 by a Thiol-Disulfide Breaking Mechanism: An Explanation of Its Extracellular Antioxidant Activity. Antioxidants 2020, 9, 367. [Google Scholar] [CrossRef] [PubMed]
- Calzetta, L.; Matera, M.G.; Rogliani, P.; Cazzola, M. Multifaceted activity of N-acetyl-l-cysteine in chronic obstructive pulmonary disease. Expert Rev. Respir. Med. 2018, 12, 693–708. [Google Scholar] [CrossRef]
- Prescott, L.F.; Illingworth, R.N.; Critchley, J.A.; Stewart, M.J.; Adam, R.D.; Proudfoot, A.T. Intravenous N-acetylcystine: The treatment of choice for paracetamol poisoning. Br. Med. J. 1979, 2, 1097–1100. [Google Scholar] [CrossRef]
- Raghu, G.; Berk, M.; Campochiaro, P.A.; Jaeschke, H.; Marenzi, G.; Richeldi, L.; Wen, F.Q.; Nicoletti, F.; Calverley, P.M.A. The Multifaceted Therapeutic Role of N-Acetylcysteine (NAC) in Disorders Characterized by Oxidative Stress. Curr. Neuropharmacol. 2021, 19, 1202–1224. [Google Scholar] [CrossRef]
- Quintavalle, C.; Donnarumma, E.; Fiore, D.; Briguori, C.; Condorelli, G. Therapeutic strategies to prevent contrast-induced acute kidney injury. Curr. Opin. Cardiol. 2013, 28, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.H.; Xu, C.Y.; Fan, G.H. Efficacy of N-acetylcysteine in preventing atrial fibrillation after cardiac surgery: A meta-analysis of published randomized controlled trials. BMC Cardiovasc. Disord. 2014, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Deepmala; Slattery, J.; Kumar, N.; Delhey, L.; Berk, M.; Dean, O.; Spielholz, C.; Frye, R. Clinical trials of N-acetylcysteine in psychiatry and neurology: A systematic review. Neurosci. Biobehav. Rev. 2015, 55, 294–321. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I. Glutathione homeostasis and functions: Potential targets for medical interventions. J. Amino Acids 2012, 2012, 736837. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N-Acetylcysteine as an antioxidant and disulphide breaking agent: The reasons why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar] [CrossRef]
- Griendling, K.K.; Camargo, L.L.; Rios, F.J.; Alves-Lopes, R.; Montezano, A.C.; Touyz, R.M. Oxidative Stress and Hypertension. Circ. Res. 2021, 128, 993–1020. [Google Scholar] [CrossRef]
- Aldini, G.; Yeum, K.J.; Russell, R.M.; Krinsky, N.I. A method to measure the oxidizability of both the aqueous and lipid compartments of plasma. Free Radic. Biol. Med. 2001, 31, 1043–1050. [Google Scholar] [CrossRef]
- Squellerio, I.; Caruso, D.; Porro, B.; Veglia, F.; Tremoli, E.; Cavalca, V. Direct glutathione quantification in human blood by LC-MS/MS: Comparison with HPLC with electrochemical detection. J. Pharm. Biomed. Anal. 2012, 71, 111–118. [Google Scholar] [CrossRef]
- Soldini, D.; Zwahlen, H.; Gabutti, L.; Marzo, A.; Marone, C. Pharmacokinetics of N-acetylcysteine following repeated intravenous infusion in haemodialysed patients. Eur. J. Clin. Pharmacol. 2005, 60, 859–864. [Google Scholar] [CrossRef]
- Anders, M.W.; Dekant, W. Aminoacylases. Adv. Pharmacol. 1994, 27, 431–448. [Google Scholar] [CrossRef]
- Arfsten, D.; Johnson, E.; Thitoff, A.; Jung, A.; Wilfong, E.; Lohrke, S.; Bausman, T.; Eggers, J.; Bobb, A. Impact of 30-day oral dosing with N-acetyl-L-cysteine on Sprague-Dawley rat physiology. Int. J. Toxicol. 2004, 23, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Zembron-Lacny, A.; Slowinska-Lisowska, M.; Szygula, Z.; Witkowski, K.; Szyszka, K. The comparison of antioxidant and hematological properties of N-acetylcysteine and alpha-lipoic acid in physically active males. Physiol. Res. 2009, 58, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; Vistoli, G.; Regazzoni, L.; Gamberoni, L.; Facino, R.M.; Yamaguchi, S.; Uchida, K.; Carini, M. Albumin is the main nucleophilic target of human plasma: A protective role against pro-atherogenic electrophilic reactive carbonyl species? Chem. Res. Toxicol. 