
Vitamin E-Inhibited Phoxim-Induced Renal Oxidative Stress and Mitochondrial Apoptosis In Vivo and In Vitro of Piglets

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Experimental Design
2.2.1. Animal Experiments
2.2.2. Cell Experiments
2.2.3. Sample Collection
2.2.4. Serum Biochemical Analysis
2.2.5. Oxidation–Antioxidant Parameters
2.2.6. Histopathological Examinations
2.2.7. Ultrastructure of the Mitochondria
2.2.8. Mitochondrial Function
2.2.9. Flow Cytometer Analysis of Apoptosis
2.2.10. Mitochondrial Membrane Potential
2.2.11. Quantitative Real-Time PCR (qRT-PCR)
2.3. Statistical Analysis
3. Results
3.1. Serum Biochemical Analysis
3.2. Oxidant-Antioxidant Parameters
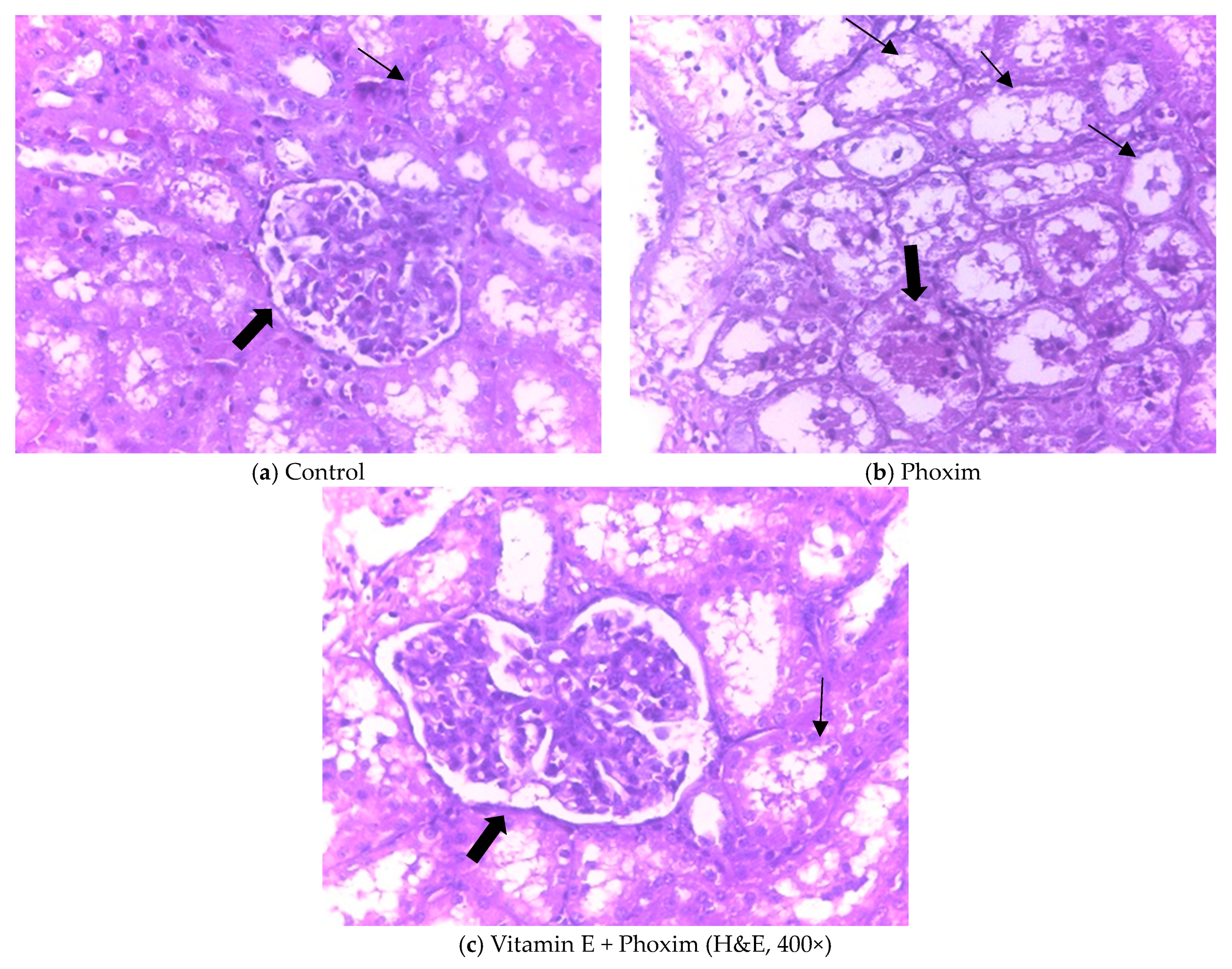
3.3. Histopathological Findings
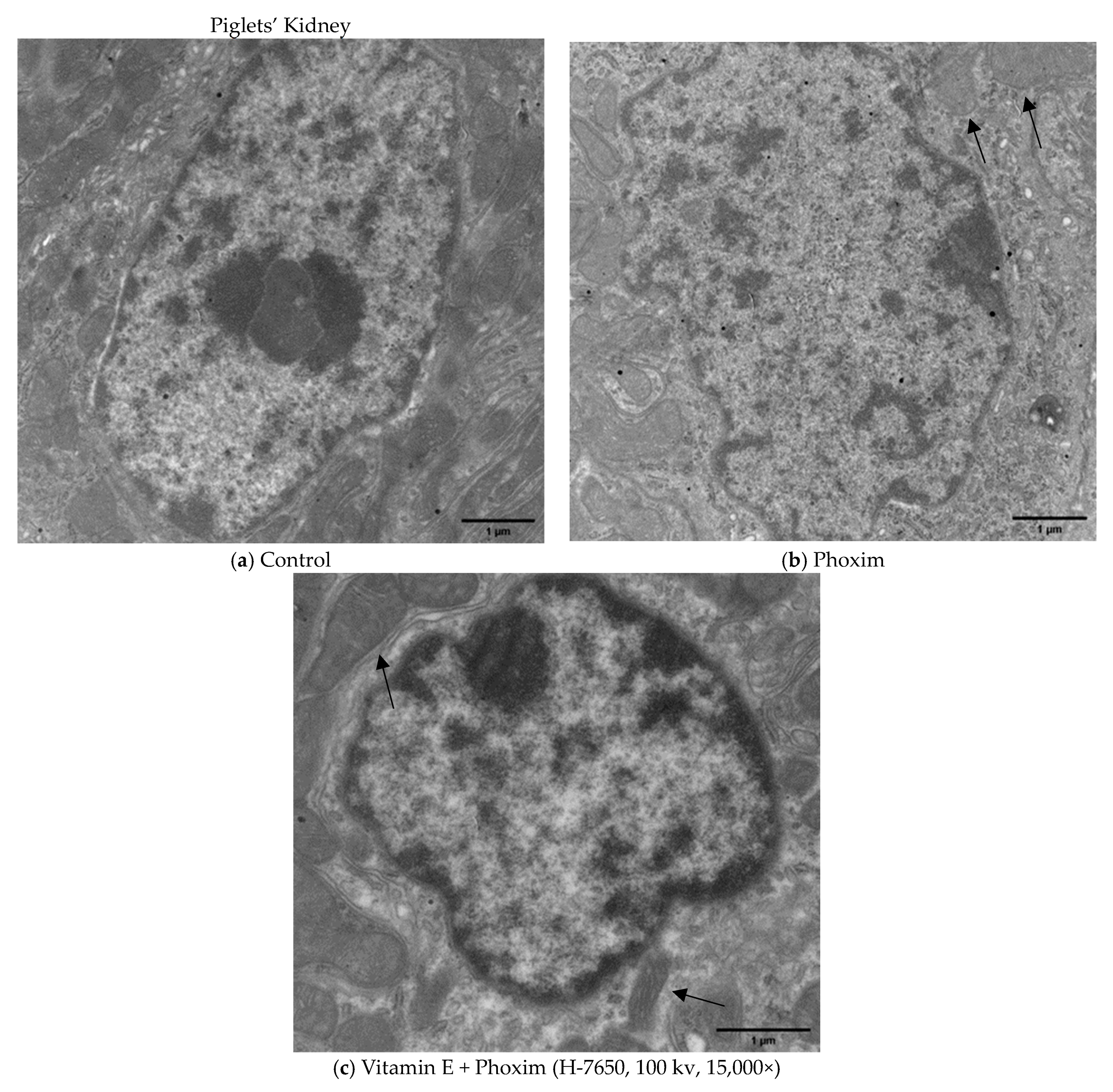
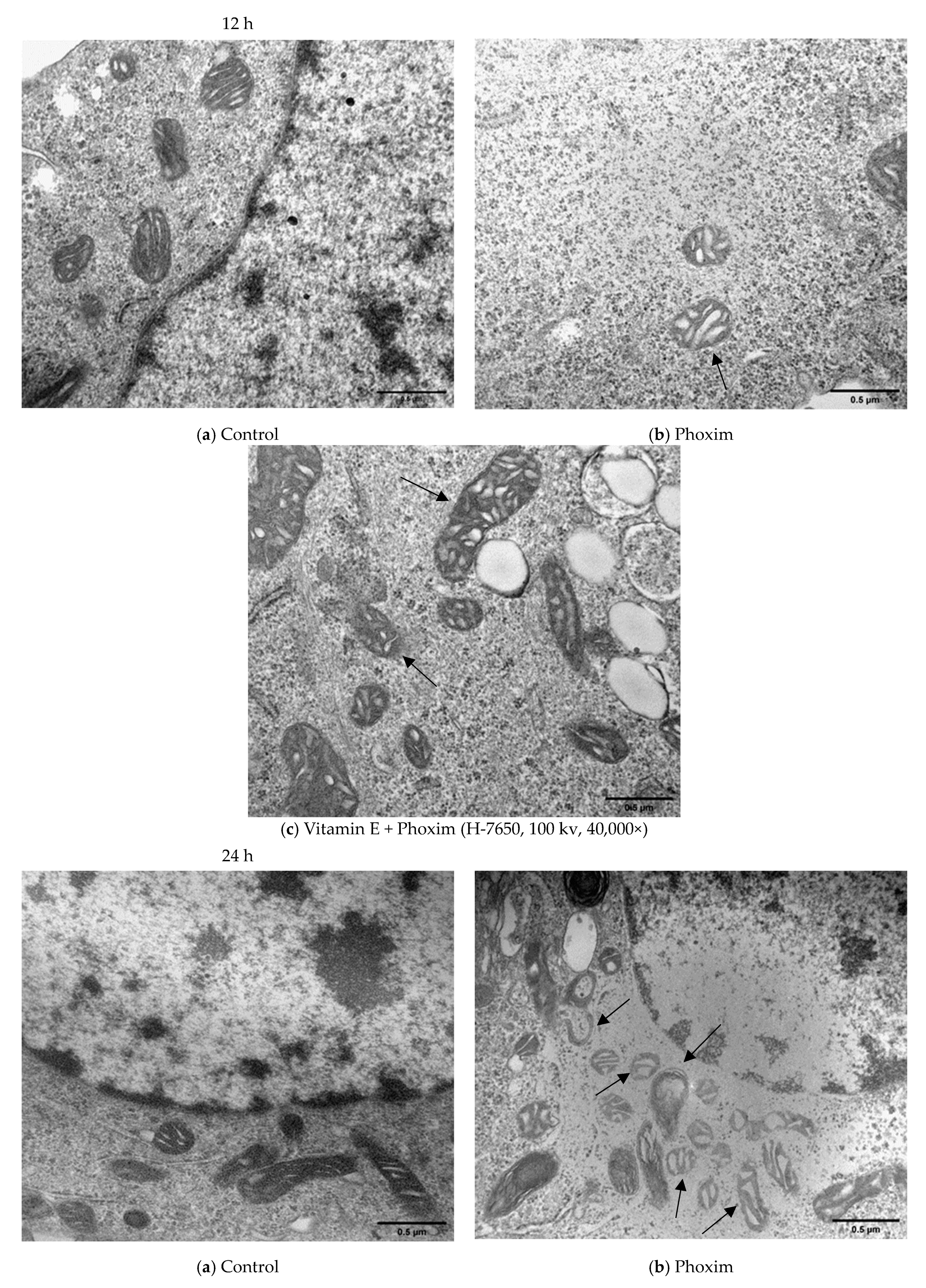
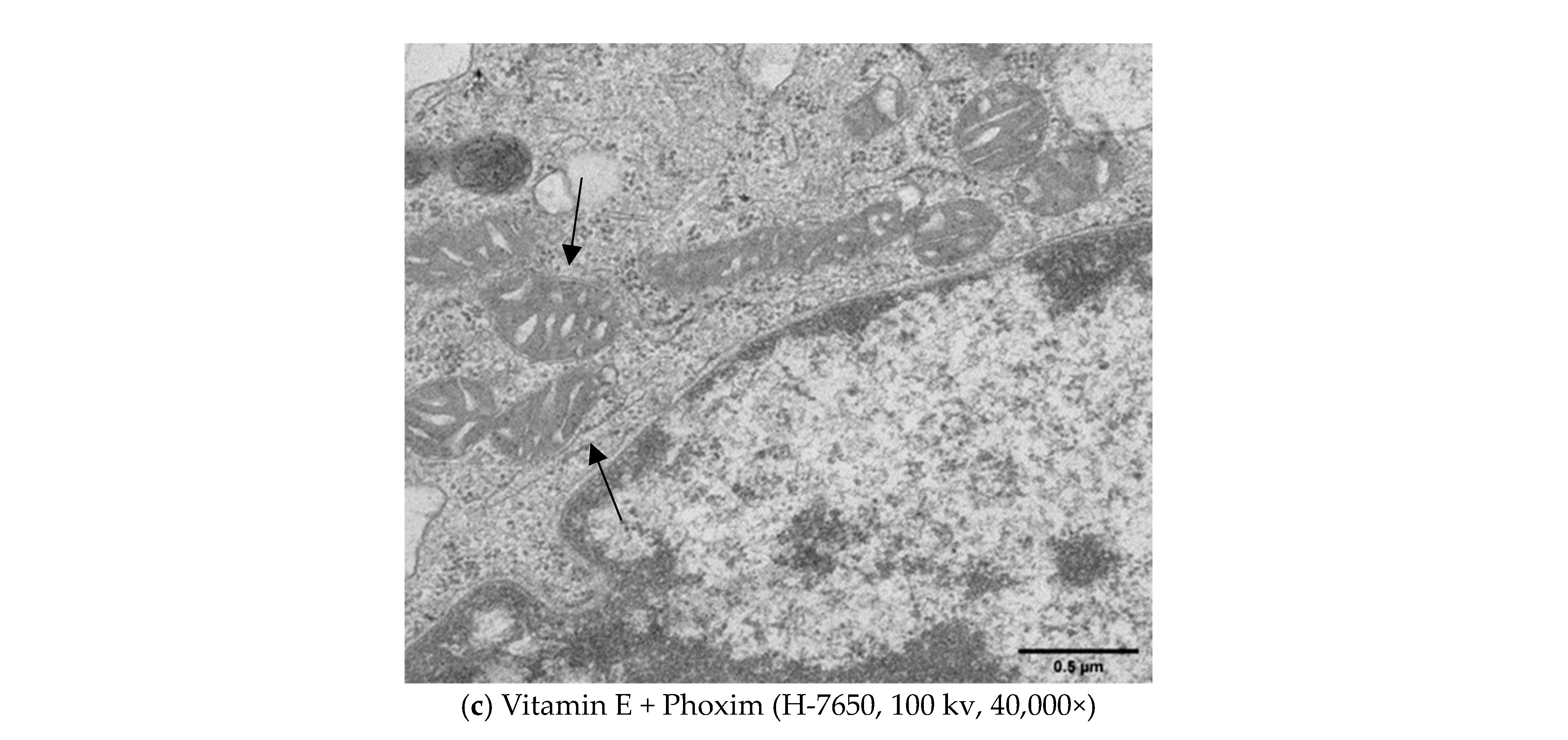
3.4. Ultrastructure of the Mitochondria
3.5. Mitochondrial Function
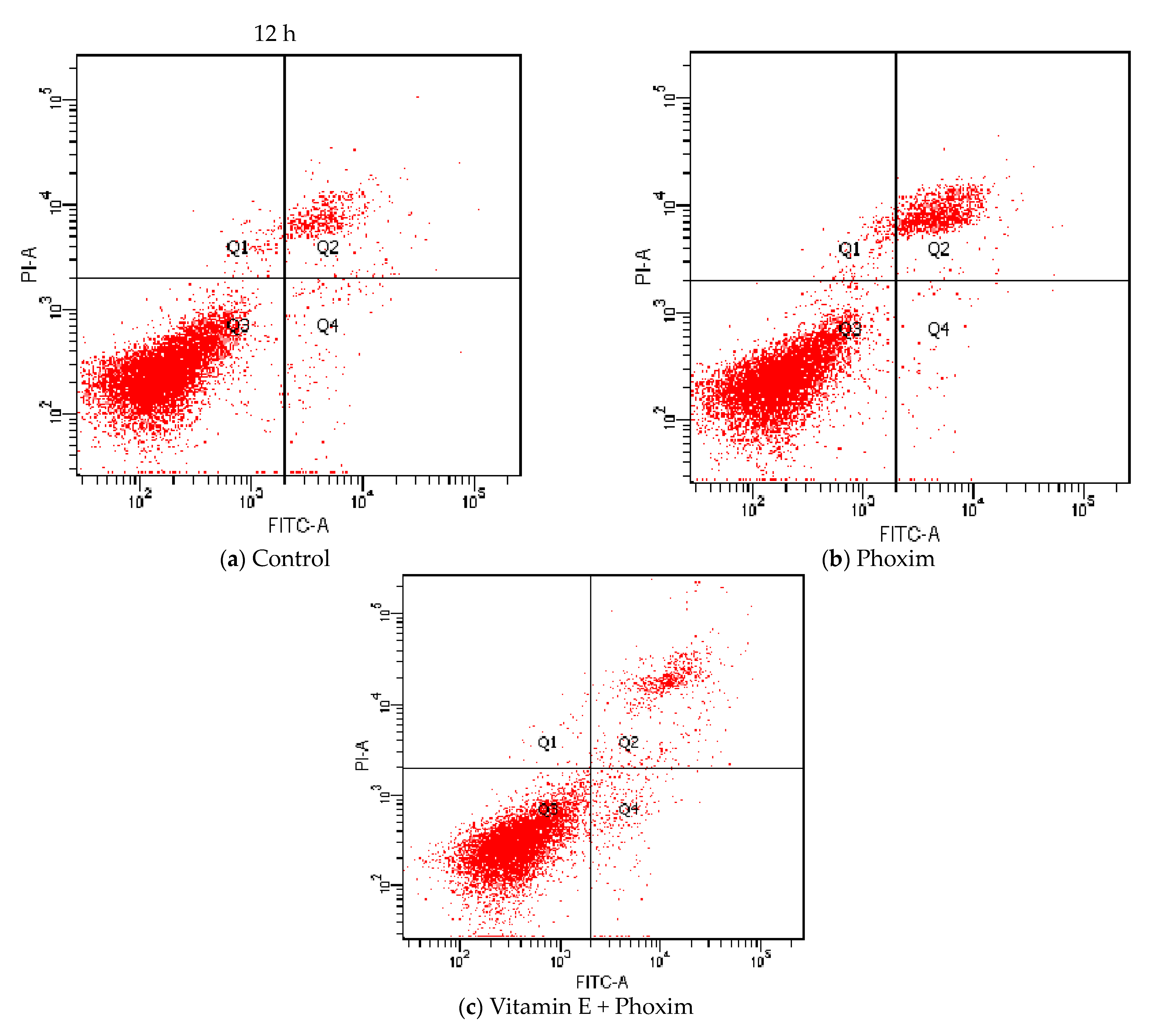
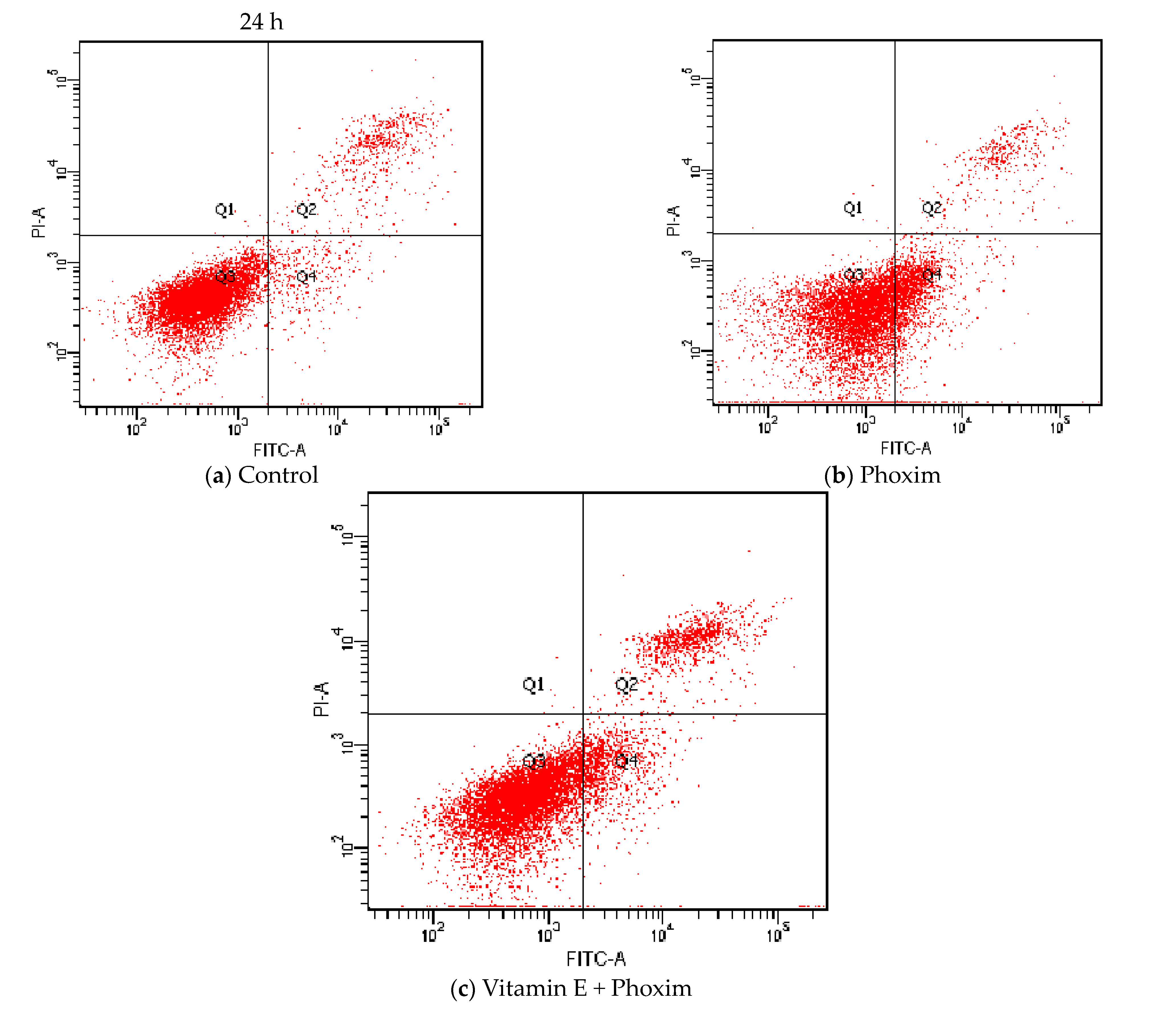
3.6. Cell Apoptosis Assay
3.7. Mitochondrial Membrane Potential
3.8. Quantitative Real-Time PCR (qRT-PCR)
4. Discussion
4.1. Serum Biochemical Analysis
4.2. Oxidation-Antioxidation
4.3. Histopathological Findings
4.4. Ultrastructure of the Mitochondria
4.5. Mitochondrial Enzyme Activity
4.6. Apoptosis
4.6.1. Mitochondrial Transmembrane Potential
4.6.2. Apoptosis Rate
4.6.3. Apoptotic Factor mRNA
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, J.; Zhang, Q.; Zhang, Z.; Zhou, Z.; Lu, T.; Sun, L.; Qian, H. Evaluation of phoxim toxicity on aquatic and zebrafish intestinal microbiota by metagenomics and 16S rRNA gene sequencing analysis. Environ. Sci. Pollut. Res. Int. 2022, 29, 63017–63027. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, J.; Song, W.; Shan, A. Vitamin E alleviates phoxim-induced toxic effects on intestinal oxidative stress, barrier function, and morphological changes in rats. Environ. Sci. Pollut. Res. 2018, 25, 26682–26692. [Google Scholar] [CrossRef] [PubMed]
- Forget, J.; Bocquene, G. Partial purification and enzymatic characterization of acetylcholinesterase from the intertidal marine copepod Tigriopus brevicornis. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1999, 123, 345–350. [Google Scholar] [CrossRef]
