
Bilirubin Levels Are Negatively Correlated with Adiposity in Obese Men and Women, and Its Catabolized Product, Urobilin, Is Positively Associated with Insulin Resistance

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Subject Participation
2.2. Plasma Bilirubin Measurements
2.3. Plasma Urobilin Measurements
2.4. Statistical Analysis
3. Results
3.1. Bilirubin and Urobilin Levels in Obese and Lean Men and Women
Phenotyping of Obese and Lean Men and Women
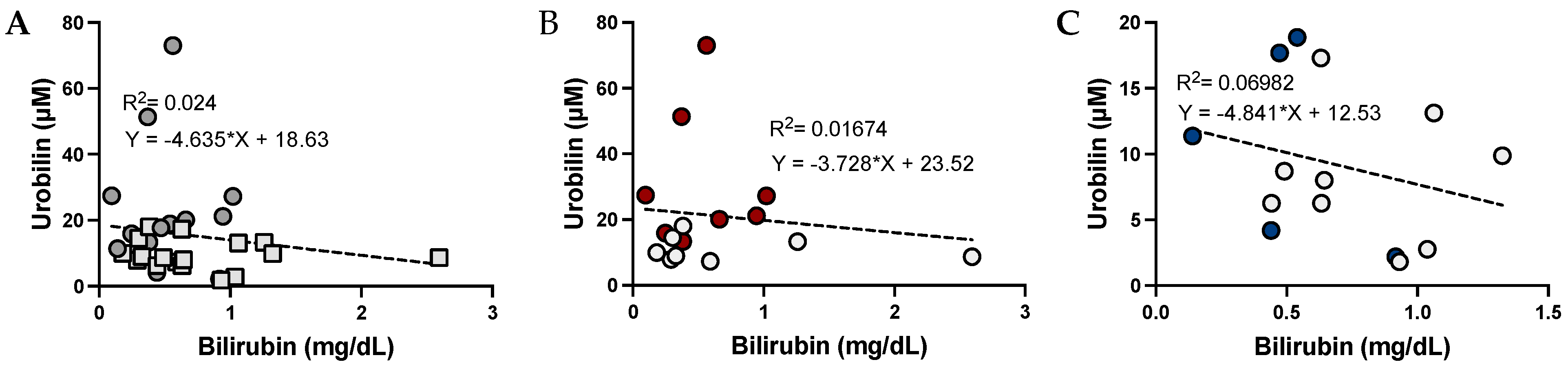
3.2. The Relationship between Urobilin and Bilirubin Levels
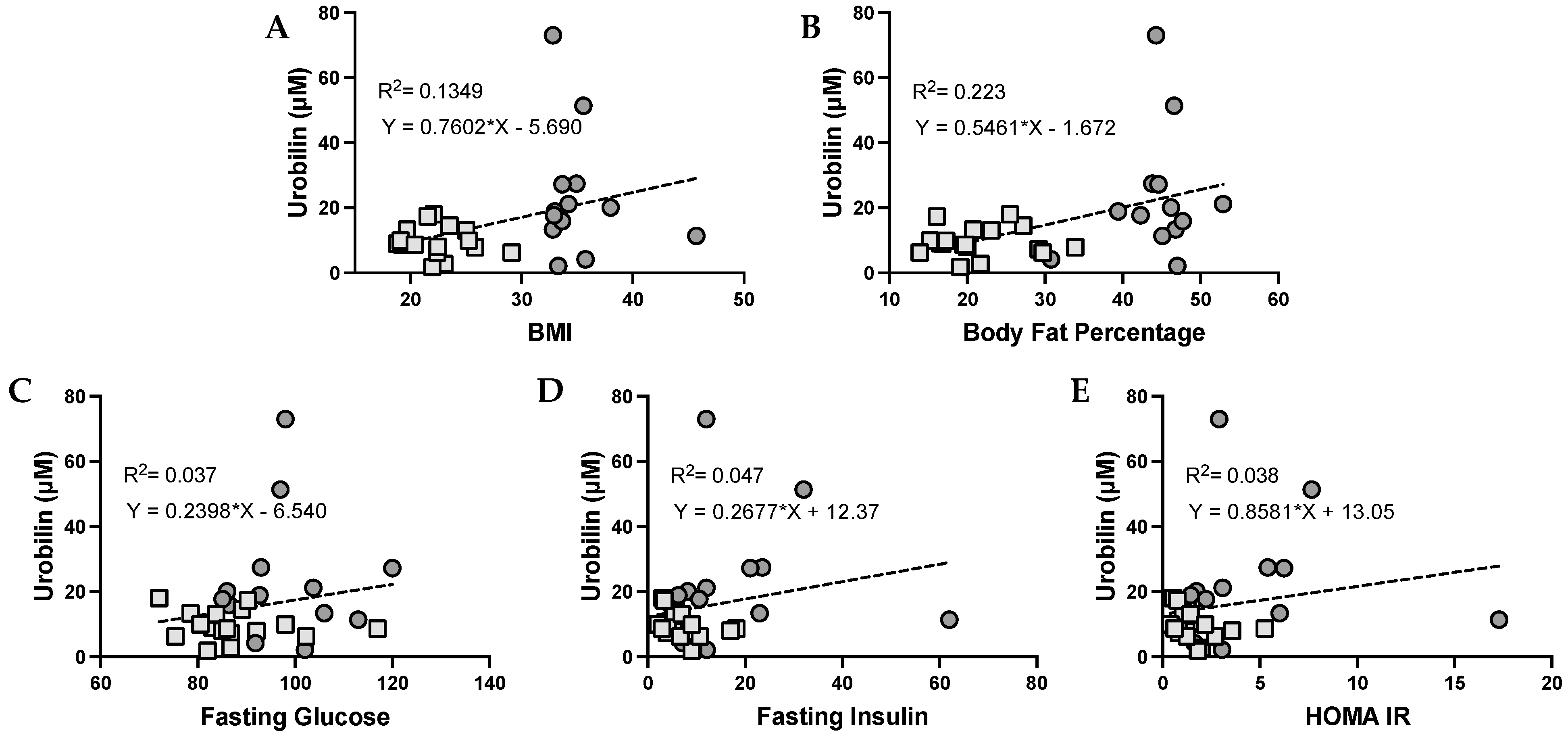
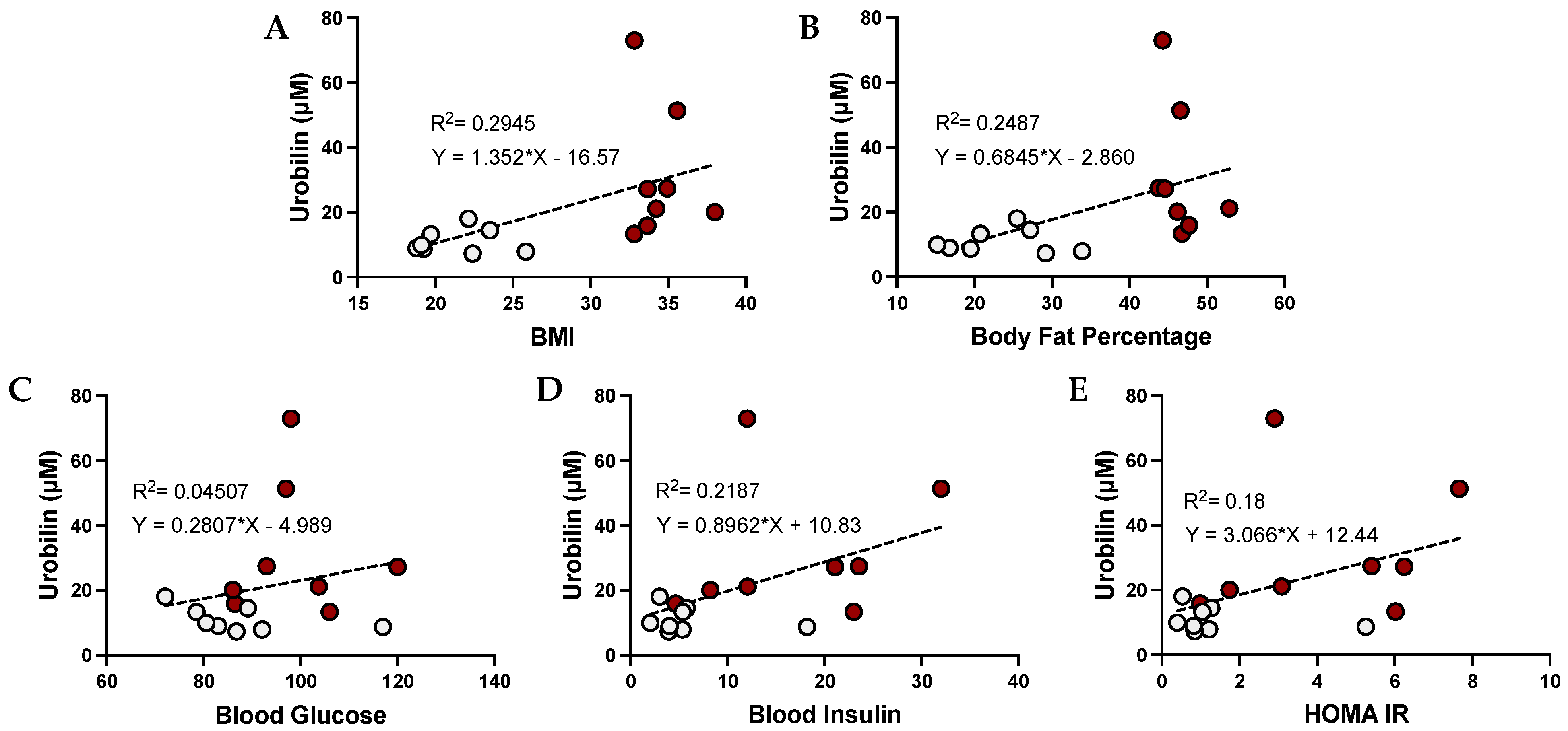
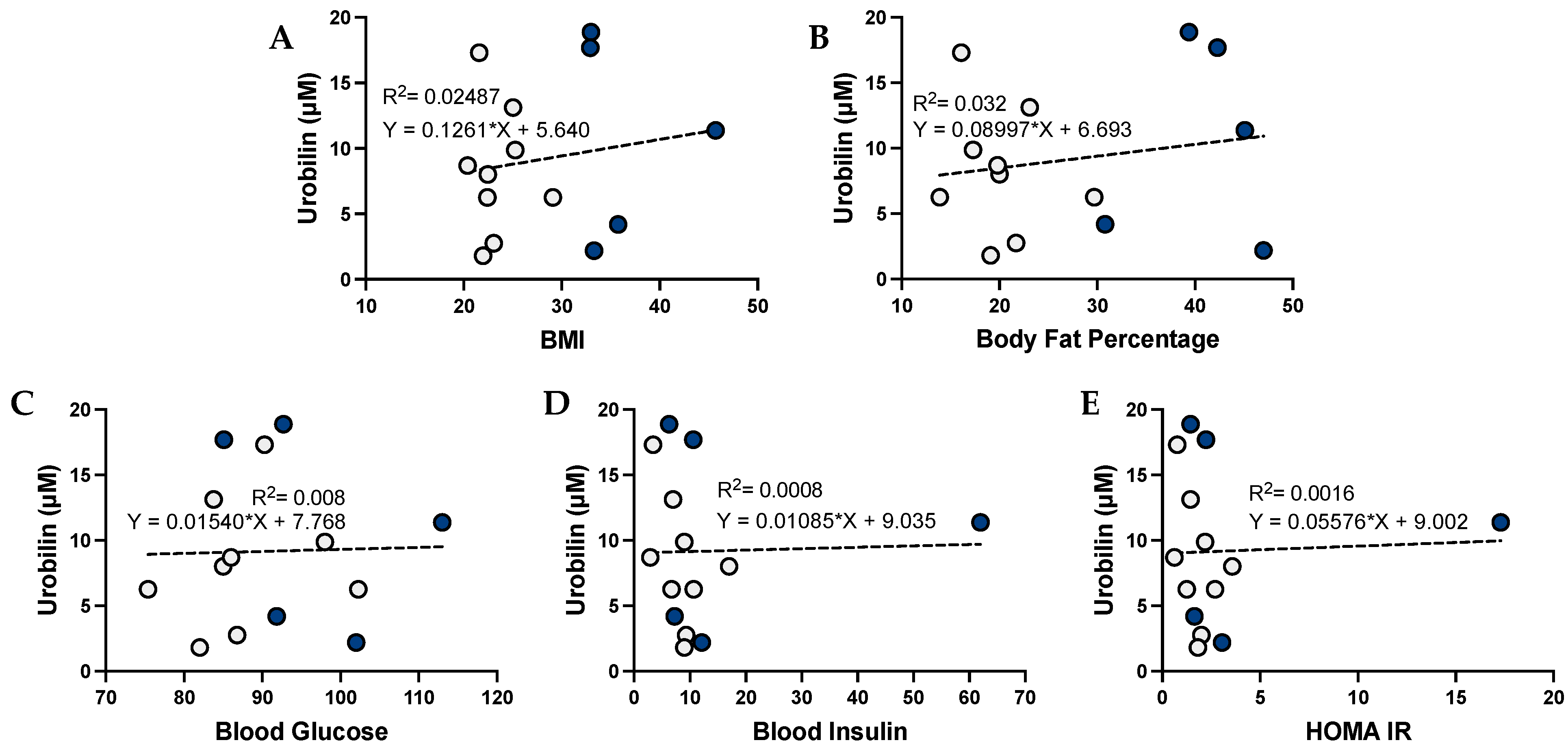
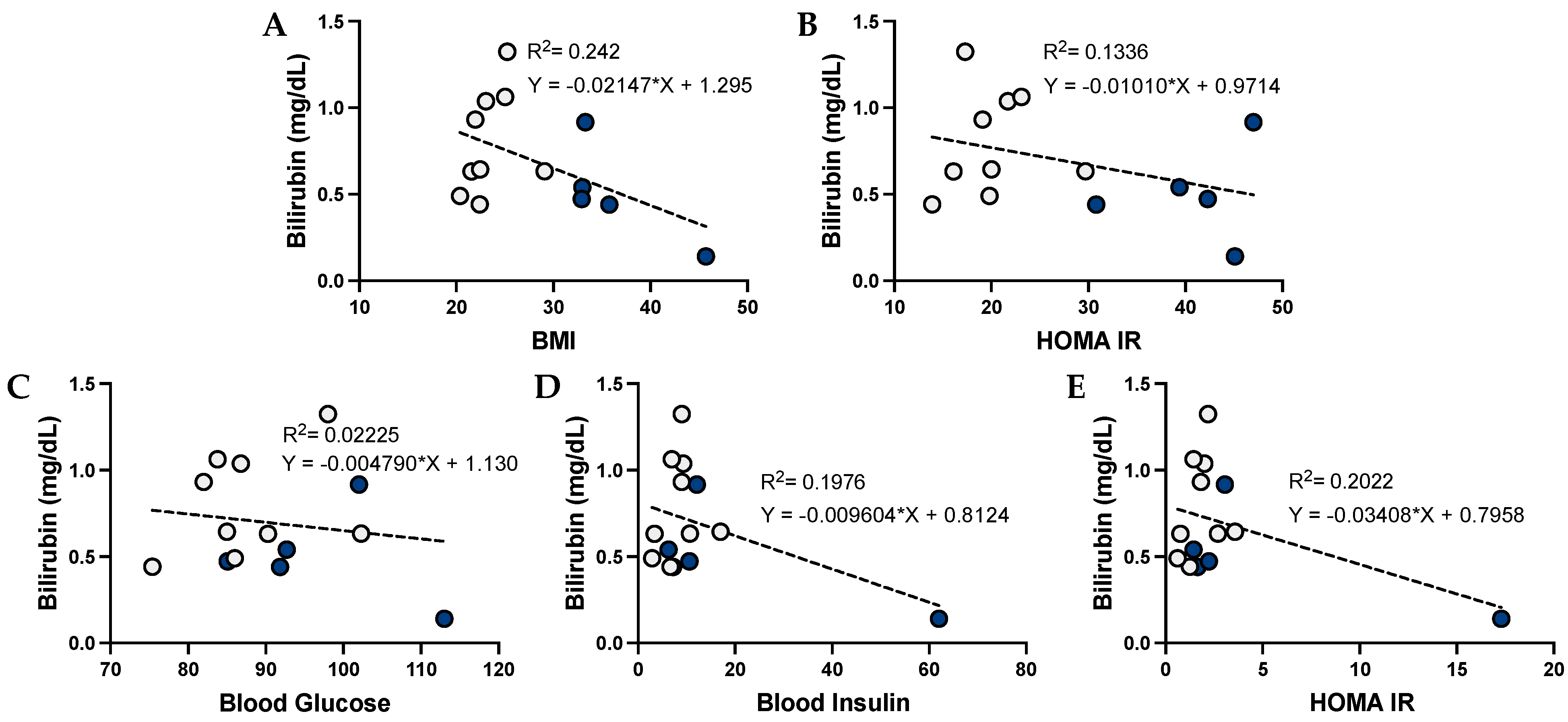
3.3. Urobilin and Its Correlation with Metabolic Dysfunction in Humans
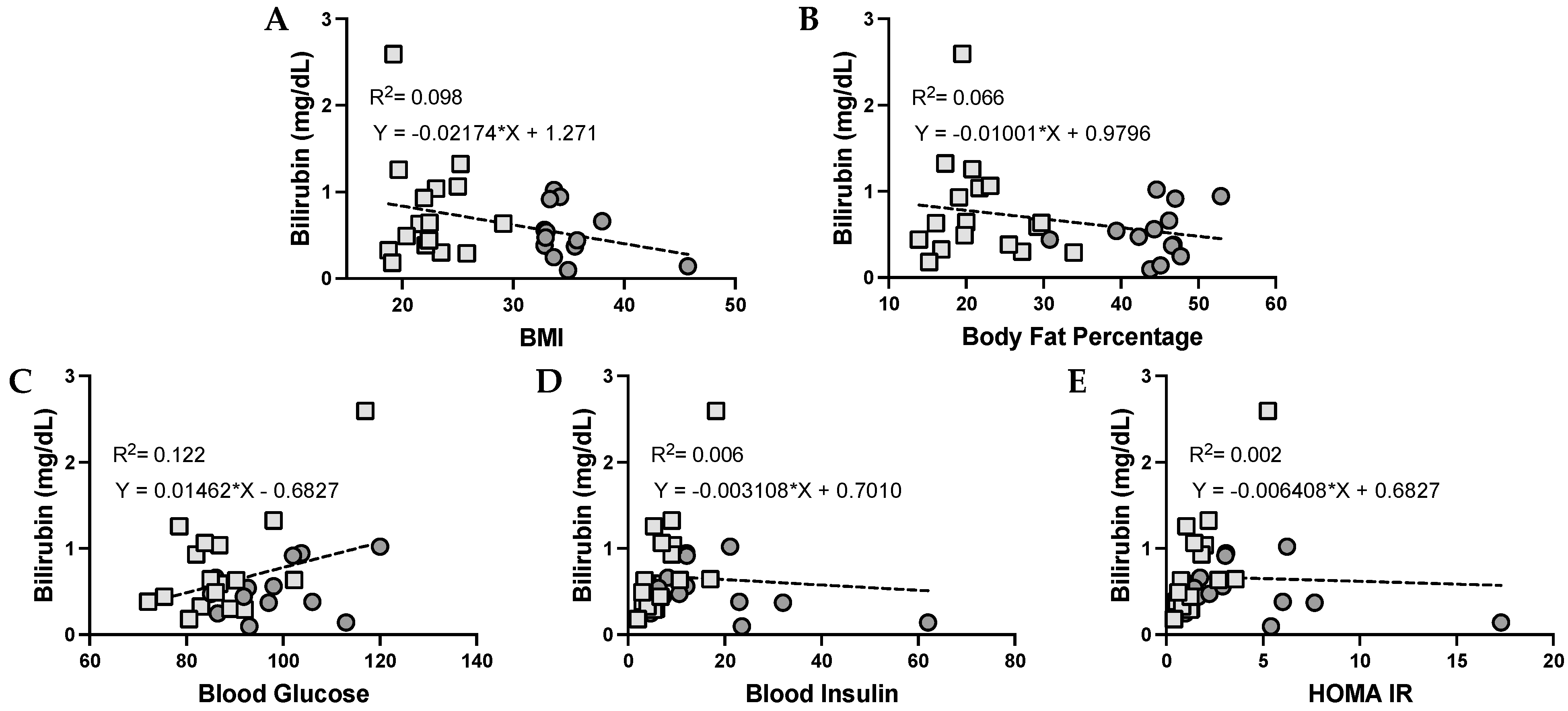
3.4. Bilirubin and Its Correlation with Metabolic Dysfunction in Humans
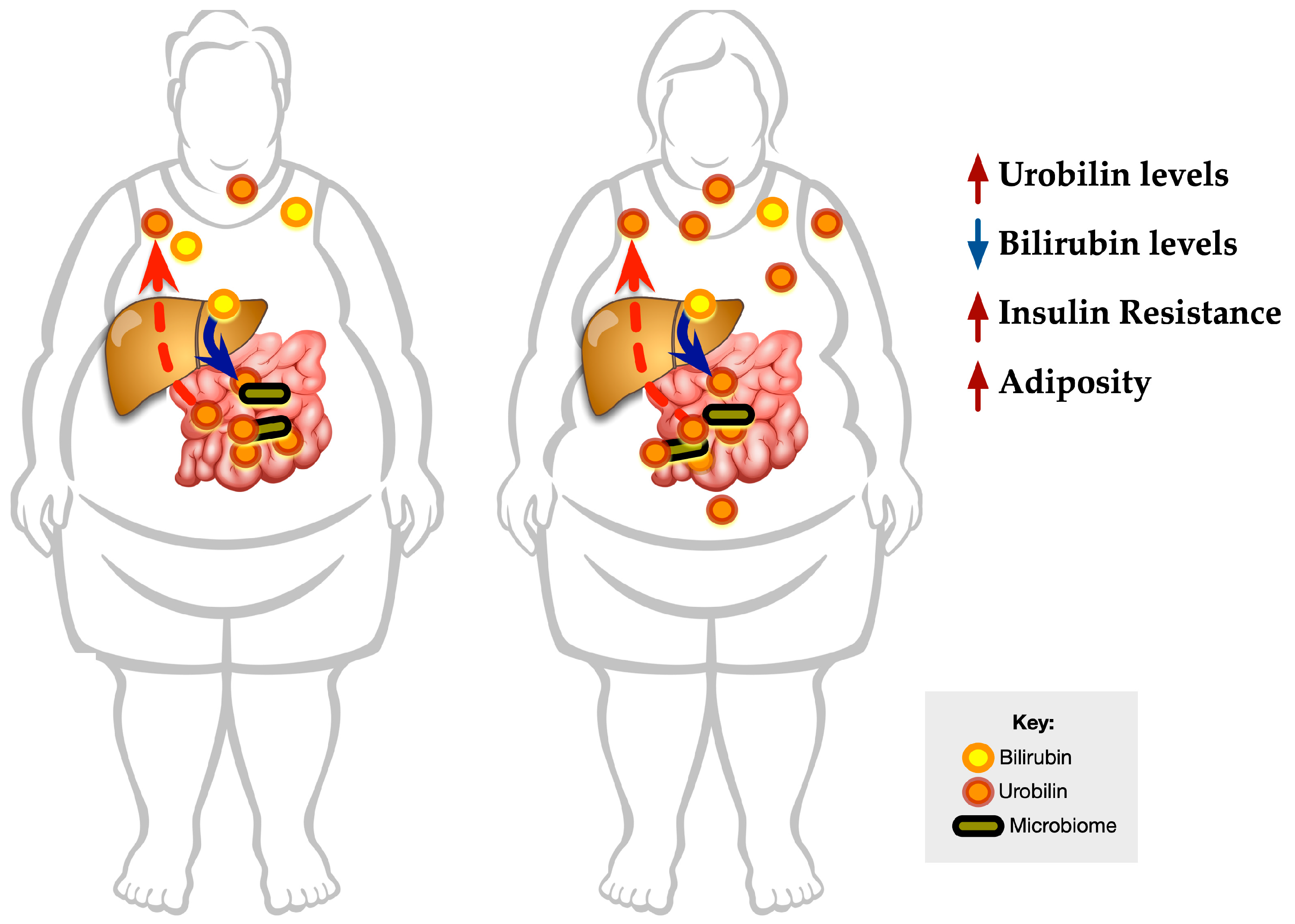
3.5. Graphical Representation of the Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weir, C.B.; Jan, A. BMI Classification Percentile and Cut Off Points; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Creeden, J.F.; Gordon, D.M.; Stec, D.E.; Hinds, T.D., Jr. Bilirubin as a metabolic hormone: The physiological relevance of low levels. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E191–E207. [Google Scholar] [CrossRef] [PubMed]
- Hinds, T.D., Jr.; Stec, D.E. Bilirubin Safeguards Cardiorenal and Metabolic Diseases: A Protective Role in Health. Curr. Hypertens. Rep. 2019, 21, 87. [Google Scholar] [CrossRef] [PubMed]
- Weaver, L.; Hamoud, A.R.; Stec, D.E.; Hinds, T.D., Jr. Biliverdin reductase and bilirubin in hepatic disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 314, G668–G676. [Google Scholar] [CrossRef]
- Hinds, T.D., Jr.; Stec, D.E. Bilirubin, a Cardiometabolic Signaling Molecule. Hypertension 2018, 72, 788–795. [Google Scholar] [CrossRef]
- Hamoud, A.R.; Weaver, L.; Stec, D.E.; Hinds, T.D., Jr. Bilirubin in the Liver-Gut Signaling Axis. Trends Endocrinol. Metab. 2018, 29, 140–150. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, L.; Hosick, P.A.; John, K.; Stec, D.E.; Hinds, T.D., Jr. Biliverdin reductase isozymes in metabolism. Trends Endocrinol. Metab. 2015, 26, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Jansen, T.; Daiber, A. Direct Antioxidant Properties of Bilirubin and Biliverdin. Is there a Role for Biliverdin Reductase? Front. Pharm. 2012, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Stocker, R.; Yamamoto, Y.; McDonagh, A.F.; Glazer, A.N.; Ames, B.N. Bilirubin is an antioxidant of possible physiological importance. Science 1987, 235, 1043–1046. [Google Scholar] [CrossRef]
