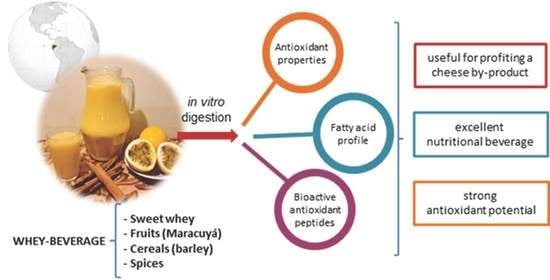
3.1. Nutritional Composition of the Colada Beverage
Beverages based on liquid whey mixed with fruit juices are a great way to use a by- product of the cheese industry which is full of nutrients. Composition of the main ingredient, the whey, depends on the origin of the milk, type of cheese, and the cheese processing method [
33]. Based on pH, whey may be classified as acidic whey (pH < 5) or sweet whey (pH 6–7); since acidic pH may cause protein sedimentation due to heat treatment, sweet whey is considered more suitable as the raw material for the whey-based beverages [
15].
The chemical composition of the whey beverage is depicted in
Table 1. The Ecuadorian technical standard NTE INEN 2609 establishes the requirements for whey-based beverages, i.e., those whose main ingredient is whey and are intended for direct consumption (FAO, 2022). The protein content found in the
Colada (1.29%) significantly exceeds the minimum percentage established in Ecuadorian regulations of 0.4%, which reinforce its nutritional value. The result is within the range found in the bibliography for whey beverages, which may vary from 0.94% in beverages prepared with hydrolyzed collagen, milk powder and sucrose [
34] to 1.21–1.61% in traditional whey blended with fruit juices [
35].
The
Colada beverage showed an ash content of around 0.5%; this value, that provides an estimate of the minerals present in the beverage, was slightly higher than the average content observed for whey (0.25–0.4%, [
36]). Whey is recognized as a source of valuable minerals, especially sodium and potassium, followed by calcium and magnesium [
37]. Due to the addition of barley, the
Colada beverage also provided high amounts of phosphorus, which may reinforce the mineral contribution to the diet.
Analysis of the amino acid content revealed that the proteins of the
Colada are rich in essential amino acids, with the exception of valine and methionine (tryptophan was not determined) (
