
Yomogin, Isolated from Artemisia iwayomogi, Inhibits Neuroinflammation Stimulated by Lipopolysaccharide via Regulating MAPK Pathway


 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction and Isolation of Yomogin
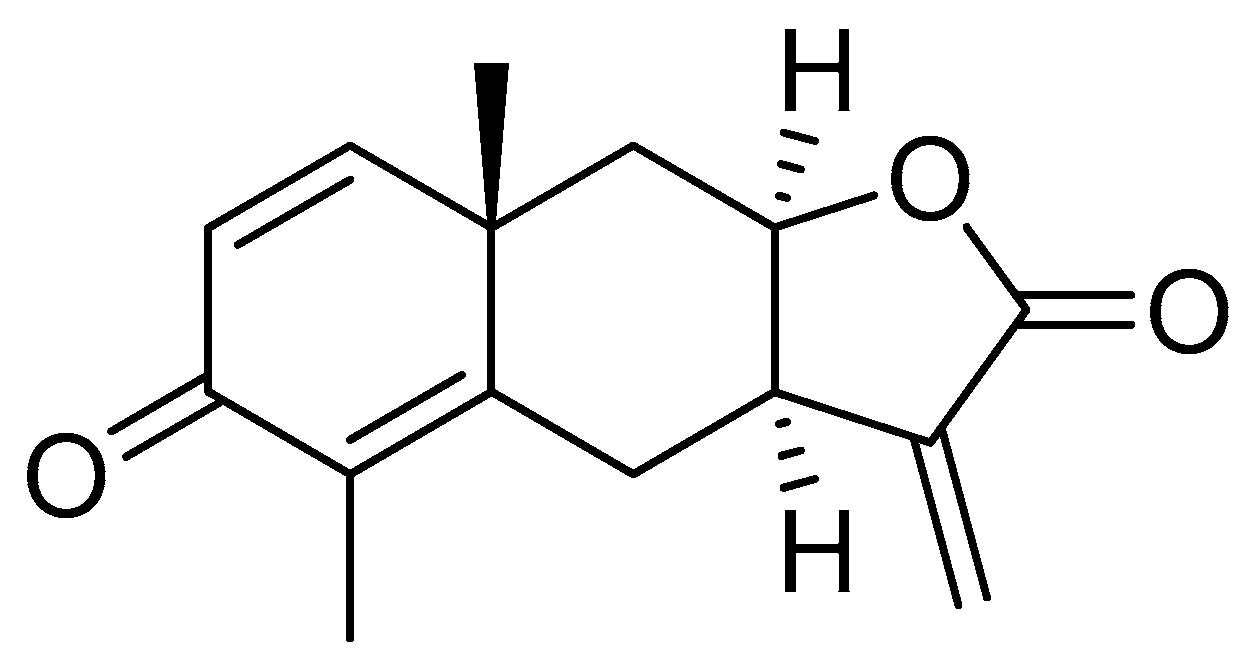
Yomogin
2.3. Cell Culture and Measurement of Cell Viability
2.4. Measurement of Extracellular NO
2.5. Molecular Docking Calculation
2.6. Western Blot Analysis
2.7. RNA Extraction and qRT-PCR Analysis
2.8. Animals and Treatment
2.9. Experimental Design of Animal Study
2.10. FST
2.11. Brain Tissue Preparation
2.12. Immunofluorescence Staining
2.13. Statistical Analysis
3. Results
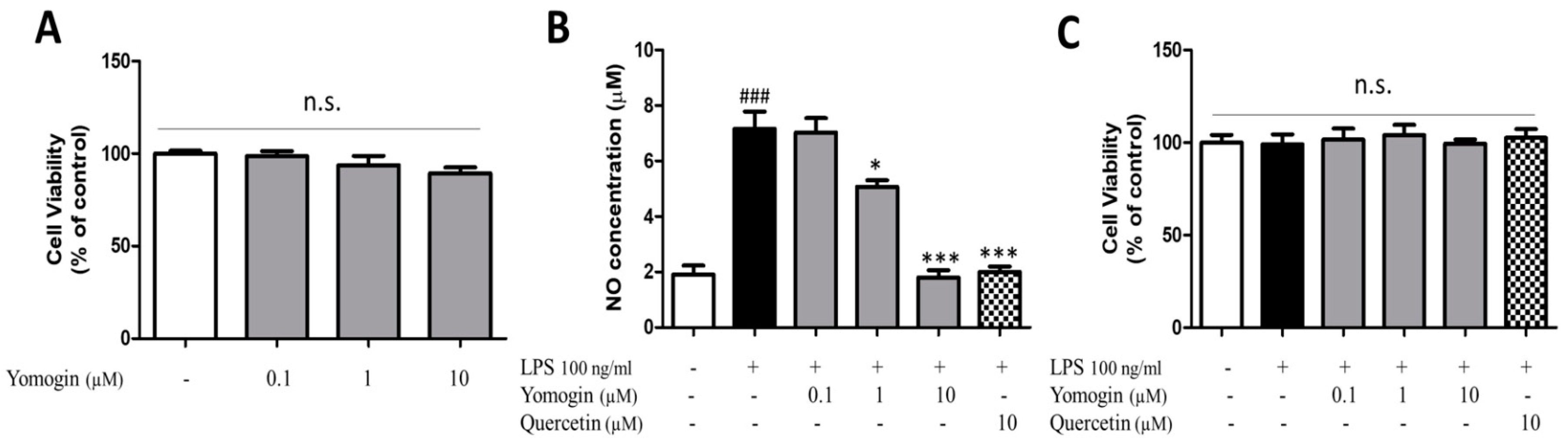
3.1. Effects of Yomogin on Cell Viability and NO Production in LPS-Stimulated BV2 Microglial Cells
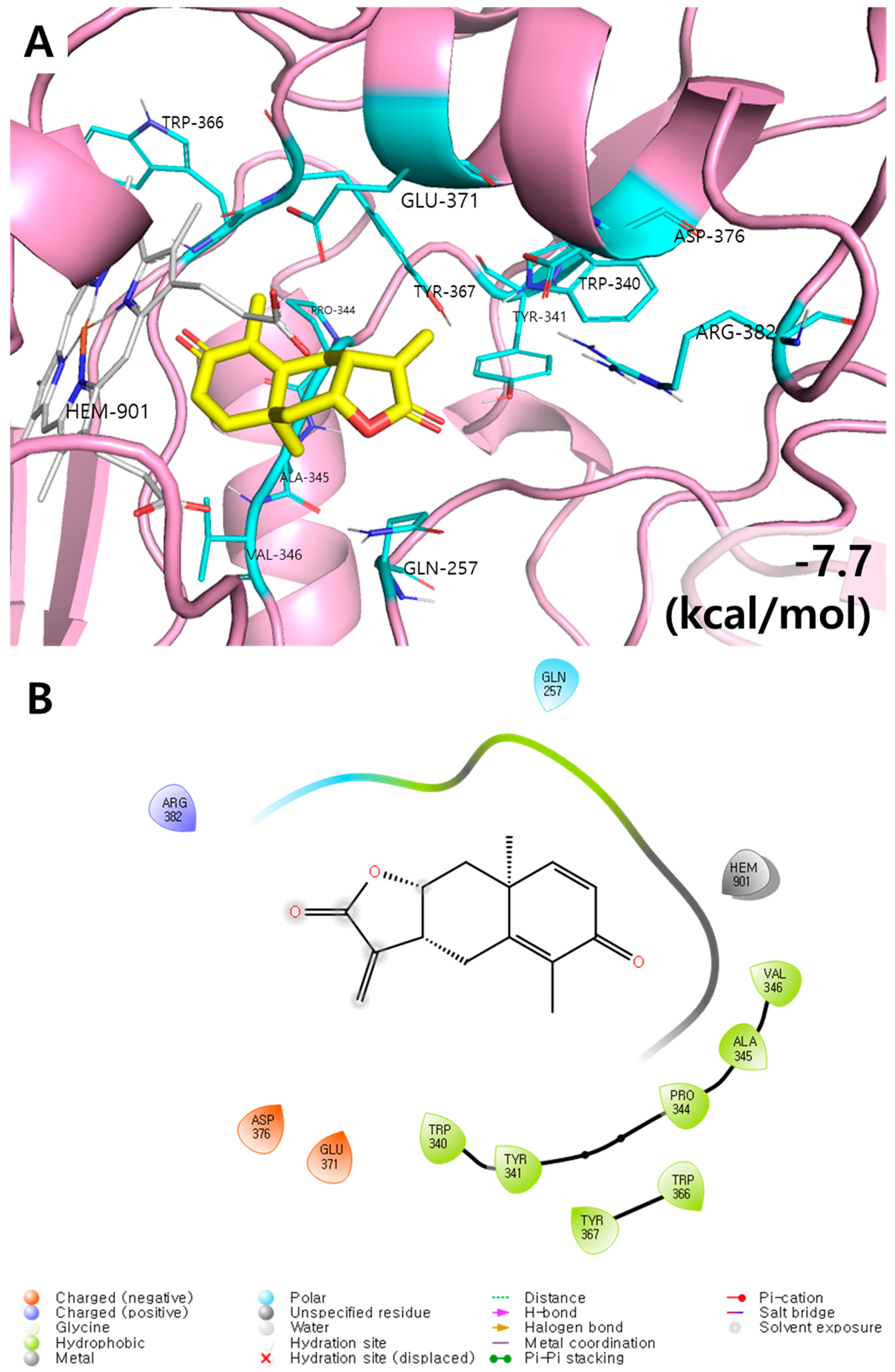
3.2. Molecular Docking Results of Yomogin and iNOS Enzymes
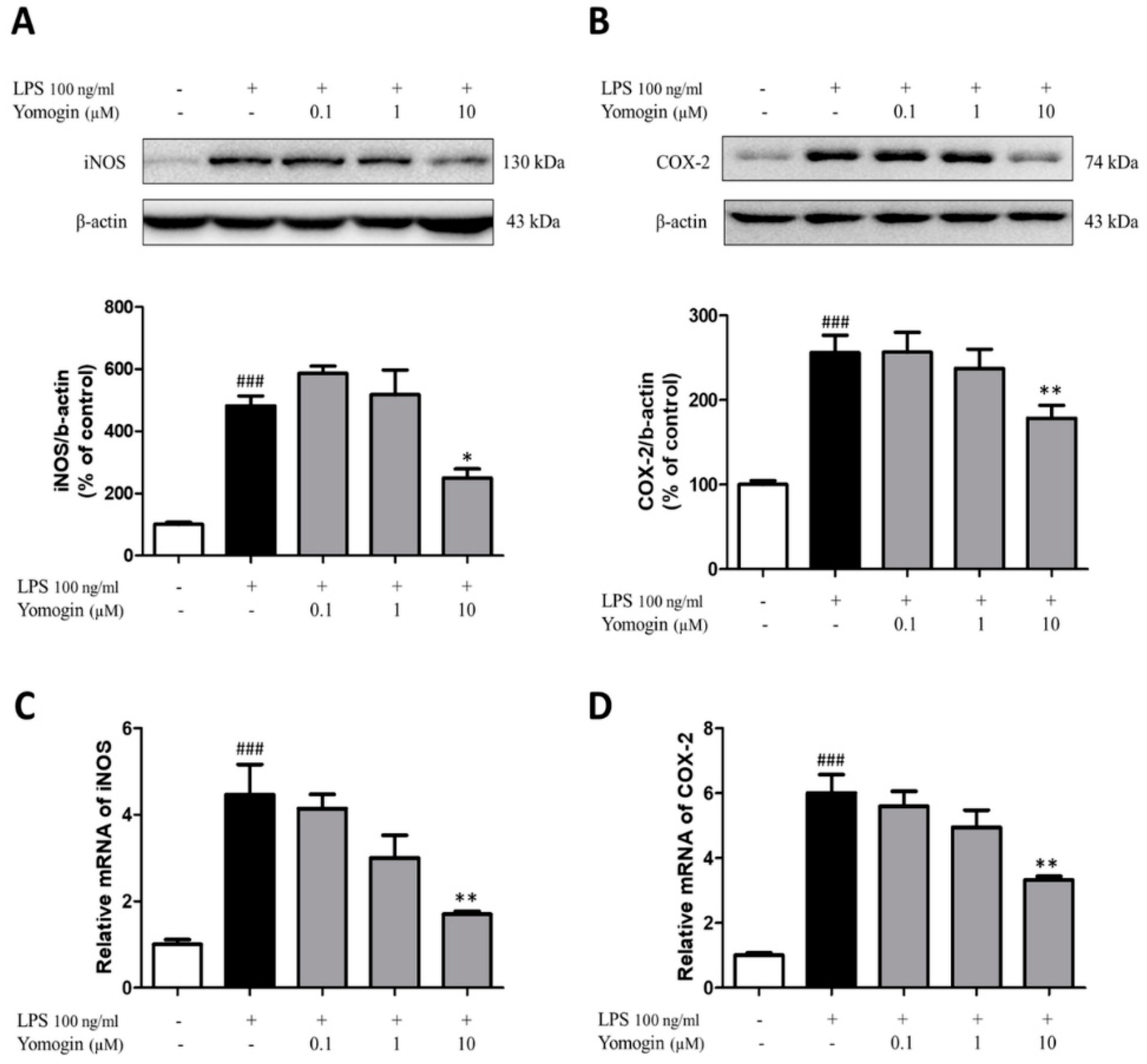
3.3. Effects of Yomogin on Expressions of iNOS and COX-2 in LPS-Stimulated BV2 Microglial Cells
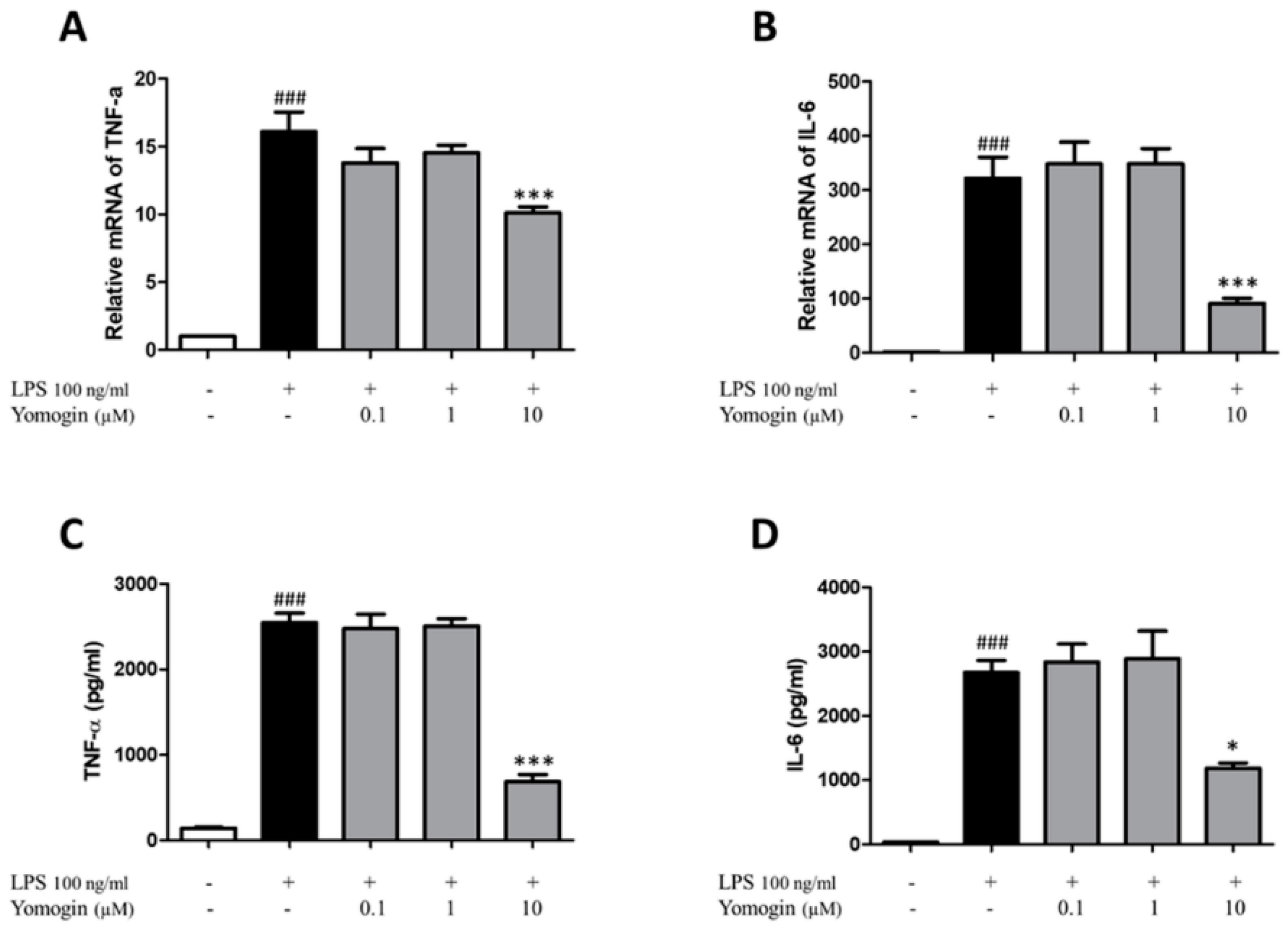
3.4. Effects of Yomogin on Secretion of TNF-α and IL-6 in LPS-Stimulated BV2 Microglial Cells
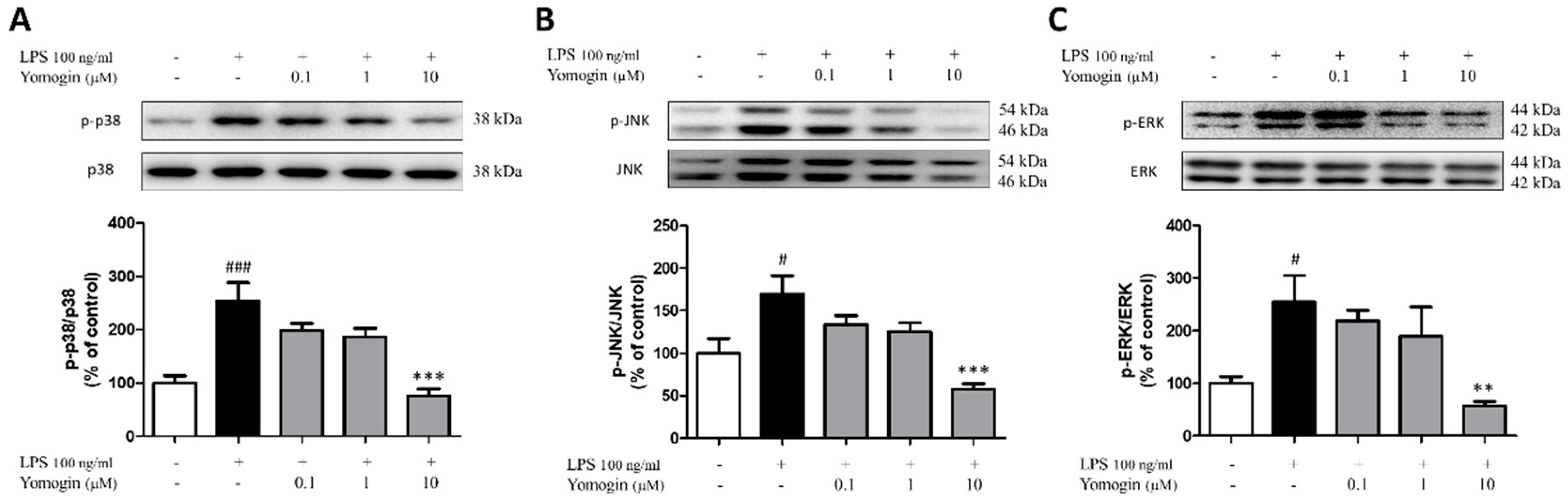
3.5. Effects of Yomogin on MAPK Signaling Pathway in LPS-Stimulated BV2 Microglial Cells
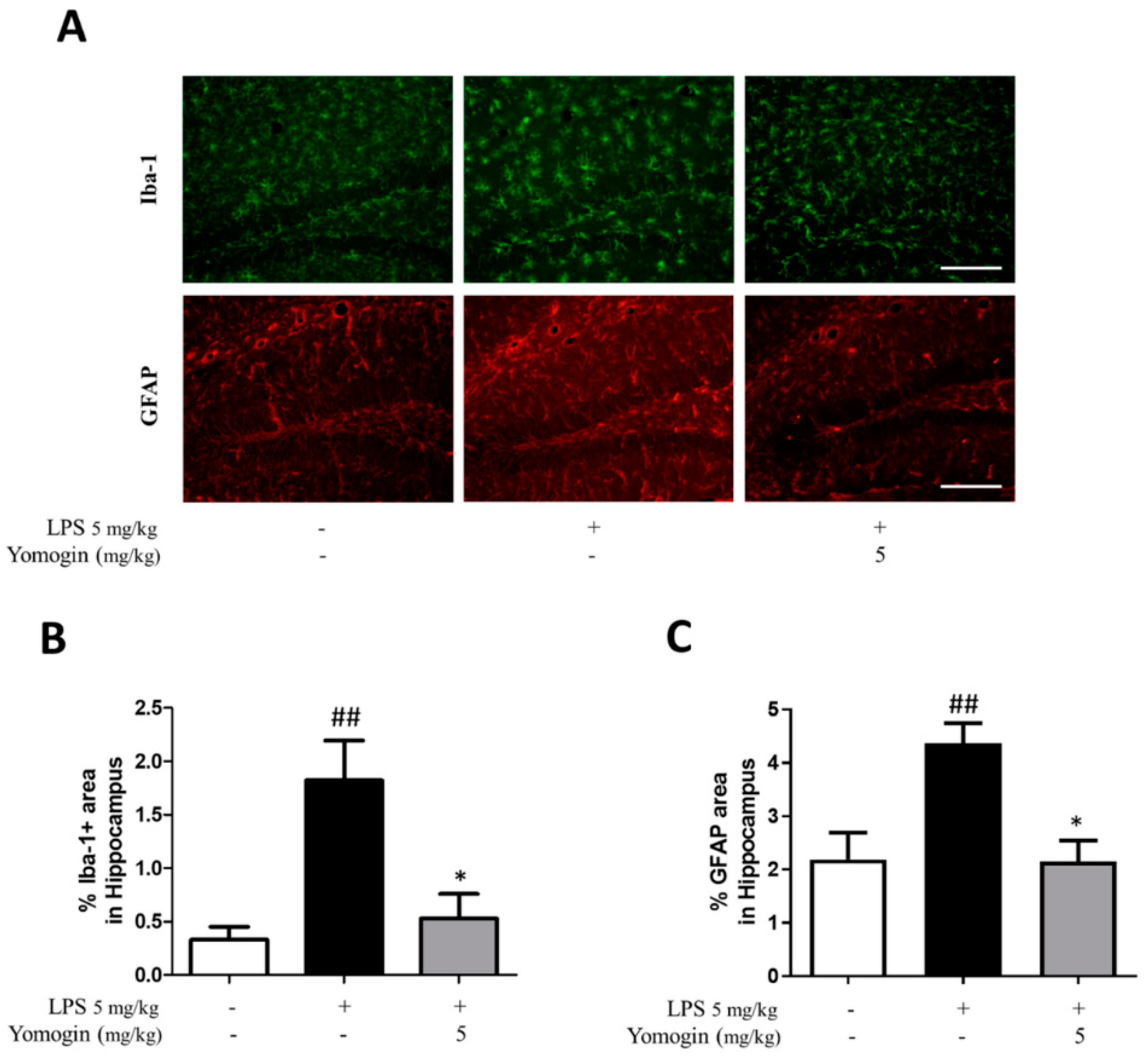
3.6. Effects of Yomogin on Microglia and Astrocyte Activation in LPS-Treated Mouse Brain
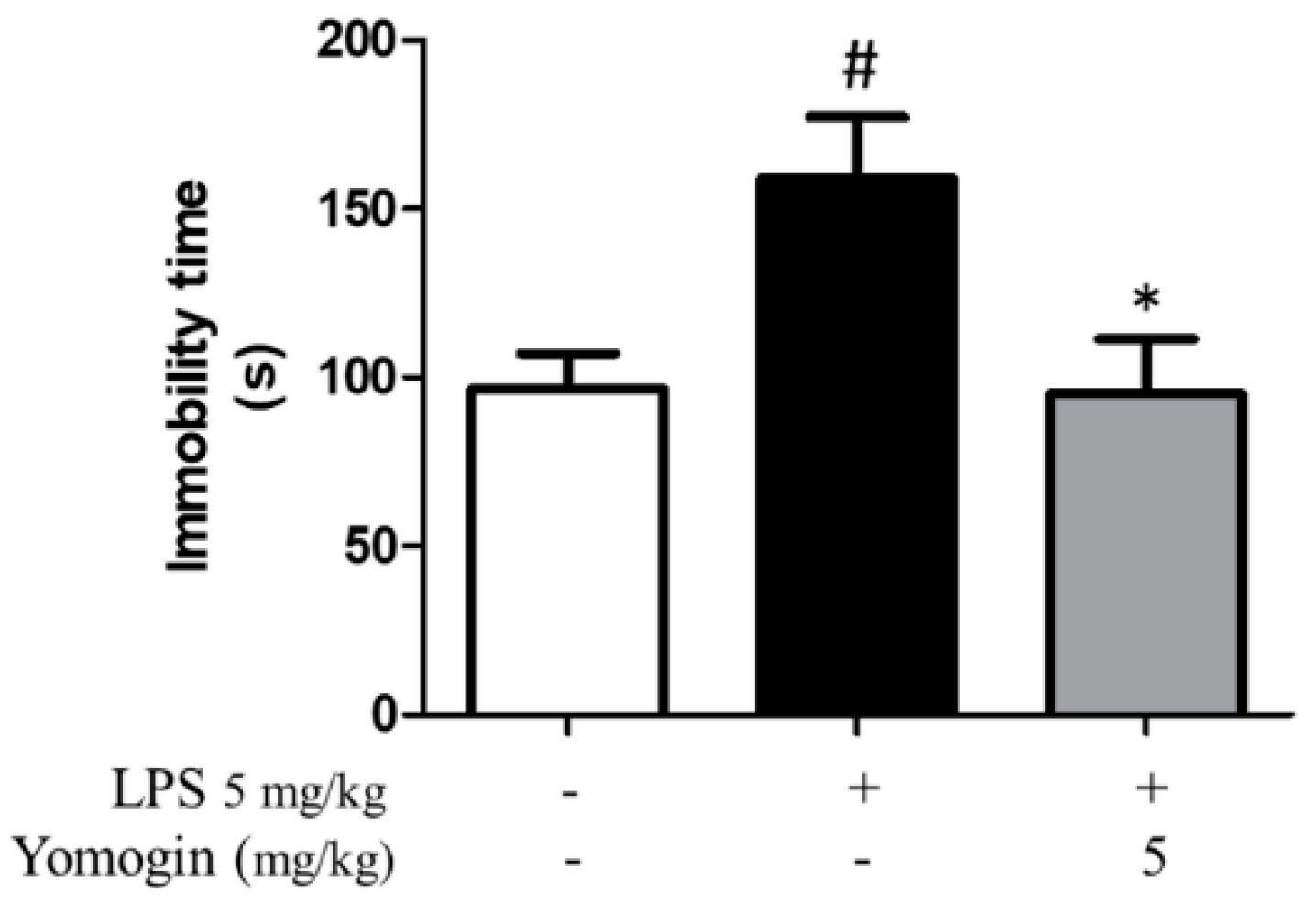
3.7. Effects of Yomogin on Depressive Behavior in LPS-Treated Mouse