- Pope, C.; Karanth, S.; Liu, J. Pharmacology and toxicology of cholinesterase inhibitors: Uses and misuses of a common mechanism of action. Environ. Toxicol. Pharmacol. 2005, 19, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Song, W.; Sun, Y.; Shan, A. Effects of phoxim-induced hepatotoxicity on SD rats and the protection of vitamin E. Environ. Sci. Pollut. Res. 2017, 24, 24916–24927. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zheng, S.; Pu, Y.; Shu, L.; Sun, L.; Liu, W.; Fu, Z. Cypermethrin has the potential to induce hepatic oxidative stress, DNA damage and apoptosis in adult zebrafish (Danio rerio). Chemosphere 2011, 82, 398–404. [Google Scholar] [CrossRef]
- Traber, M.G.; Atkinson, J. Vitamin E, antioxidant and nothing more. Free Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef]
- Peh, H.Y.; Tan, W.D.; Liao, W.; Wong, W.F. Vitamin E therapy beyond cancer: Tocopherol versus tocotrienol. Pharmacol. Ther. 2016, 162, 152–169. [Google Scholar] [CrossRef]
- Weng, J.; Cai, B.; Chen, J.; Chen, B.; Wu, H.; Wen, C.; Wang, Z. Metabolic changes in methomyl poisoned rats treated by vitamin E. Hum. Exp. Toxicol. 2018, 37, 390–398. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Abdeen, A. Protective effects of rosuvastatin and vitamin E against fipronil-mediated oxidative damage and apoptosis in rat liver and kidney. Food Chem. Toxicol. 2018, 114, 69–77. [Google Scholar] [CrossRef]
- Zhang, J. Role of Endoplasmic Reticulum Stress-Induced Disorder of Mitochondrial Morphology and Function in Lead Neurotoxicity and the Underlying Molecular Mechanism. Doctoral Dissertation, Fourth Military Medical University, Xi’an, China, 2014. [Google Scholar]
- Fu, Y.; Yang, X. Mechanisms of IMS proteins transport. J. Med. Mol. Biol. 2006, 3, 194–197. [Google Scholar]
- Nohl, H.; Gille, L.; Staniek, K. The mystery of reactive oxygen species derived from cell respiration. Acta Biochim. Pol. 2004, 51, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Toklu, H.Z.; Şehirli, Ö.; Erşahin, M.; Süleymanoğlu, S.; Yiğiner, Ö.; Emekli-Alturfan, E.; Şener, G. Resveratrol improves cardiovascular function and reduces oxidative organ damage in the renal, cardiovascular and cerebral tissues of two-kidney, one-clip hypertensive rats. J. Pharm. Pharmacol. 2010, 62, 1784–1793. [Google Scholar] [CrossRef] [PubMed]
- Van Houten, B.; Woshner, V.; Santos, J.H. Role of mitochondrial DNA in toxic responses to oxidative stress. DNA Repair 2006, 5, 145–152. [Google Scholar] [CrossRef]
- Di Paola, D.; D’Amico, R.; Genovese, T.; Siracusa, R.; Cordaro, M.; Crupi, R.; Peritore, A.F.; Gugliandolo, E.; Interdonato, L.; Impellizzeri, D.; et al. Chronic Exposure to Vinclozolin Induced Fibrosis, Mitochondrial Dysfunction, Oxidative Stress, and Apoptosis in Mice Kidney. Int. J. Mol. Sci. 2022, 23, 11296. [Google Scholar] [CrossRef]