Table 2). The major amino acids were glutamic, aspartic, leucine, proline and lysine. Methionine and histidine were the limiting amino acids. The amino acid composition of the beverage is mainly determined by its principal component, sweet whey. The amino acid composition of whey depends on the milk origin and on the stage of lactation that influences on the β-lactoglobulin/α-lactoalbumin ratio [
38].
3.2. Proteolysis and Peptide Release during In Vitro Digestion
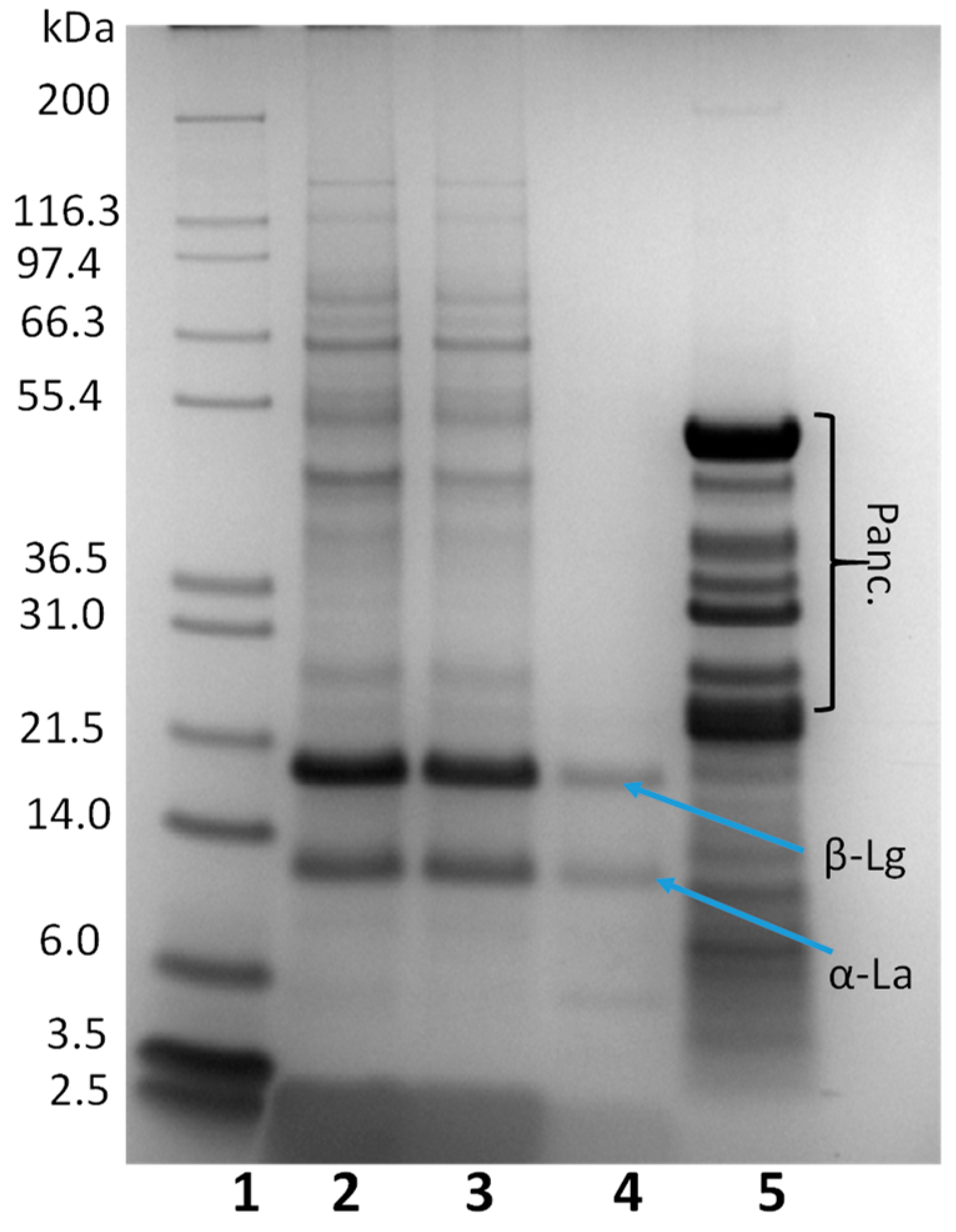
Once the nutritional composition of the Colada had been established, the beverage was digested, following the INFOGEST protocol in order to study the DH and the changes in the protein profile. A negative control of the digestion was performed, with no enzymes to confirm there is no protein autolysis during the process (data not shown). The proteolysis started after the gastric phase with a 14 ± 2% DH and was much higher after the intestinal phase, with a 57.69 ± 3% DH measured by OPA. The protein profile after in vitro digestion was studied by SDS-PAGE (
Figure 1) and exclusion molecular chromatography (SEC) (
Figure 2). As expected, there was no protein degradation after the oral phase being the protein profile identical to the undigested
Colada (lanes 2 and 3). Proteolysis started with the action of pepsin during the gastric digestion (lane 4). Most proteins of a molecular weight higher than 24 KDa were completely digested after 2 h. Two major protein bands at an apparent molecular weight around 21 and 14 KDa were observed, corresponding to β-lactoglobulin and α-lactoabumin. These two proteins have a globular structure which prevents the access of enzymes to potential cleavage sites and have been described as resistant to the action of the pepsin [
39,
40]. The great number of proteins of apparent molecular weight higher than 20 KDa, observed in the SDS-PAGE after the complete in vitro digestion, correspond to pancreatin background (lane 5). Most of the proteins are digested, and only small peptides are present after the digestive process.