- Stocker, R.; Peterhans, E. Antioxidant properties of conjugated bilirubin and biliverdin: Biologically relevant scavenging of hypochlorous acid. Free Radic. Res. Commun. 1989, 6, 57–66. [Google Scholar] [CrossRef]
- Li, Y.; Huang, B.; Ye, T.; Wang, Y.; Xia, D.; Qian, J. Physiological concentrations of bilirubin control inflammatory response by inhibiting NF-κB and inflammasome activation. Int. Immunopharmacol. 2020, 84, 106520. [Google Scholar] [CrossRef]
- Chen, W.; Tumanov, S.; Fazakerley, D.J.; Cantley, J.; James, D.E.; Dunn, L.L.; Shaik, T.; Suarna, C.; Stocker, R. Bilirubin deficiency renders mice susceptible to hepatic steatosis in the absence of insulin resistance. Redox Biol. 2021, 47, 102152. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.M.; Hong, S.H.; Kipp, Z.A.; Hinds, T.D., Jr. Identification of Binding Regions of Bilirubin in the Ligand-Binding Pocket of the Peroxisome Proliferator-Activated Receptor-A (PPARalpha). Molecules 2021, 26, 2975. [Google Scholar] [CrossRef] [PubMed]
- Hinds, T.D., Jr.; Creeden, J.F.; Gordon, D.M.; Stec, D.F.; Donald, M.C.; Stec, D.E. Bilirubin Nanoparticles Reduce Diet-Induced Hepatic Steatosis, Improve Fat Utilization, and Increase Plasma beta-Hydroxybutyrate. Front. Pharm. 2020, 11, 594574. [Google Scholar] [CrossRef]
- Gordon, D.M.; Neifer, K.L.; Hamoud, A.A.; Hawk, C.F.; Nestor-Kalinoski, A.L.; Miruzzi, S.A.; Morran, M.P.; Adeosun, S.O.; Sarver, J.G.; Erhardt, P.W.; et al. Bilirubin remodels murine white adipose tissue by reshaping mitochondrial activity and the coregulator profile of peroxisome proliferator-activated receptor alpha. J. Biol. Chem. 2020, 295, 9804–9822. [Google Scholar] [CrossRef]
- Gordon, D.M.; Blomquist, T.M.; Miruzzi, S.A.; McCullumsmith, R.; Stec, D.E.; Hinds, T.D., Jr. RNA sequencing in human HepG2 hepatocytes reveals PPAR-alpha mediates transcriptome responsiveness of bilirubin. Physiol. Genom. 2019, 51, 234–240. [Google Scholar] [CrossRef]
- Stec, D.E.; John, K.; Trabbic, C.J.; Luniwal, A.; Hankins, M.W.; Baum, J.; Hinds, T.D., Jr. Bilirubin Binding to PPARalpha Inhibits Lipid Accumulation. PLoS ONE 2016, 11, e0153427. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Shao, X.; Bao, Y.; Zhu, J.; Chen, L.; Zhang, L.; Ma, X.; Zhong, X.B. Impact of obese levels on the hepatic expression of nuclear receptors and drug-metabolizing enzymes in adult and offspring mice. Acta Pharm. Sin. B 2020, 10, 171–185. [Google Scholar] [CrossRef]