- Forbes, J.M. Mitochondria-Power Players in Kidney Function? Trends Endocrinol. Metab. 2016, 27, 441–442. [Google Scholar] [CrossRef]
- Li, Z.; Liu, S.; Zhao, Y.; Wang, J.; Ma, X. Compound organic acid could improve the growth performance, immunity and antioxidant properties, and intestinal health by altering the microbiota profile of weaned piglets. J. Anim. Sci. 2023, 101, skad196. [Google Scholar] [CrossRef]
- Ma, K.; Wu, H.Y.; Wang, S.Y.; Li, B.X. The Keap1/Nrf2-ARE signaling pathway is involved in atrazine induced dopaminergic neurons degeneration via microglia activation. Ecotoxicol. Environ. Saf. 2021, 226, 112862. [Google Scholar] [CrossRef]
- Gao, C.; He, H.; Qiu, W.; Zheng, Y.; Chen, Y.; Hu, S.; Zhao, X. Oxidative Stress, Endocrine Disturbance, and Immune Interference in Humans Showed Relationships to Serum Bisphenol Concentrations in a Dense Industrial Area. Environ. Sci. Technol. 2021, 55, 1953–1963. [Google Scholar] [CrossRef]
- El-Sawi Mahmoud, N.M.; El-Wassimy, M.M.; Youssef, O.A. Effect of verrucarin J toxin on liver and kidney tissue of male mice. Tex. J. Sci. 2001, 53, 375–384. [Google Scholar]
- Pandey, P.C.; Mishra, A.P. Novel potentiometric sensing of creatinine. Sens. Actuators B Chem. 2004, 99, 230–235. [Google Scholar] [CrossRef]
- Leomanni, A.; Schettino, T.; Calisi, A.; Gorbi, S.; Mezzelani, M.; Regoli, F.; Lionetto, M.G. Antioxidant and oxidative stress related responses in the Mediterranean land snail Cantareus apertus exposed to the carbamate pesticide Carbaryl. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2015, 168, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Cemek, M.; Büyükben, A.; Büyükokuroğlu, M.E.; Aymelek, F.; Tür, L. Protective roles of vitamin E (α-tocopherol), selenium and vitamin E plus selenium in organophosphate toxicity in vivo: A comparative study. Pestic. Biochem. Physiol. 2010, 96, 113–118. [Google Scholar] [CrossRef]
- Han, Y.; Song, S.; Wu, H.; Zhang, J.; Ma, E. Antioxidant enzymes and their role in phoxim and carbaryl stress in Caenorhabditis elegans. Pestic. Biochem. Physiol. 2017, 138, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Rehman, Z.U.; Che, L.; Ren, S.; Liao, Y.; Qiu, X.; Yu, S.; Ding, Z. Supplementation of vitamin E protects chickens from Newcastle disease virus-mediated exacerbation of intestinal oxidative stress and tissue damage. Cell. Physiol. Biochem. 2018, 47, 1655–1666. [Google Scholar] [CrossRef]