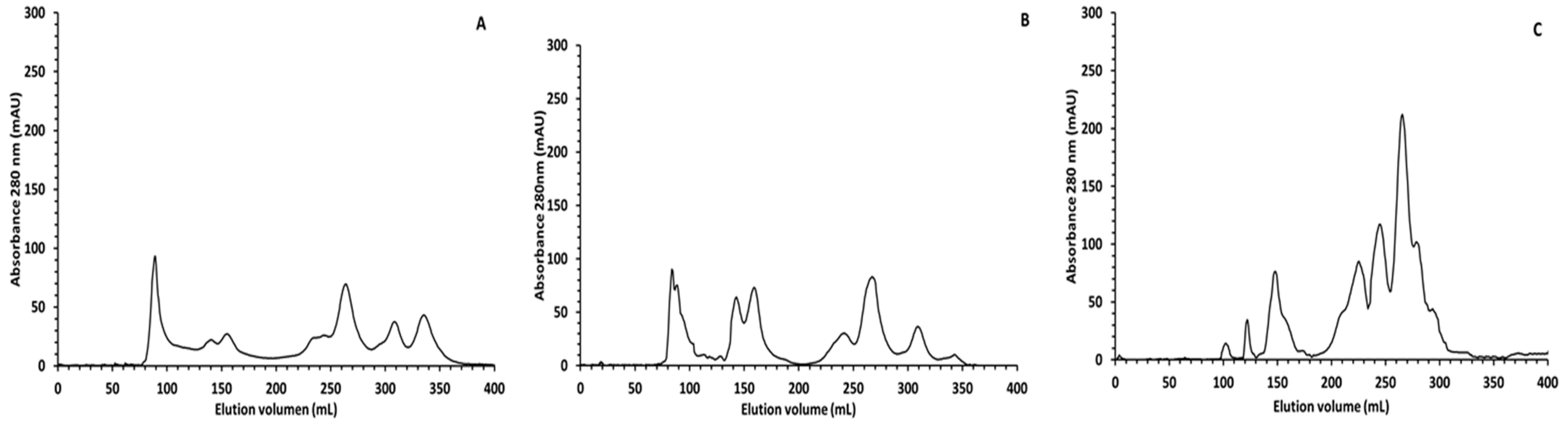
In order to have a better picture of the peptide profile after digestion, SEC was used, and a control sample (digestion without sample) was ran and subtracted from the chromatogram of the completely digested sample to reduce the enzyme background observed in SDS-PAGE. The SEC chromatograms give valuable information on size distributions of proteins, large peptides, and smaller protein degradation products. The SEC profile of the
Colada (
Figure 2) indicated that the main difference in the oral phase (part A) compared to the gastric phase (part B) was observed in the range of 40–13 KDa (130–180 mL of elution volume). The two main peaks observed at 145 mL and 165 mL probably corresponded to β-lactoglobulin and α-lactoalbumin that were still undigested in the gastric phase, as it was observed in the SDS-PAGE. Although the digestion process starts in the gastric phase, the great majority of proteins were digested in the intestinal phase (part C). After this phase there was a great increase in the intensity in the peaks corresponding to peptides ≤6 KDa (240 mL of elution volume) and all the proteins with an apparent molecular weight ≥40 KDa were hydrolyzed into smaller peptides/free aa.
3.3. Antioxidant Properties
Currently, it is widely accepted that the effect of any dietary component is defined by the bioavailable amount rather than the dose ingested, and that bioaccessibility constitutes a prerequisite and the most influential factor of bioavailability [
41]. Thus, determination of bioaccessibility in vitro reflects reasonably well bioavailability in vivo [
42].
The digestive process had a general positive effect on the antioxidant properties of the
Colada, represented by significant increases of around 2 fold on ABTS and FRAP assays after digestion, although a slight decline was observed in DPPH values (
Table 3). Moreover, a strong increment of TPC was observed (>6 fold).