- Xu, J.; Kulkarni, S.R.; Li, L.; Slitt, A.L. UDP-glucuronosyltransferase expression in mouse liver is increased in obesity- and fasting-induced steatosis. Drug Metab. Dispos. Biol. Fate Chem. 2012, 40, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Sundararaghavan, V.L.; Sindhwani, P.; Hinds, T.D., Jr. Glucuronidation and UGT isozymes in bladder: New targets for the treatment of uroepithelial carcinomas? Oncotarget 2017, 8, 3640–3648. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.H.; Kim, M.; Kim, M.; Kang, M.; Yoo, H.J.; Lee, N.H.; Kim, Y.H.; Song, M.; Lee, J.H. Metabolites distinguishing visceral fat obesity and atherogenic traits in individuals with overweight. Obesity 2017, 25, 323–331. [Google Scholar] [CrossRef]
- Stenemo, M.; Ganna, A.; Salihovic, S.; Nowak, C.; Sundstrom, J.; Giedraitis, V.; Broeckling, C.D.; Prenni, J.E.; Svensson, P.; Magnusson, P.K.E.; et al. The metabolites urobilin and sphingomyelin (30:1) are associated with incident heart failure in the general population. ESC Heart Fail. 2019, 6, 764–773. [Google Scholar] [CrossRef] [Green Version]
- Ottosson, F.; Smith, E.; Fernandez, C.; Melander, O. Plasma Metabolites Associate with All-Cause Mortality in Individuals with Type 2 Diabetes. Metabolites 2020, 10, 315. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Culliton, K.; Louati, H.; Laneuville, O.; Ramsay, T.; Trudel, G. Six degrees head-down tilt bed rest caused low-grade hemolysis: A prospective randomized clinical trial. npj Microgravity 2021, 7, 4. [Google Scholar] [CrossRef]
- Kotal, P.; Fevery, J. Quantitation of urobilinogen in feces, urine, bile and serum by direct spectrophotometry of zinc complex. Clin. Chim. Acta 1991, 202, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Trudel, G.; Uhthoff, H.K.; Laneuville, O. Hemolysis during and after 21 days of head--down--tilt bed rest. Physiol. Rep. 2017, 5, e13469. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.; Pfitzner, B.; Neschen, S.; Kahle, M.; Harir, M.; Lucio, M.; Moritz, F.; Tziotis, D.; Witting, M.; Rothballer, M.; et al. Distinct signatures of host-microbial meta-metabolome and gut microbiome in two C57BL/6 strains under high-fat diet. ISME J. 2014, 8, 2380–2396. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Jia, H.; Li, J.; Yu, M.; Yang, Y.; Tian, D.; Zhang, H.; Zou, Z. Cecal Gut Microbiota and Metabolites Might Contribute to the Severity of Acute Myocardial Ischemia by Impacting the Intestinal Permeability, Oxidative Stress, and Energy Metabolism. Front. Microbiol. 2019, 10, 1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torgerson, J.S.; Lindroos, A.K.; Sjostrom, C.D.; Olsson, R.; Lissner, L.; Sjostrom, L. Are elevated aminotransferases and decreased bilirubin additional characteristics of the metabolic syndrome? Obes. Res. 1997, 5, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Hinds, T.D., Jr.; Creeden, J.F.; Gordon, D.M.; Spegele, A.C.; Britton, S.L.; Koch, L.G.; Stec, D.E. Rats Genetically Selected for High Aerobic Exercise Capacity Have Elevated Plasma Bilirubin by Upregulation of Hepatic Biliverdin Reductase-A (BVRA) and Suppression of UGT1A1. Antioxidants 2020, 9, 889. [Google Scholar] [CrossRef] [PubMed]