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ju, I.G.; Choi, J.G.; Kim, N.; Kwak, C.; Lee, J.K.; Oh, M.S. Peucedani Japonici Radix ameliorates lipopolysaccharide-induced neuroinflammation by regulating microglial responses. Neurosci. Lett. 2018, 686, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Bachiller, S.; Jimenez-Ferrer, I.; Paulus, A.; Yang, Y.; Swanberg, M.; Deierborg, T.; Boza-Serrano, A. Microglia in Neurological Diseases: A Road Map to Brain-Disease Dependent-Inflammatory Response. Front. Cell. Neurosci. 2018, 12, 488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lively, S.; Schlichter, L.C. Microglia Responses to Pro-inflammatory Stimuli (LPS, IFNgamma+TNFalpha) and Reprogramming by Resolving Cytokines (IL-4, IL-10). Front. Cell. Neurosci. 2018, 12, 215. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhong, L.; Yu, Z.; Qi, J. Anti-neuroinflammatory effects of tannic acid against lipopolysaccharide-induced BV2 microglial cells via inhibition of NF-kappaB activation. Drug Dev. Res. 2019, 80, 262–268. [Google Scholar] [CrossRef]
- Degan, D.; Ornello, R.; Tiseo, C.; Carolei, A.; Sacco, S.; Pistoia, F. The Role of Inflammation in Neurological Disorders. Curr. Pharm. Des. 2018, 24, 1485–1501. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Giuliani, F. The Role of Inflammation in Depression and Fatigue. Front. Immunol. 2019, 10, 1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzman-Martinez, L.; Maccioni, R.B.; Andrade, V.; Navarrete, L.P.; Pastor, M.G.; Ramos-Escobar, N. Neuroinflammation as a Common Feature of Neurodegenerative Disorders. Front. Pharmacol. 2019, 10, 1008. [Google Scholar] [CrossRef] [Green Version]