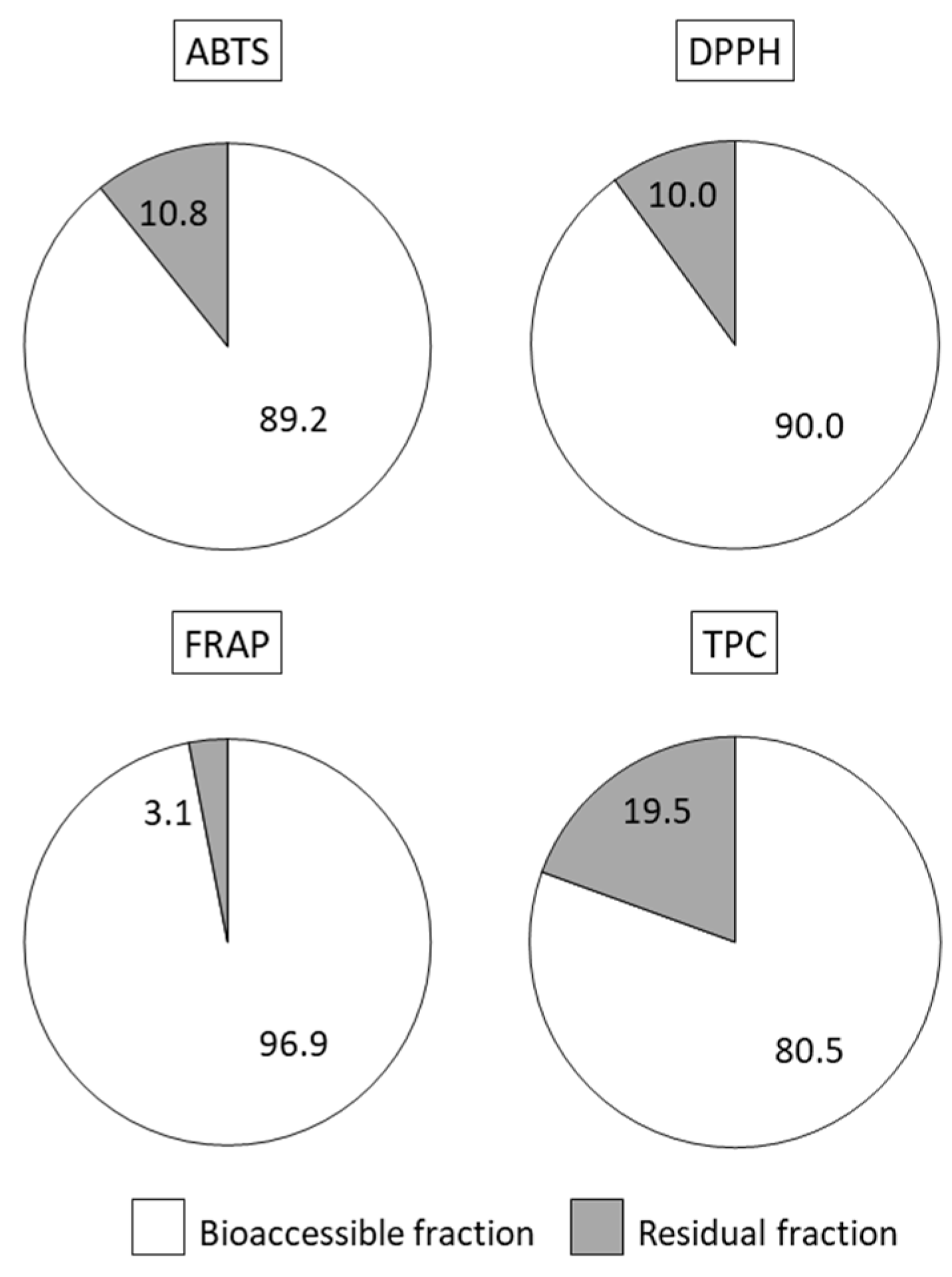
Both antioxidant properties and polyphenols post digestion were mainly located in the bioaccessible fraction (90–97% of ABTS, DPPH and FRAP; 80% of polyphenols,
Figure 3), which included compounds released from the food matrix that are potentially absorbable from the lumen. In the residual fraction, usually discarded when studying bioavailability of bioactive compounds, there still remained low but appreciable amounts of antioxidant activity (10% of radical scavenging ability) and phenolic compounds (20%). These may exert antioxidant and anti-inflammatory local action [
43] or be metabolized by the intestinal microbiota in potentially absorbable complexes [
44].