- Stec, D.E.; Hinds, T.D., Jr. Natural Product Heme Oxygenase Inducers as Treatment for Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2020, 21, 9493. [Google Scholar] [CrossRef] [PubMed]
- Woronyczova, J.; Novákova, M.; Lenicek, M.; Batovsky, M.; Bolek, E.; Cifkova, R.; Vitek, L. Serum bilirubin concentrations and the prevalence of Gilbert syndrome in elite athletes. Sport. Med. Open Access 2022, 8, 84. [Google Scholar] [CrossRef]
- Swift, D.L.; Johannsen, N.M.; Earnest, C.P.; Blair, S.N.; Church, T.S. Effect of different doses of aerobic exercise training on total bilirubin levels. Med. Sci. Sport. Exerc. 2012, 44, 569–574. [Google Scholar] [CrossRef] [Green Version]
- Loprinzi, P.D.; Abbott, K. Physical activity and total serum bilirubin levels among insulin sensitive and insulin resistant U.S. adults. J. Diabetes Metab. Disord. 2014, 13, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, D.T.; DelCimmuto, N.R.; Flack, K.D.; Stec, D.E.; Hinds, T.D., Jr. Reactive Oxygen Species (ROS) and Antioxidants as Immunomodulators in Exercise: Implications for Heme Oxygenase and Bilirubin. Antioxidants 2022, 11, 179. [Google Scholar] [CrossRef]
- Flack, K.D.; Vítek, L.; Fry, C.; Stec, D.E.; Hinds, T.D., Jr. Cutting Edge Concepts: Does Bilirubin Enhance Exercise Performance? Front. Sport. Act. Living 2022, 4, 509. [Google Scholar] [CrossRef]
- Nath, K.A.; d’Uscio, L.V.; Juncos, J.P.; Croatt, A.J.; Manriquez, M.C.; Pittock, S.T.; Katusic, Z.S. An analysis of the DOCA-salt model of hypertension in HO-1-/- mice and the Gunn rat. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H333–H342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pflueger, A.; Croatt, A.J.; Peterson, T.E.; Smith, L.A.; d’Uscio, L.V.; Katusic, Z.S.; Nath, K.A. The hyperbilirubinemic Gunn rat is resistant to the pressor effects of angiotensin II. Am. J. Physiol. Ren. Physiol. 2005, 288, F552–F558. [Google Scholar] [CrossRef]
- Bakrania, B.; Du Toit, E.F.; Ashton, K.J.; Kiessling, C.J.; Wagner, K.H.; Headrick, J.P.; Bulmer, A.C. Hyperbilirubinemia modulates myocardial function, aortic ejection, and ischemic stress resistance in the Gunn rat. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H1142–H1149. [Google Scholar] [CrossRef] [Green Version]
- Bakrania, B.; Du Toit, E.F.; Ashton, K.J.; Wagner, K.H.; Headrick, J.P.; Bulmer, A.C. Chronically elevated bilirubin protects from cardiac reperfusion injury in the male Gunn rat. Acta Physiol. 2017, 220, 461–470. [Google Scholar] [CrossRef]