2008, 21, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Taverna, M.; Marie, A.L.; Mira, J.P.; Guidet, B. Specific antioxidant properties of human serum albumin. Ann. Intensive Care 2013, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Turell, L.; Carballal, S.; Botti, H.; Radi, R.; Alvarez, B. Oxidation of the albumin thiol to sulfenic acid and its implications in the intravascular compartment. Braz. J. Med. Biol. Res. 2009, 42, 305–311. [Google Scholar] [CrossRef]
- Aldini, G.; Gamberoni, L.; Orioli, M.; Beretta, G.; Regazzoni, L.; Maffei Facino, R.; Carini, M. Mass spectrometric characterization of covalent modification of human serum albumin by 4-hydroxy-trans-2-nonenal. J. Mass Spectrom. 2006, 41, 1149–1161. [Google Scholar] [CrossRef]
- Negre-Salvayre, A.; Garoby-Salom, S.; Swiader, A.; Rouahi, M.; Pucelle, M.; Salvayre, R. Proatherogenic effects of 4-hydroxynonenal. Free Radic. Biol. Med. 2017, 111, 127–139. [Google Scholar] [CrossRef]
- Gargiulo, S.; Testa, G.; Gamba, P.; Staurenghi, E.; Poli, G.; Leonarduzzi, G. Oxysterols and 4-hydroxy-2-nonenal contribute to atherosclerotic plaque destabilization. Free Radic. Biol. Med. 2017, 111, 140–150. [Google Scholar] [CrossRef]
- Aldini, G.; Regazzoni, L.; Orioli, M.; Rimoldi, I.; Facino, R.M.; Carini, M. A tandem MS precursor-ion scan approach to identify variable covalent modification of albumin Cys34: A new tool for studying vascular carbonylation. J. Mass Spectrom. 2008, 43, 1470–1481. [Google Scholar] [CrossRef]
- Nagumo, K.; Tanaka, M.; Chuang, V.T.; Setoyama, H.; Watanabe, H.; Yamada, N.; Kubota, K.; Tanaka, M.; Matsushita, K.; Yoshida, A.; et al. Cys34-cysteinylated human serum albumin is a sensitive plasma marker in oxidative stress-related chronic diseases. PLoS ONE 2014, 9, e85216. [Google Scholar] [CrossRef]
- Al-Mshhdani, B.A.; Grounds, M.D.; Arthur, P.G.; Terrill, J.R. A Blood Biomarker for Duchenne Muscular Dystrophy Shows That Oxidation State of Albumin Correlates with Protein Oxidation and Damage in Mdx Muscle. Antioxidants 2021, 10, 1241. [Google Scholar] [CrossRef] [PubMed]
- Witort, E.; Capaccioli, S.; Becatti, M.; Fiorillo, C.; Batignani, G.; Pavoni, V.; Piccini, M.; Orioli, M.; Carini, M.; Aldini, G.; et al. Albumin Cys34 adducted by acrolein as a marker of oxidative stress in ischemia-reperfusion injury during hepatectomy. Free Radic. Res. 2016, 50, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, H.; Schiffman, C.; Gunter, M.J.; Naccarati, A.; Polidoro, S.; Dagnino, S.; Dudoit, S.; Vineis, P.; Rappaport, S.M. Cys34 Adductomics Links Colorectal Cancer with the Gut Microbiota and Redox Biology. Cancer Res. 2019, 79, 6024–6031. [Google Scholar] [CrossRef] [PubMed]
- Tada, S.; Yasukawa, K.; Yatomi, Y.; Uchiki, T. A simple colorimetric assay to determine the concentration and proportion of human mercaptalbumin. Pract. Lab. Med. 2022, 31, e00281. [Google Scholar] [CrossRef] [PubMed]
- Lim, Z.X.; Duong, M.N.; Boyatzis, A.E.; Golden, E.; Vrielink, A.; Fournier, P.A.; Arthur, P.G. Oxidation of cysteine 34 of plasma albumin as a biomarker of oxidative stress. Free Radic. Res. 2020, 54, 91–103. [Google Scholar] [CrossRef]
- Teder, K.; Maddison, L.; Soeorg, H.; Meos, A.; Karjagin, J. The Pharmacokinetic Profile and Bioavailability of Enteral N-Acetylcysteine in Intensive Care Unit. Medicina 2021, 57, 1218. [Google Scholar] [CrossRef]
- Olsson, B.; Johansson, M.; Gabrielsson, J.; Bolme, P. Pharmacokinetics and bioavailability of reduced and oxidized N-acetylcysteine. Eur. J. Clin. Pharmacol. 1988, 34, 77–82. [Google Scholar] [CrossRef]