- Heneka, M.T.; Carson, M.J.; El Khoury, J.; Landreth, G.E.; Brosseron, F.; Feinstein, D.L.; Jacobs, A.H.; Wyss-Coray, T.; Vitorica, J.; Ransohoff, R.M.; et al. Neuroinflammation in Alzheimer’s disease. Lancet Neurol. 2015, 14, 388–405. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.W.; Zhang, X.; Huang, W.J. Role of neuroinflammation in neurodegenerative diseases (Review). Mol. Med. Rep. 2016, 13, 3391–3396. [Google Scholar] [CrossRef] [Green Version]
- Shal, B.; Ding, W.; Ali, H.; Kim, Y.S.; Khan, S. Anti-neuroinflammatory Potential of Natural Products in Attenuation of Alzheimer’s Disease. Front. Pharmacol. 2018, 9, 548. [Google Scholar] [CrossRef]
- Liu, Y.M.; Niu, L.; Wang, L.L.; Bai, L.; Fang, X.Y.; Li, Y.C.; Yi, L.T. Berberine attenuates depressive-like behaviors by suppressing neuro-inflammation in stressed mice. Brain Res. Bull. 2017, 134, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Thingore, C.; Kshirsagar, V.; Juvekar, A. Amelioration of oxidative stress and neuroinflammation in lipopolysaccharide-induced memory impairment using Rosmarinic acid in mice. Metab. Brain Dis. 2020, 36, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Ali, T.; Park, H.Y.; Badshah, H.; Rehman, S.U.; Kim, M.O. Neuroprotective Effect of Fisetin Against Amyloid-Beta-Induced Cognitive/Synaptic Dysfunction, Neuroinflammation, and Neurodegeneration in Adult Mice. Mol. Neurobiol. 2017, 54, 2269–2285. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kim, T.S. Synergistic induction of 1,25-dihydroxyvitamin D(3)- and all-trans-retinoic acid-induced differentiation of HL-60 leukemia cells by yomogin, a sesquiterpene lactone from Artemisia princeps. Planta Med. 2002, 68, 886–890. [Google Scholar] [CrossRef]