- Bianco, A.; Pinci, S.; Tiribelli, C.; Bellarosa, C. Life-Long Hyperbilirubinemia Exposure and Bilirubin Priming Prevent In Vitro Metabolic Damage. Front Pharm. 2021, 12, 646953. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.M.; Adeosun, S.O.; Ngwudike, S.I.; Anderson, C.D.; Hall, J.E.; Hinds, T.D., Jr.; Stec, D.E. CRISPR Cas9-mediated deletion of biliverdin reductase A (BVRA) in mouse liver cells induces oxidative stress and lipid accumulation. Arch. Biochem. Biophys. 2019, 672, 108072. [Google Scholar] [CrossRef] [PubMed]
- Hinds, T.D., Jr.; Burns, K.A.; Hosick, P.A.; McBeth, L.; Nestor-Kalinoski, A.; Drummond, H.A.; AlAmodi, A.A.; Hankins, M.W.; Vanden Heuvel, J.P.; Stec, D.E. Biliverdin reductase A attenuates hepatic steatosis by inhibition of glycogen synthase kinase (GSK) 3beta phosphorylation of serine 73 of peroxisome proliferator-activated receptor (PPAR) alpha. J. Biol. Chem. 2016, 291, 25179–25191. [Google Scholar] [CrossRef] [Green Version]
- Stec, D.E.; Gordon, D.M.; Nestor-Kalinoski, A.L.; Donald, M.C.; Mitchell, Z.L.; Creeden, J.F.; Hinds, T.D., Jr. Biliverdin Reductase A (BVRA) Knockout in Adipocytes Induces Hypertrophy and Reduces Mitochondria in White Fat of Obese Mice. Biomolecules 2020, 10, 387. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Maghzal, G.J.; Ayer, A.; Suarna, C.; Dunn, L.L.; Stocker, R. Absence of the biliverdin reductase-a gene is associated with increased endogenous oxidative stress. Free Radic. Biol. Med. 2018, 115, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Adeosun, S.O.; Moore, K.H.; Lang, D.M.; Nwaneri, A.C.; Hinds, T.D., Jr.; Stec, D.E. A Novel Fluorescence-Based Assay for the Measurement of Biliverdin Reductase Activity. React. Oxyg. Species 2018, 5, 35–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.; Gordon, D.; Stec, D.E.; Hinds, T.D. Bilirubin: A Ligand of the PPARα Nuclear Receptor. In Nuclear Receptors: The Art and Science of Modulator Design and Discovery; Badr, M.Z., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 463–482. [Google Scholar] [CrossRef]
- Sundararaghavan, V.L.; Binepal, S.; Stec, D.E.; Sindhwani, P.; Hinds, T.D., Jr. Bilirubin, a new therapeutic for kidney transplant? Transplant. Rev. 2018, 32, 234–240. [Google Scholar] [CrossRef]
- Hinds, T.D., Jr.; Hosick, P.A.; Hankins, M.W.; Nestor-Kalinoski, A.; Stec, D.E. Mice with hyperbilirubinemia due to Gilbert’s Syndrome polymorphism are resistant to hepatic steatosis by decreased serine 73 phosphorylation of PPARalpha. Am. J. Physiol. Endocrinol. Metab. 2017, 312, E244–E252. [Google Scholar] [CrossRef] [Green Version]