Usually, antioxidant properties are measured in vitro by the ability to transfer hydrogen atoms or electrons to an oxidant, and ABTS, DPPH and FRAP methods are commonly applied with this aim. However, the use of cell model systems, such as the Caco-2 intestinal cell line, is recommendable to better assess the in vivo antioxidant potential of dietary compounds [
37]. Preincubating Caco-2 cells for 2 h with the BF of the
Colada beverage significantly reduced ROS generation in basal conditions (
Figure 4A). In addition, a protective effect against induced oxidative stress (25% of decrease in ROS production) was observed (
Figure 2B), supporting the antioxidant activity of the
Colada digest at the cell level.
Our results showing the increasing release of bioactive antioxidant compounds during digestion agree with those previously observed in oils [
21,
27] and other foods [
45], although disappointing data have also been observed in coffee extracts [
46]. The antioxidant behavior of dietary components during digestion seems to be strongly dependent on the food matrix and the kind of antioxidant compounds; phytochemicals (carotenoids and polyphenols, among others) from fruit and vegetables are traditionally designed as the main contributors [
47]. In the last years, whey-derived peptides with oxidation inhibitory activities have received special attention [
37,
48]. Although antioxidant properties of whey peptides and their hydrolysates have been deeply studied, little information exists on the effect of the digestion process and their interactions with other components when whey is included as a part of a complex food or beverage. Previous studies [
48] demonstrated that antioxidant properties of isolated bovine whey proteins, tested by ABTS, FRAP and ORAC methods, significantly increase after simulated gastrointestinal digestion, α-lactalbumin having the highest antioxidant post-digestion activity. Therefore, the increasing radical scavenging ability (by ABTS) and ferric reducing power found in the present assay could be at least partly due to the digested whey proteins of the
Colada, mainly α-lactoalbumin, as indicated by the results obtained by SDS-PAGE.
Interestingly, in Caco-2 cells, isolated digested whey proteins were unable to reduce induced ROS formation [
8], although were effective in HT-29 cells and for stimulating antioxidant enzymes. Thus, in our study, other food components of the
Colada digest could have contributed to the protecting ability against radical generation observed in Caco-2 cells.
It has been shown that combination of whey with additional antioxidant ingredients (vitamin B12, astaxanthin and plant extracts) improves the bioactivity of the formulated beverages [
49], and that values of antioxidant activity in whey-based drinks added with fruit and vegetables are positively correlated with the polyphenol content [
17]. Results of the present assay are comparable with those found for homemade beverages prepared with 50% whey and with fruits (pear, cherry and banana) and vegetables (carrot and parsley) added, in which DPPH and ORAC values were around 0.78 and 0.71 mM trolox Eq./kg, respectively [
17]. Therefore, it seems clear that antioxidant properties of the intact
Colada were due, in addition to the whey proteins, to antioxidant compounds, especially polyphenols, provided by barley and Maracuyá [
18,
19]. Moreover, polyphenol extracts of Maracuyá have shown preventive effects against induced barrier dysfunction in Caco-2 cells by increasing transepithelial electrical resistance [
50], a reliable indicator of the normal and functional state of cell membranes which is linked to reducing level of oxidative stress and ROS accumulation [
51].