- Tigno, X.T.; Gumila, E. In vivo microvascular actions of Artemisia vulgaris L. in a model of ischemia-reperfusion injury in the rat intestinal mesentery. Clin. Hemorheol. Microcirc. 2000, 23, 159–165. [Google Scholar]
- Jeong, S.H.; Koo, S.J.; Ha, J.H.; Ryu, S.Y.; Park, H.J.; Lee, K.T. Induction of apoptosis by yomogin in human promyelocytic leukemic HL-60 cells. Biol. Pharm. Bull. 2004, 27, 1106–1111. [Google Scholar] [CrossRef] [Green Version]
- Ryu, S.Y.; Oak, M.H.; Kim, K.M. Yomogin inhibits the degranulation of mast cells and the production of the nitric oxide in activated RAW 264.7 cells. Planta Med. 2000, 66, 171–173. [Google Scholar] [CrossRef]
- Ryu, J.H.; Lee, H.J.; Jeong, Y.S.; Ryu, S.Y.; Han, Y.N. Yomogin, an inhibitor of nitric oxide production in LPS-activated macrophages. Arch. Pharmacal Res. 1998, 21, 481–484. [Google Scholar] [CrossRef]
- Son, S.R.; Ju, I.G.; Kim, J.; Park, K.T.; Oh, M.S.; Jang, D.S. Chemical Constituents from the Aerial Parts of Artemisia iwayomogi and Their Anti-Neuroinflammatory Activities. Plants 2022, 11, 1954. [Google Scholar] [CrossRef]
- Jakupovic, J.; Schuster, A.; Bohlmann, F.; Dillon, M. Lumiyomogin, ferreyrantholide, fruticolide and other sesquiterpene lactones from Ferreyranthus fruticosus. Phytochemistry 1988, 27, 1113–1120. [Google Scholar] [CrossRef]
- Coleman, J.W. Nitric oxide in immunity and inflammation. Int. Immunopharmacol. 2001, 1, 1397–1406. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Martinez, C.C.; Jang, D.S.; Lee, J.K.; Oh, M.S. Anti-neuroinflammatory effect of Iresine celosia on lipopolysaccharide-stimulated microglial cells and mouse. Biomed. Pharm. 2019, 111, 1359–1366. [Google Scholar] [CrossRef]