- Adeosun, S.O.; Gordon, D.M.; Weeks, M.F.; Moore, K.H.; Hall, J.E.; Hinds, T.D., Jr.; Stec, D.E. Loss of biliverdin reductase-A promotes lipid accumulation and lipotoxicity in mouse proximal tubule cells. Am. J. Physiol. Renal Physiol. 2018, 315, F323–F331. [Google Scholar] [CrossRef]
- Vitek, L.; Tiribelli, C. Bilirubin: The yellow hormone? J. Hepatol. 2021, 75, 1485–1490. [Google Scholar] [CrossRef]
- Ai, W.; Bae, S.; Ke, Q.; Su, S.; Li, R.; Chen, Y.; Yoo, D.; Lee, E.; Jon, S.; Kang, P.M. Bilirubin Nanoparticles Protect Against Cardiac Ischemia/Reperfusion Injury in Mice. J. Am. Heart Assoc. 2021, 10, e021212. [Google Scholar] [CrossRef]
- Hjelkrem, M.; Morales, A.; Williams, C.D.; Harrison, S.A. Unconjugated hyperbilirubinemia is inversely associated with non-alcoholic steatohepatitis (NASH). Aliment. Pharmacol. Ther. 2012, 35, 1416–1423. [Google Scholar] [CrossRef]
- Petrtyl, J.; Dvorak, K.; Stritesky, J.; Lenicek, M.; Jiraskova, A.; Smid, V.; Haluzik, M.; Bruha, R.; Vitek, L. Association of Serum Bilirubin and Functional Variants of Heme Oxygenase 1 and Bilirubin UDP-Glucuronosyl Transferase Genes in Czech Adult Patients with Non-Alcoholic Fatty Liver Disease. Antioxidants 2021, 10, 2000. [Google Scholar] [CrossRef]
- Salomone, F.; Li Volti, G.; Rosso, C.; Grosso, G.; Bugianesi, E. Unconjugated bilirubin, a potent endogenous antioxidant, is decreased in patients with non-alcoholic steatohepatitis and advanced fibrosis. J. Gastroenterol. Hepatol. 2013, 28, 1202–1208. [Google Scholar] [CrossRef]
- Kwak, M.S.; Kim, D.; Chung, G.E.; Kang, S.J.; Park, M.J.; Kim, Y.J.; Yoon, J.H.; Lee, H.S. Serum bilirubin levels are inversely associated with nonalcoholic fatty liver disease. Clin. Mol. Hepatol. 2012, 18, 383–390. [Google Scholar] [CrossRef]
- Badmus, O.O.; Hillhouse, S.A.; Anderson, C.D.; Hinds, T.D.; Stec, D.E. Molecular mechanisms of metabolic associated fatty liver disease (MAFLD): Functional analysis of lipid metabolism pathways. Clin. Sci. 2022, 136, 1347–1366. [Google Scholar] [CrossRef] [PubMed]
- Tarantino, G.; Colao, A.; Capone, D.; Conca, P.; Tarantino, M.; Grimaldi, E.; Chianese, D.; Finelli, C.; Contaldo, F.; Scopacasa, F.; et al. Circulating levels of cytochrome C, gamma-glutamyl transferase, triglycerides and unconjugated bilirubin in overweight/obese patients with non-alcoholic fatty liver disease. J. Biol. Regul. Homeost. Agents 2011, 25, 47–56. [Google Scholar]
- Kunutsor, S.K.; Frysz, M.; Verweij, N.; Kieneker, L.M.; Bakker, S.J.L.; Dullaart, R.P.F. Circulating total bilirubin and risk of non-alcoholic fatty liver disease in the PREVEND study: Observational findings and a Mendelian randomization study. Eur. J. Epidemiol. 2020, 35, 123–137. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.H.; Yun, K.E.; Choi, H.J. Relationships between serum total bilirubin levels and metabolic syndrome in Korean adults. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, M.; Xu, M.; Bi, Y.; Li, X.; Chen, Y.; Ning, G.; Wang, W. Low serum total bilirubin concentrations are associated with increased prevalence of metabolic syndrome in Chinese. J. Diabetes 2011, 3, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K.M.; Kam, J.H.; Kim, M.Y.; Kim, M.Y.; Chung, C.H.; Kim, J.K.; Linton, J.A.; Eom, A.; Koh, S.B.; Kang, H.T. Inverse association between total bilirubin and metabolic syndrome in rural korean women. J. Women’s Health 2011, 20, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Guzek, M.; Jakubowski, Z.; Bandosz, P.; Wyrzykowski, B.; Smoczynski, M.; Jabloiska, A.; Zdrojewski, T. Inverse association of serum bilirubin with metabolic syndrome and insulin resistance in Polish population. Prz. Epidemiol. 2012, 66, 495–501. [Google Scholar]
- Ghem, C.; Sarmento-Leite, R.E.; de Quadros, A.S.; Rossetto, S.; Gottschall, C.A. Serum bilirubin concentration in patients with an established coronary artery disease. Int. Heart J. 2010, 51, 86–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwertner, H.A.; Jackson, W.G.; Tolan, G. Association of low serum concentration of bilirubin with increased risk of coronary artery disease. Clin. Chem. 1994, 40, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.F.; Chen, Y.Z.; Su, J.L.; Wang, F.Y.; Wang, L.X. Relationship between serum bilirubin and carotid atherosclerosis in hypertensive patients. Intern. Med. 2009, 48, 1595–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.-M.; Gao, Y.; Zheng, Y.-Y.; Chen, Y.; Liu, F.; Ma, Y.-T.; Xie, X. Association of Fasting Serum Bilirubin Levels with Clinical Outcomes After Percutaneous Coronary Intervention: A Prospective Study. Cardiovasc. Toxicol. 2017, 17, 471–477. [Google Scholar] [CrossRef]
- Hinds, T.D., Jr.; Adeosun, S.O.; Alamodi, A.A.; Stec, D.E. Does bilirubin prevent hepatic steatosis through activation of the PPARalpha nuclear receptor? Med. Hypotheses 2016, 95, 54–57. [Google Scholar] [CrossRef] [Green Version]
- Seyed Khoei, N.; Grindel, A.; Wallner, M.; Molzer, C.; Doberer, D.; Marculescu, R.; Bulmer, A.; Wagner, K.H. Mild hyperbilirubinaemia as an endogenous mitigator of overweight and obesity: Implications for improved metabolic health. Atherosclerosis 2018, 269, 306–311. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement | Lean | Obese | p-Value |
|---|---|---|---|
| Age | 31.55 ± 13.09 | 40.91 ± 17.11 | 0.632 |
| BMI | 22.47 ± 2.73 | 35.11 ± 3.53 | 0.057 |
| Body Weight (kg) | 64.60 ± 11.03 | 99.37 ± 12.21 | 0.256 |
| Total Fat (kg) | 14.73 ± 4.80 | 45.28 ± 7.33 | 0.022 * |
| Body Fat % | 21.69 ± 5.66 | 44.42 ± 5.16 | 0.003 * |