However, these antioxidant properties may be altered during gastrointestinal transit. Interestingly, it has been reported that antioxidant activity after simulated digestion increases in whey beverages but decreases when drinks were added with additional antioxidant compounds [
49]. It seems that adding carotenoids and polyphenols interact with the digestibility of whey proteins, reducing the release of antioxidant peptides and amino acids [
18] which, in turn, decrease the antioxidant activity of the formulation [
49]. This fact would explain the decreasing value of DPPH post digestion found in the
Colada beverage. However, the possible negative influence of antioxidant phytochemicals seems to be counteracted with the high antioxidant potential of the whey proteins, since whey was the major component of the formulation, leading to strongly increased ABTS and FRAP values. On the other hand, discrepancies between different antioxidant assays (ABTS and FRAP) with DPPH have been previously observed [
48].
The digestive process also produces structural transformations in food polyphenols, mainly due to changes of pH and interactions with other compounds, which may alter their bioaccessibility [
52]. Anthocyanins, the major polyphenols of Maracuyá, are highly stable during simulated gastric digestion but are degraded to different catabolic products during the intestinal phase [
53]. Therefore, the large increase found in TPC of the
Colada BF could be due to the different chemical structures of the existing polyphenols and their biotransformation during digestive conditions. In addition, the fact that the Folin–Ciocalteu reagent, used for the determination of polyphenolic compounds, could react with amino acids from whey digest leading to overestimation of results [
17], cannot be discarded.
3.4. The Lipid Fraction
The type of dietary fatty acids (FA) affects the physiological body response; saturated FA (SFA) have been related to adverse health effects, whereas unsaturated fatty acids, especially monounsaturated (MUFA) and n3 polyunsaturated (PUFA), are thought to be protective. They reduce the risk of cardiovascular and inflammatory diseases, although PUFA are more susceptible to being oxidized [
54].
The FA profile and the related indexes of the
Colada beverage before and after the in vitro digestion (BF) are shown in
Table 4 and
Figure 5, respectively.
Before digestion, the major FA of the
Colada was palmitic acid (C16:0, 32%), followed by oleic (C18.1n9, 28%) and stearic acid (C18:0, 17%); this is in agreement with the main FA observed in whey [
55] and in milk fat [
56]. Total SFA represented the major fraction of the undigested sample (65%), whereas MUFA and PUFA contributed 31 and 4%, respectively. After digestion, significant changes in the FA profile of the beverage were observed: SFA were drastically reduced nearly to the half, whereas total MUFA and PUFA increased approximately 2- and 3-fold, respectively, compared with values pre-digestion.
Compositional studies of whey and whey beverages have been mainly focused on the protein components, and little attention has been given to the lipid fraction, probably due to its minority presence in the final product. Moreover, there is a lack of information concerning the FA profile of whey or whey-based drinks after the digestive process and the subsequent bioavailability. Santillo et al. [
57] showed that the percentage distribution of FA in in vitro-digested milk from different species did not reflect the patterns of the corresponding milk sources, which is according with our results. This finding might be explained by different reasons. On the one hand, Ca may affect FA bioaccessibility since, at pH intestinal conditions, it reacts with saturated long chain FA (>14 C) and forms the corresponding insoluble soaps, which significantly reduces their availability [
56]. Therefore, the high Ca level of the
Colada could have induced the precipitation of myristic (C14:0), palmitic (C16:0), and stearic (C18:0) acid as non-soluble soaps in the RF, thus decreasing their levels in the soluble fraction.