- Karami, M.; Mostafazadeh, M.; Sadeghi, H.; Sadeghi, H.; Mehraban, F.; Kokhdan, E.P.; Abtahi, S.R. Nephroprotective effect of Nasturtium officinale (watercress) ethanol extract and Vitamin E on vancomycin-induced nephrotoxicity in rats. Jundishapur J. Nat. Pharm. Prod. 2018, 13, 67178. [Google Scholar] [CrossRef]
- Soleimani, E.; Moghadam, R.H.; Ranjbar, A. Occupational exposure to chemicals and oxidative toxic stress. Toxicol. Environ. Health Sci. 2015, 7, 1–24. [Google Scholar] [CrossRef]
- Wani, W.Y.; Gudup, S.; Sunkaria, A.; Bal, A.; Singh, P.P.; Kandimalla, R.J.; Gill, K.D. Protective efficacy of mitochondrial targeted antioxidant MitoQ against dichlorvos induced oxidative stress and cell death in rat brain. Neuropharmacology 2011, 61, 1193–1201. [Google Scholar] [CrossRef]
- Binukumar, B.K.; Bal, A.; Sunkaria, A.; Gill, K.D. Mitochondrial energy metabolism impairment and liver dysfunction following chronic exposure to dichlorvos. Toxicology 2010, 270, 77–84. [Google Scholar] [CrossRef]
- Pavithra, D.; Praveen, D.; Chowdary, P.R.; Aanandhi, M.V. A review on role of Vitamin E supplementation in type 2 diabetes mellitus. Drug Invent. Today 2018, 10, 236–240. [Google Scholar]
- Reshi, M.L.; Su, Y.C.; Hong, J.R. RNA viruses: ROS-mediated cell death. Int. J. Cell Biol. 2014, 2014, 467452. [Google Scholar] [CrossRef] [PubMed]
- Min, Y.N.; Niu, Z.Y.; Sun, T.T.; Wang, Z.P.; Jiao, P.X.; Zi, B.B.; Liu, F.Z. Vitamin E and vitamin C supplementation improves antioxidant status and immune function in oxidative-stressed breeder roosters by up-regulating expression of GSH-Px gene. Poult. Sci. 2018, 97, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Oyagbemi, A.A.; Omobowale, T.O.; Ola-Davies, O.E.; Adejumobi, O.A.; Asenuga, E.R.; Adeniji, F.K.; Yakubu, M.A. Protective Effect of Azadirachta indica and Vitamin E Against Arsenic Acid-Induced Genotoxicity and Apoptosis in Rats. J. Diet. Suppl. 2018, 15, 251–268. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F. Antioxidant systems in poultry biology: Superoxide dismutase. J. Anim. Res. Nutr. 2016, 1, 8. [Google Scholar] [CrossRef]
- Spodniewska, A.; Barski, D.; Giżejewska, A. Effect of enrofloxacin and chlorpyrifos on the levels of vitamins A and E in Wistar rats. Environ. Toxicol. Pharmacol. 2015, 40, 587–591. [Google Scholar] [CrossRef]
- DelCurto, H.; Wu, G.; Satterfield, M.C. Nutrition and reproduction: Links to epigenetics and metabolic syndrome in offspring. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 385–391. [Google Scholar] [CrossRef]