- Troubat, R.; Barone, P.; Leman, S.; Desmidt, T.; Cressant, A.; Atanasova, B.; Brizard, B.; El Hage, W.; Surget, A.; Belzung, C.; et al. Neuroinflammation and depression: A review. Eur. J. Neurosci. 2020, 53, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Batista, C.R.A.; Gomes, G.F.; Candelario-Jalil, E.; Fiebich, B.L.; de Oliveira, A.C.P. Lipopolysaccharide-Induced Neuroinflammation as a Bridge to Understand Neurodegeneration. Int. J. Mol. Sci. 2019, 20, 2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiebich, B.L.; Batista, C.R.A.; Saliba, S.W.; Yousif, N.M.; de Oliveira, A.C.P. Role of Microglia TLRs in Neurodegeneration. Front. Cell. Neurosci. 2018, 12, 329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, W.; Bi, D.; Zheng, R.; Cai, N.; Xu, H.; Zhou, R.; Lu, J.; Wan, M.; Xu, X. Identification and activation of TLR4-mediated signalling pathways by alginate-derived guluronate oligosaccharide in RAW264.7 macrophages. Sci. Rep. 2017, 7, 1663. [Google Scholar] [CrossRef] [Green Version]
- Duman, C.H.; Schlesinger, L.; Kodama, M.; Russell, D.S.; Duman, R.S. A role for MAP kinase signaling in behavioral models of depression and antidepressant treatment. Biol. Psychiatry 2007, 61, 661–670. [Google Scholar] [CrossRef]
- Zhao, Y.W.; Pan, Y.Q.; Tang, M.M.; Lin, W.J. Blocking p38 Signaling Reduces the Activation of Pro-inflammatory Cytokines and the Phosphorylation of p38 in the Habenula and Reverses Depressive-Like Behaviors Induced by Neuroinflammation. Front. Pharmacol. 2018, 9, 511. [Google Scholar] [CrossRef]