| Total Bilirubin (mg/dL) | 0.77 ± 0.59 | 0.52 ± 0.30 | 0.529 |
| Urobilin (μM) | 9.57 ± 4.50 | 23.38 ± 19.30 | 0.026 * |
| Blood Glucose | 87.57 ± 10.66 | 98.06 ± 10.64 | 0.246 |
| Blood Insulin | 7.21 ± 4.66 | 18.06 ± 15.51 | 0.398 |
| HOMA IR | 1.63 ± 1.25 | 4.59 ± 4.37 | 0.398 |
| Measurement | Lean Female | Obese Female | p-Value | Lean Male | Obese Male | p-Value |
|---|---|---|---|---|---|---|
| Age | 29.76 ± 12.82 | 39.31 ± 12.61 | 0.155 | 33.14 ± 13.91 | 43.45 ± 24.23 | 0.4180 |
| BMI | 21.33 ± 2.54 | 34.46 ± 1.72 | 3.40 × 10−8 * | 23.47 ± 2.62 | 36.14 ± 5.48 | 0.0045 * |
| Body Weight (g) | 56.11 ± 7.21 | 93.07 ± 7.85 | 1.25 × 10−7 * | 72.14 ± 7.85 | 109.42 ± 11.59 | 0.0006 * |
| Total Fat (g) | 14.05 ± 4.97 | 44.68 ± 6.29 | 5.88 × 10−8 * | 15.32 ± 4.86 | 46.24 ± 9.50 | 0.0008 * |
| Body Fat % | 23.51 ± 6.50 | 46.61 ± 2.88 | 4.38 × 10−6 * | 20.08 ± 4.57 | 40.92 ± 6.35 | 0.0005 * |
| Total Bilirubin (mg/dL) | 0.74 ± 0.82 | 0.54 ± 0.33 | 0.527 | 0.79 ± 0.30 | 0.50 ± 0.28 | 0.0954 |
| Urobilin (μM) | 11.07 ± 3.79 | 31.21 ± 20.58 | 0.028 * | 8.23 ± 4.85 | 10.86 ± 7.60 | 0.5108 |
| Blood Glucose | 87.38 ± 13.53 | 98.78 ± 11.21 | 0.089 | 87.73 ± 8.19 | 96.93 ± 10.81 | 0.1440 |
| Blood Insulin | 5.95 ± 5.11 | 17.06 ± 9.26 | 0.013 * | 8.33 ± 4.20 | 19.64 ± 23.8 | 0.3493 |
| HOMA IR | 1.42 ± 1.52 | 4.26 ± 2.40 | 0.016 * | 1.81 ± 0.94 | 5.13 ± 6.83 | 0.3394 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kipp, Z.A.; Xu, M.; Bates, E.A.; Lee, W.-H.; Kern, P.A.; Hinds, T.D., Jr. Bilirubin Levels Are Negatively Correlated with Adiposity in Obese Men and Women, and Its Catabolized Product, Urobilin, Is Positively Associated with Insulin Resistance. Antioxidants 2023, 12, 170. https://doi.org/10.3390/antiox12010170
Kipp ZA, Xu M, Bates EA, Lee W-H, Kern PA, Hinds TD Jr. Bilirubin Levels Are Negatively Correlated with Adiposity in Obese Men and Women, and Its Catabolized Product, Urobilin, Is Positively Associated with Insulin Resistance. Antioxidants. 2023; 12(1):170. https://doi.org/10.3390/antiox12010170
Chicago/Turabian StyleKipp, Zachary A., Mei Xu, Evelyn A. Bates, Wang-Hsin Lee, Philip A. Kern, and Terry D. Hinds, Jr. 2023. "Bilirubin Levels Are Negatively Correlated with Adiposity in Obese Men and Women, and Its Catabolized Product, Urobilin, Is Positively Associated with Insulin Resistance" Antioxidants 12, no. 1: 170. https://doi.org/10.3390/antiox12010170
APA StyleKipp, Z. A., Xu, M., Bates, E. A., Lee, W. -H., Kern, P. A., & Hinds, T. D., Jr. (2023). Bilirubin Levels Are Negatively Correlated with Adiposity in Obese Men and Women, and Its Catabolized Product, Urobilin, Is Positively Associated with Insulin Resistance. Antioxidants, 12(1), 170. https://doi.org/10.3390/antiox12010170