- Ghlissi, Z.; Hakim, A.; Mnif, H.; Kallel, R.; Zeghal, K.; Boudawara, T.; Sahnoun, Z. Effect of vitamin E on reversibility of renal function following discontinuation of colistin in rats: Histological and biochemical investigations. Saudi J. Kidney Dis. Transplant. 2018, 29, 10–18. [Google Scholar] [CrossRef]
- Kalender, S.; Ogutcu, A.; Uzunhisarcikli, M.; Açikgoz, F.; Durak, D.; Ulusoy, Y.; Kalender, Y. Diazinon-induced hepatotoxicity and protective effect of vitamin E on some biochemical indices and ultrastructural changes. Toxicology 2005, 211, 197–206. [Google Scholar] [CrossRef]
- Ourique, G.M.; Saccol, E.M.; Pês, T.S.; Glanzner, W.G.; Schiefelbein, S.H.; Woehl, V.M.; Barreto, K.P. Protective effect of vitamin E on sperm motility and oxidative stress in valproic acid treated rats. Food Chem. Toxicol. 2016, 95, 159–167. [Google Scholar] [CrossRef]
- Emam, H.; Ahmed, E.; Abdel-Daim, M. Antioxidant capacity of omega-3-fatty acids and vitamin E against imidacloprid-induced hepatotoxicity in Japanese quails. Environ. Sci. Pollut. Res. 2018, 25, 11694–11702. [Google Scholar] [CrossRef]
- Dalia, A.M.; Loh, T.C.; Sazili, A.Q.; Jahromi, M.F.; Samsudin, A.A. Effects of vitamin E, inorganic selenium, bacterial organic selenium, and their combinations on immunity response in broiler chickens. BMC Vet. Res. 2018, 14, 249. [Google Scholar] [CrossRef] [PubMed]
- Richardson, N.; Gordon, A.K.; Muller, W.J.; Pletschke, B.I.; Whitfield, A.K. The use of liver histopathology, lipid peroxidation and acetylcholinesterase assays as biomarkers of contaminant-induced stress in the cape stumpnose, Rhabdosargus holubi (teleostei: Sparidae), from selected south african estuaries. Water Sa 2010, 36, 407–416. [Google Scholar] [CrossRef]
- Deng, Y.; Zhang, Y.; Lu, Y.; Zhao, Y.; Ren, H. Hepatotoxicity and nephrotoxicity induced by the chlorpyrifos and chlorpyrifos-methyl metabolite, 3,5,6-trichloro-2-pyridinol, in orally exposed mice. Sci. Total Environ. 2016, 544, 507–514. [Google Scholar] [CrossRef]
- Yadav, B.; Niyogi, D.; Tripathi, K.K.; Singh, G.K.; Yadav, A.; Kumar, M. Patho-morphological effects in broiler birds induced with sub-acute chlorpyrifos toxicity and its amelioration with vitamin E and selenium. J. Pharmacogn. Phytochem. 2018, 7, 1877–1882. [Google Scholar]
- Kalender, S.; Uzun, F.G.; Durak, D.; Demir, F.; Kalender, Y. Malathion-induced hepatotoxicity in rats: The effects of vitamins C and E. Food Chem. Toxicol. 2010, 48, 633–638. [Google Scholar] [CrossRef]