- El Rawas, R.; Amaral, I.M.; Hofer, A. Is p38 MAPK Associated to Drugs of Abuse-Induced Abnormal Behaviors? Int. J. Mol. Sci. 2020, 21, 4833. [Google Scholar] [CrossRef]
- Hollos, P.; Marchisella, F.; Coffey, E.T. JNK Regulation of Depression and Anxiety. Brain Plast. 2018, 3, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Mohammad, H.; Marchisella, F.; Ortega-Martinez, S.; Hollos, P.; Eerola, K.; Komulainen, E.; Kulesskaya, N.; Freemantle, E.; Fagerholm, V.; Savontaus, E.; et al. JNK1 controls adult hippocampal neurogenesis and imposes cell-autonomous control of anxiety behaviour from the neurogenic niche. Mol. Psychiatry 2018, 23, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lin, W.; Tang, M.; Zhao, Y.; Zhang, K.; Wang, X.; Li, Y. Inhibition of JNK ameliorates depressive-like behaviors and reduces the activation of pro-inflammatory cytokines and the phosphorylation of glucocorticoid receptors at serine 246 induced by neuroinflammation. Psychoneuroendocrinology 2020, 113, 104580. [Google Scholar] [CrossRef] [PubMed]
- Catorce, M.N.; Gevorkian, G. LPS-induced Murine Neuroinflammation Model: Main Features and Suitability for Pre-clinical Assessment of Nutraceuticals. Curr. Neuropharmacol. 2016, 14, 155–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Jalabi, W.; Shpargel, K.B.; Farabaugh, K.T.; Dutta, R.; Yin, X.; Kidd, G.J.; Bergmann, C.C.; Stohlman, S.A.; Trapp, B.D. Lipopolysaccharide-induced microglial activation and neuroprotection against experimental brain injury is independent of hematogenous TLR4. J. Neurosci. 