On the other hand, the digestion may induce lipid oxidation, mainly due to acidic pH in the gastric fluid, the disintegration of the food matrix or the action of digestive enzymes [
58]. Oxidation of PUFA led to the formation of lipid oxidation by-products and subsequent decrease of PUFA bioaccessibility, especially n3 [
59]. However, after digestion of the
Colada, higher levels of n6 and n3 in the BF were observed. This may be attributed to the high level of antioxidant compounds present in the formulation; specifically, it has been suggested that the addition of polyphenols during simulated digestion limits the lipid oxidation and promotes the bioavailability of non-oxidized PUFA [
59]. Moreover, the food matrix also may impact on the release of FA during in vitro intestinal digestion. In this line, it has been shown that adding barley to a meal increases the release of long-chain PUFA and their percentage contribution to the total FA in the digest [
60]. Therefore, the ingredients added to the whey beverage
Colada, such as barley and Maracuyá polyphenols, could have contributed to increase the PUFA proportions in the BF after the in vitro digestion.
After digestion of
Colada, strong increases in C18:1n9 and C18:2n6 were also observed. These FA are of interest in human nutrition, since oleic acid has noticeable health effects in preventing degenerative diseases, and linoleic acid an essential FA precursor of other important long chain FA [
61]. Moreover, substitution of dietary saturated fat by oleic acid and/or PUFA produces cardiovascular benefits by reducing blood lipids [
61].
Hence, the in vitro digestion of the Colada induces positive transformation in the composition and bioaccessibility of the FA, which could be at least partly due to the presence of antioxidant compounds provided by its different ingredients.
3.5. Peptide Identification with Antioxidant Potential Activity
Since antioxidant properties of the
Colada could be also determined by the protein composition, we analyzed the peptides released after the in vitro digestion. Bioactive peptides are encrypted amino acid sequences inactive in the native protein that could be released through different processes including gastrointestinal digestion [
62]. Ingested proteins from the
Colada were hydrolyzed by proteinases present in the gastrointestinal tract that are used in the in vitro digestion process to produce peptides of various sizes, as shown in
Figure 2. To identify peptides potentially responsible for the antioxidant activity observed in the digested sample, the bioaccesible fraction was analyzed by NanoLC-MS/MS. A total of 251 peptides of different lengths from 5 to 25 aminoacids were identified. The peptides released after the digestion of the
Colada beverage not only originated from whey proteins but also from barley (Hordeon Vulgare) used in the formulation. Surprisingly, many of the peptides identified were fragments of β-casein normally not present in sweet whey. The presence of casein peptides in the digestion of the beverage made using sweet whey, suggests the leakage of casein into the whey fraction in the process of cheese making, a fact that has been previously described [
8]. Peptides from κ-casein were also identified and the longer peptides were obtained from α-lactoalbumin and β-lactoglobulin. These last proteins are described as the least digestible proteins from whey. The sequences of the generated peptides were analyzed through BIOPEP a database of biologically active peptide sequences [
31]. The peptides analyzed showed homology with several bioactive peptide sequences, whose activities include ACE inhibition, DDP-IV enzyme inhibition, antithrombotic, antibacterial and antioxidant effects. Here we wanted to highlight those peptides with antioxidant activity (
Table 5).
Antioxidant peptides derived from milk proteins have been mostly associated with bovine casein [
63,
64]. The in vitro digestion of the
Colada produced 11 peptides with sequences described as antioxidant activity from β-casein (
Table 5). The antioxidant activity of casein derived peptides has been extensively studied and described [
63]. One of the peptides identified, YPFPGPI, which shares sequence with other peptides found, has been recently described as a potent antioxidant [
65]. Although antioxidant peptides derived from milk proteins have been mostly associated with bovine casein, hydrolysis of whey proteins may also result in the production of antioxidant peptides [
63,
64]. In our digested beverage we have identified 18 peptides from β-lactoglobulin, all with different biological activities but only 7 with potential antioxidant activity (
Table 5). β-lactoglobulin is the major whey protein and biological active peptides derived from this protein is an area of intense research [
66,
67]. Four of the longer peptides identified (QTMKGLDIQKVAGTWYSLAMAASD, VAGTWYSLAMAASDISLLDAQSA, GLDIQKVAGTWYSLAMAASDISLL, DIQKVAGTWYSLAMAASDISLLDA) contain in their sequence previously described potent antioxidant peptides (WYSLAMAASDI, WYSLAMA, WYSLAM, WYSLA, WYSL, WYS, and WY). One of these peptides, WYSLAMAASDI, possessed higher radical scavenging activity than butylated hydroxyanisole (BHA), a potent antioxidant used in the food industry [
68]. Only two of the sequenced peptides, GLDIQKVAGTWYSLAMAASDISLL and DIQKVAGTWYSLAMAASDISLLDA have a relatively high enough FRS score, predicted through AnOxprep-1.0, to be considered as potential antioxidant peptides. The presence of tyrosine and tryptophan could determine its potent activity, as these two amino acids have been reported to be the main contributors to the peroxyl radical-scavenging activity of food-derived peptides [
69]. All of these peptides could still be considered as potential antioxidants since they could be further hydrolyzed by peptidases present in the intestinal brush border. The action of the enzymes present in the brush border is very intense and could release new peptides available for absorption with important biological activity [
70,
71].