- Flemming, N.B.; Gallo, L.A.; Forbes, J.M. Mitochondrial Dysfunction and Signaling in Diabetic Kidney Disease: Oxidative Stress and Beyond. Semin. Nephrol. 2018, 38, 101–110. [Google Scholar] [CrossRef]
- Mao, R.W.; He, S.P.; Lan, J.G.; Zhu, W.Z. Honokiol ameliorates cisplatin-induced acute kidney injury via inhibition of mitochondrial fission. Br. J. Pharmacol. 2022, 179, 3886–3904. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Dong, Z.; Harris, R.; Murray, P.; Parikh, S.M.; Rosner, M.H.; Kellum, J.A.; Ronco, C.; for the Acute Dialysis Quality Initiative XIII Working Group. Cellular and Molecular Mechanisms of AKI. J. Am. Soc. Nephrol. 2016, 27, 1288–1299. [Google Scholar] [CrossRef]
- Noh, M.R.; Kim, K.Y.; Han, S.J.; Kim, J.I.; Kim, H.Y.; Park, K.M. Methionine Sulfoxide Reductase A Deficiency Exacerbates Cisplatin-Induced Nephrotoxicity via Increased Mitochondrial Damage and Renal Cell Death. Antioxid. Redox Signal. 2017, 27, 727–741. [Google Scholar] [CrossRef]
- Dirican, E.K.; Kalender, Y. Dichlorvos-induced testicular toxicity in male rats and the protective role of vitamins C and E. Exp. Toxicol. Pathol. 2012, 64, 821–830. [Google Scholar] [CrossRef]
- Li, D. Studies on the Toxicology of Trichlorfon to Rat Cortical Neuron. Master’s Thesis, Yangzhou University, Yangzhou, China, 2010. [Google Scholar]
- Kleszczyński, K.; Stepnowski, P.; Składanowski, A.C. Mechanism of cytotoxic action of perfluorinated acids: II. Disruption of mitochondrial bioenergetics. Toxicol. Appl. Pharmacol. 2009, 235, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Martinou, J.C.; Herzig, S. Mitochondrial dynamics: To be in good shape to survive. Curr. Mol. Med. 2008, 8, 131–137. [Google Scholar] [CrossRef]
- Liu, X.-z.; Zou, H.; Yuan, Y.; Bian, J.-c.; Liu, Z.-p. Effect of trichlorfon on [Ca2+]i, ATPase and GJIC of primary-cultured hepatocytes in SD rat. Chin. J. Vet. Sci. 2010, 30, 1689–1693. [Google Scholar] [CrossRef]
- Xiao, C.; He, F.-s.; Zheng, Y.-x.; Leng, S.-g.; Qin, F.-k.; Niu, Y.; Shi, Q.-l. Genetic susceptibility to intermediate myasthenia syndrome following organophosphate insecticides poisoning. Chin. J. Prev. Med. 2003, 37, 259–262. (In Chinese) [Google Scholar]
- Su, L.; Zhang, J.; Gomez, H.; Kellum, J.A.; Peng, Z. Mitochondria ROS and mitophagy in acute kidney injury. Autophagy 2023, 19, 401–414. [Google Scholar] [CrossRef]
- Zou, H. Studies on the Toxicology of Trichlorfon to Rats’ Hepatocytes. Master’s Thesis, Yangzhou University, Yangzhou, China, 2009. [Google Scholar]