2012, 32, 11706–11715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felger, J.C.; Lotrich, F.E. Inflammatory cytokines in depression: Neurobiological mechanisms and therapeutic implications. Neuroscience 2013, 246, 199–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.T.; Howell, J.C.; Ozturk, T.; Gangishetti, U.; Kollhoff, A.L.; Hatcher-Martin, J.M.; Anderson, A.M.; Tyor, W.R. CSF Cytokines in Aging, Multiple Sclerosis, and Dementia. Front. Immunol. 2019, 10, 480. [Google Scholar] [CrossRef] [Green Version]
- Vogelzangs, N.; de Jonge, P.; Smit, J.H.; Bahn, S.; Penninx, B.W. Cytokine production capacity in depression and anxiety. Transl. Psychiatry 2016, 6, e825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banks, W.A. Characteristics of compounds that cross the blood-brain barrier. BMC Neurol. 2009, 9 (Suppl. S1), S3. [Google Scholar] [CrossRef] [Green Version]
- Ivanescu, B.; Miron, A.; Corciova, A. Sesquiterpene Lactones from Artemisia Genus: Biological Activities and Methods of Analysis. J. Anal. Methods Chem. 2015, 2015, 247685. [Google Scholar] [CrossRef] [Green Version]
- Haruwaka, K.; Ikegami, A.; Tachibana, Y.; Ohno, N.; Konishi, H.; Hashimoto, A.; Matsumoto, M.; Kato, D.; Ono, R.; Kiyama, H.; et al. Dual microglia effects on blood brain barrier permeability induced by systemic inflammation. Nat. Commun. 2019, 10, 5816. [Google Scholar] [CrossRef] [Green Version]
- Galea, I. The blood-brain barrier in systemic infection and inflammation. Cell. Mol. Immunol. 2021, 18, 2489–2501. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Primer Sequence | Accession Number |
|---|---|---|