The second most important milk protein, α-lactalbumin, is also known to be an important source of essential amino acids that present bioactive peptides encrypted in their sequence [
8]. Ten peptides were identified from α-lactalbumin, all with previously described biological activities but none of them noted as antioxidant. Previous studies have shown that the antioxidant activity of α-lactalbumin increases after hydrolysis with alcalase and even more when this hydrolysate was subjected to a gastrointestinal digestion in vitro [
72,
73]. Very recently the sequence of these peptides has been reported [
74]. Five of the peptides identified in our samples, LDDDL, LDDDLTDD, DDDLTDDI, LDDDLTDDI and DDDLTDDIM are fragments of the peptides previously identified as potent antioxidants (
Table 6). The peptide VSLPE is a fragment of the antioxidant peptide VSLPEW sequenced earlier [
75].
Despite the fact that α-lactoalbumin has been described as a potent antioxidant [
12,
69,
75] only a few peptides have been described and none of them obtained after gastrointestinal digestion. Previous studies on peptides from this protein obtained by hydrolysis with Corolase PP
®, a complex mixture of enzymes similar to pancreatin, proved its antioxidant activity but did not purify or identify any of the responsible peptides in the hydrolyzed fraction [
69]. It is well known that the potential antioxidant activity of the peptides depends on the abundance of certain amino acids and the order in the sequence. Basic and acidic amino acid residues can act as hydrogen donors, and hydrophobic residues facilitate access to hydrophobic radical species. Hydrophobic amino acids such as Ala (A), Pro (P), Val (V), Ile (I), Leu (L), Phe (F), Tyr (Y) can contribute on antioxidant properties [
12]. All the peptides found from α-lactalbumin have leucine present in its sequence, and could be in theory considered potential antioxidants, but the predictions using the Anoxprep-0.1 tool gives very low values for these peptides (
Table 6). From our results, we can conclude that the peptides obtained after digestion of α-lactalbumin in the
Colada need further investigation in order to establish their antioxidant potential.
Twenty-three peptides of barley proteins were also identified in our samples (
Table 7).
None of these peptides were reported as antioxidant in the BIOPEP database, probably because it is a protein source not as well studied as milk. In order to establish the potential bioactivity of the sequenced peptides, these were analyzed using PeptideRanker. This bioactive peptide predictor provides a computational prediction, assigning scores between 0 and 1 to peptides based on the probability of being bioactive from the N-to-1 neural network (N1-NN). This is computed based on the peptide primary sequence trained from different bioactive peptide databases. Any peptide possessing a score above the 0.5 threshold is labeled as bioactive [
76]. Sixteen of these peptides had a score above 0.5 and three of them, PQQPFP, PQQPPFG, QPQPFP, with a very high probability of being biologically active with a score above 0.9. Those peptides labelled as potentially bioactive were analyzed for their potential antioxidant activity using the Anoxprep-0.1 tool (
Table 7). All of the peptides showed a moderate probability of being antioxidants, but this will require further investigation.





 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}