- Zhu, Y.-S.; Lu, T.-Y.; Wang, R.; Huang, L.; Ma, Q.; Zhao, L.-x.; Gao, P.; Lei, X.-B.; Ni, B.-Y.; Lin, J.-L.; et al. Functional conversion of Bcl-2 into a pro-apoptotic molecule to regulate mitochondrial cytochrome c release. Chin. Bull. Life Sci. 2011, 23, 1076–1080. [Google Scholar] [CrossRef]
- Desagher, S.; Martinou, J.C. Mitochondria as the central control point of apoptosis. Trends Cell Biol. 2000, 10, 369–377. [Google Scholar] [CrossRef]
- Soltaninejad, K.; Abdollahi, M. Current opinion on the science of organophosphate pesticides and toxic stress: A systematic review. Med. Sci. Monit. 2009, 15, RA75–RA90. [Google Scholar] [CrossRef]
- Juraver-Geslin, H.A.; Durand, B.C. Early development of the neural plate: New roles for apoptosis and for one of its main effectors caspase-3. Genesis 2015, 53, 203–224. [Google Scholar] [CrossRef]
- Morgan, A.M.; Ibrahim, M.A.; Hussien, A.M. The potential protective role of Akropower against Atrazine- induced humoral immunotoxicity in rabbits. Biomed. Pharmacother. 2017, 96, 710–715. [Google Scholar] [CrossRef]
- Zou, H.; Henzel, W.J.; Liu, X.; Lutschg, A.; Wang, X. Apaf-1, a human protein homologous to C. elegans CED-4, participates in cytochrome c–dependent activation of caspase-3. Cell 1997, 90, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, M.; Gao, S.; Ren, R.; Zheng, J.; Zhang, Y. Atrazine-induced apoptosis of splenocytes in BALB/C mice. BMC Med. 2011, 9, 117. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | (%) | Componsition 3 | (%) |
|---|---|---|---|
| Corn | 71.56 | DE (MJ/kg) | 14.12 |
| Soybean meal | 15.65 | Crude protein | 16.80 |
| Fishmeal | 3.00 | Calcium | 0.67 |
| Soybean oil | 2.00 | Phosphorus | 0.56 |
| Wheat bran | 5.00 | Lysine | 0.98 |
| CaHPO4 | 0.80 | Methionine | 0.28 |
| Limestone | 0.78 | ||
| Salt | 0.35 | ||
| Lysine | 0.26 | ||
| Premix 1 | 0.50 | ||
| Choline chloride 2 | 0.10 | ||
| Total | 100.00 |
| Items | Control | Phoxim | Vitamin E + Phoxim |
|---|---|---|---|
| BUN (mmol/L) | 2.73 ± 0.04 b | 3.68 ± 0.35 a | 3.17 ± 0.09 ab |
| CRE (μmol/L) | 92.08 ± 2.62 C | 121.82 ± 3.57 A | 106.20 ± 2.13 B |
| Items | Control | Phoxim | Vitamin E + Phoxim |
|---|---|---|---|
| ROS in serum (IU/mL) | 166.09 ± 8.69 C | 289.60 ± 3.84 A | 265.01 ± 1.35 B |
| Kidney | |||
| MDA (nmol/mg protein) | 1.33 ± 0.09 B | 1.96 ± 0.16 A | 1.69 ± 0.08 AB |
| SOD (U/mg protein) | 254.39 ± 4.08 A | 203.06 ± 1.11 C | 228.06 ± 540 B |
| T-AOC (U/mg protein) | 0.43 ± 0.01 A | 0.31 ± 0.02 B | 0.36 ± 0.01 B |
| GPx (nmol/mg protein) | 31.18 ± 1.06 A | 21.8 ± 0.75 B | 26.10 ± 2.24 AB |