| iNOS | 5′-GTG TTC TTT GCT TCC ATG CT-3′ 5′- AGT TGC TCC TCT TCC AAG GT-3′ | NM_001313922.1 |
| COX-2 | 5′-TGG GGT GAT GAG CAA CTA TT-3′ 5′- AAG GAG CTC TGG GTC AAA CT-3′ | NM_011198.4 |
| TNF-α | 5′-GAT TAT GGC TCA GGG TCC AA-3′ 5′- GCT CCA GTG AAT TCG GAA AG-3′ | NM_001278601.1 |
| IL-6 | 5′-CCG GAG AGG AGA CTT CAC AG-3′ 5′- TTG CCA TTG CAC AAC TCT TT-3′ | NM_001314054.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Ju, I.G.; Kim, N.; Huh, E.; Son, S.-R.; Hong, J.P.; Choi, Y.; Jang, D.S.; Oh, M.S. Yomogin, Isolated from Artemisia iwayomogi, Inhibits Neuroinflammation Stimulated by Lipopolysaccharide via Regulating MAPK Pathway. Antioxidants 2023, 12, 106. https://doi.org/10.3390/antiox12010106
Kim JH, Ju IG, Kim N, Huh E, Son S-R, Hong JP, Choi Y, Jang DS, Oh MS. Yomogin, Isolated from Artemisia iwayomogi, Inhibits Neuroinflammation Stimulated by Lipopolysaccharide via Regulating MAPK Pathway. Antioxidants. 2023; 12(1):106. https://doi.org/10.3390/antiox12010106
Chicago/Turabian StyleKim, Jin Hee, In Gyoung Ju, Namkwon Kim, Eugene Huh, So-Ri Son, Joon Pyo Hong, Yujin Choi, Dae Sik Jang, and Myung Sook Oh. 2023. "Yomogin, Isolated from Artemisia iwayomogi, Inhibits Neuroinflammation Stimulated by Lipopolysaccharide via Regulating MAPK Pathway" Antioxidants 12, no. 1: 106. https://doi.org/10.3390/antiox12010106
APA StyleKim, J. H., Ju, I. G., Kim, N., Huh, E., Son, S.-R., Hong, J. P., Choi, Y., Jang, D. S., & Oh, M. S. (2023). Yomogin, Isolated from Artemisia iwayomogi, Inhibits Neuroinflammation Stimulated by Lipopolysaccharide via Regulating MAPK Pathway. Antioxidants, 12(1), 106. https://doi.org/10.3390/antiox12010106