- Nolin, T.D.; Ouseph, R.; Himmelfarb, J.; McMenamin, M.E.; Ward, R.A. Multiple-dose pharmacokinetics and pharmacodynamics of N-acetylcysteine in patients with end-stage renal disease. Clin. J. Am. Soc. Nephrol. 2010, 5, 1588–1594. [Google Scholar] [CrossRef]
- Liu, Y.M.; Liu, Y.; Lu, C.; Jia, J.Y.; Liu, G.Y.; Weng, L.P.; Wang, J.Y.; Li, G.X.; Wang, W.; Li, S.J.; et al. Relative bioavailability of generic and branded acetylcysteine effervescent tablets: A single-dose, open-label, randomized-sequence, two-period crossover study in fasting healthy Chinese male volunteers. Clin. Ther. 2010, 32, 2097–2105. [Google Scholar] [CrossRef]
- Kurano, M.; Yasukawa, K.; Ikeda, H.; Aoki, J.; Yatomi, Y. Redox state of albumin affects its lipid mediator binding characteristics. Free Radic. Res. 2019, 53, 892–900. [Google Scholar] [CrossRef]
- Cazzola, M.; Calzetta, L.; Page, C.; Jardim, J.; Chuchalin, A.G.; Rogliani, P.; Matera, M.G. Influence of N-acetylcysteine on chronic bronchitis or COPD exacerbations: A meta-analysis. Eur. Respir. Rev. 2015, 24, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Duailibi, M.S.; Cordeiro, Q.; Brietzke, E.; Ribeiro, M.; LaRowe, S.; Berk, M.; Trevizol, A.P. N-acetylcysteine in the treatment of craving in substance use disorders: Systematic review and meta-analysis. Am. J. Addict. 2017, 26, 660–666. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Placebo | NAC | |||
|---|---|---|---|---|
| Minutes | Thio-HSA | HSA-SH | Thio-HSA | HSA-SH |
| 0 | 21.50 ± 3.30 | 60.10 ± 5.22 | 20.70 ± 4.31 | 60.97 ± 3.98 |
| 30 | 21.97 ± 3.32 | 60.15 ± 4.29 | 16.28 ± 3.23 § | 64.63 ± 3.43 § |
| 60 | 22.37 ± 3.43 | 59.43 ± 5.27 | 15.75 ± 4.60 § | 64.85 ± 3.33 § |
| 120 | 22.98 ± 3.95 | 58.62 ± 5.30 | 17.08 ± 5.56 # | 63.78 ± 4.47 # |
| 240 | 21.92 ± 3.03 | 59.58 ± 4.94 | 18.98 ± 5.18 | 61.62 ± 4.68 |
| 360 | 21.12 ± 4.12 | 60.30 ± 6.23 | 18.43 ± 4.85 | 61.72 ± 4.14 |
| 1440 | 21.17 ± 5.46 | 59.48 ± 9.135 | 20.98 ± 4.93 | 60.02 ± 4.45 |
| Placebo | NAC | |||
|---|---|---|---|---|
| Minutes | Thio-HSA | HSA-SH | Thio-HSA | HSA-SH |
| 0 | 20.05 ± 1.96 | 65.40 ± 1.92 | 18.73 ± 1.72 | 66.65 ± 2.59 |
| 30 | 21.15 ± 1.92 | 64.58 ± 2.04 | 5.73 ± 1.69 § | 75.52 ± 3.46 § |
| 60 | 21.98 ± 1.65 | 63.42 ± 2.00 | 8.37 ± 1.73 § | 74.35 ± 3.78 # |
| 120 | 20.98 ± 1.96 | 64.83 ± 3.14 | 11.37 ± 1.86 § | 71.67 ± 3.44 |
| 240 | 19.87 ± 1.81 | 65.82 ± 3.31 | 13.80 ± 1.92 § | 69.32 ± 4.08 |
| 360 | 18.80 ± 2.81 | 67.13 ± 4.08 | 15.46 ± 1.52 * | 67.84 ± 2.65 |
| 1440 | 19.37 ± 2.74 | 66.60 ± 4.40 | 19.73 ± 2.36 | 64.35 ± 3.68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altomare, A.A.; Brioschi, M.; Eligini, S.; Bonomi, A.; Zoanni, B.; Iezzi, A.; Jemos, C.; Porro, B.; D’Alessandra, Y.; Guarino, A.; et al. N-Acetylcysteine Regenerates In Vivo Mercaptoalbumin. Antioxidants 2022, 11, 1758. https://doi.org/10.3390/antiox11091758
Altomare AA, Brioschi M, Eligini S, Bonomi A, Zoanni B, Iezzi A, Jemos C, Porro B, D’Alessandra Y, Guarino A, et al. N-Acetylcysteine Regenerates In Vivo Mercaptoalbumin. Antioxidants. 2022; 11(9):1758. https://doi.org/10.3390/antiox11091758
Chicago/Turabian StyleAltomare, Alessandra Anna, Maura Brioschi, Sonia Eligini, Alice Bonomi, Beatrice Zoanni, Ada Iezzi, Costantino Jemos, Benedetta Porro, Yuri D’Alessandra, Anna Guarino, and et al. 2022. "N-Acetylcysteine Regenerates In Vivo Mercaptoalbumin" Antioxidants 11, no. 9: 1758. https://doi.org/10.3390/antiox11091758
APA StyleAltomare, A. A., Brioschi, M., Eligini, S., Bonomi, A., Zoanni, B., Iezzi, A., Jemos, C., Porro, B., D’Alessandra, Y., Guarino, A., Omodeo Salè, E., Aldini, G., Agostoni, P., & Banfi, C. (2022). N-Acetylcysteine Regenerates In Vivo Mercaptoalbumin. Antioxidants, 11(9), 1758. https://doi.org/10.3390/antiox11091758