| CAT (U/mg protein) | 36.60 ± 2.78 A | 7.41 ± 1.55 C | 15.81 ± 0.79 B |
| GSH (U/mg protein) | 37.27 ± 3.36 A | 24.04 ± 1.10 B | 31.95 ± 1.60 AB |
| Items | Control | Phoxim | Vitamin E + Phoxim |
|---|---|---|---|
| 12 h | |||
| ROS (%) | 100.00 ± 5.70 B | 114.13 ± 2.72 A | 102.98 ± 2.05 B |
| SOD (U/mgprot) | 29.16 ± 0.22 | 25.26 ± 0.32 | 26.28 ± 1.87 |
| MDA (μmol/mg) | 9.82 ± 0.14 B | 11.38 ± 0.18 A | 11.02 ± 0.15 A |
| 24 h | |||
| ROS (%) | 100.00 ± 2.75 B | 121.83 ± 2.17 A | 107.62 ± 1.88 B |
| SOD (U/mgprot) | 26.81 ± 0.72 a | 21.94 ± 0.96 b | 23.42 ± 1.56 ab |
| MDA(μmol/mg) | 11.51 ± 0.1 B | 15.42 ± 0.59 A | 14.11 ± 0.48 A |
| Items | Control | Phoxim | Vitamin E + Phoxim |
|---|---|---|---|
| 12 h | |||
| COX (IU/L) | 360.24 ± 32.66 | 354.54 ± 7.51 | 326.82 ± 8.90 |
| Ca2+-Mg2+-ATPase (ng/L) | 338.34 ± 13.20 | 310.80 ± 11.90 | 361.36 ± 29.94 |
| SDH (U/L) | 597.06 ± 12.61 | 580.26 ± 27.89 | 589.03 ± 6.64 |
| 24 h | |||
| COX (IU/L) | 363.74 ± 14.00 a | 315.29 ± 14.09 b | 334.33 ± 3.76 ab |
| Ca2+-Mg2+-ATPase (ng/L) | 389.42 ± 4.09 A | 320.88 ± 9.41 B | 333.17 ± 16.59 AB |
| SDH (U/L) | 637.77 ± 3.83 A | 475.46 ± 11.54 B | 592.68 ± 18.03 A |
| Items | Control | Phoxim | Vitamin E + Phoxim |
|---|---|---|---|
| 12 h (%) | |||
| Cells apoptosis rate | 4.50 ± 0.12 | 8.16 ± 0.68 | 7.03 ± 1.70 |
| 24 h (%) | |||
| Cells apoptosis rate | 10.15 ± 0.49 B | 19.90 ±1.38 A | 15.24 ± 1.40 AB |
| Items | Control | Phoxim | Vitamin E + Phoxim |
|---|---|---|---|
| 12 h (%) | |||
| Mitochondrial membrane potential | 100.00 ± 5.18 a | 84.45 ± 3.02 b | 92.88 ± 3.97 ab |
| 24 h (%) | |||
| Mitochondrial membrane potential | 100.00 ± 5.44 A | 82.64 ± 2.44 B | 87.98 ± 4.20 AB |
| Items | Control | Phoxim | Vitamin E + Phoxim |
|---|---|---|---|
| Bcl-2 | 1.25 ± 0.08 A | 0.74 ± 0.09 B | 1.19 ± 0.14 AB |
| Bad | 0.82 ± 0.12 b | 1.35 ± 0.16 a | 1.12 ± 0.11 ab |
| Bax | 0.65 ± 0.07 b | 1.18 ± 0.17 a | 1.00 ± 0.14 ab |
| Caspase-3 | 0.78 ± 0.08 B | 1.22 ± 0.11 A | 1.02 ± 0.06 AB |
| Caspase-8 | 0.66 ± 0.22 b | 1.32 ± 0.05 a | 0.92 ± 0.12 ab |
| Caspase-9 | 0.80 ± 0.02 b | 1.34 ± 0.15 a | 1.32 ± 0.23 a |
| Cyto-C | 0.85 ± 0.08 | 1.16 ± 0.06 | 0.94 ± 0.16 |
| FAS | 1.13 ± 0.25 | 0.87 ± 0.09 | 0.85 ± 0.10 |
| Items | Control | Phoxim | Vitamin E + Phoxim |
|---|---|---|---|
| 12 h | |||
| Bad | 0.79 ± 0.13 B | 1.64 ± 0.14 A | 1.09 ± 0.04 B |
| Bax | 0.74 ± 0.08 B | 1.13 ± 0.05 A | 0.98 ± 0.07 AB |
| Bcl-2 | 1.38 ± 0.20 a | 0.81 ± 0.07 b | 1.07 ± 0.09 ab |
| Caspase-3 | 0.72 ± 0.25 b | 1.40 ± 0.12 a | 1.18 ± 0.10 ab |
| Caspase-8 | 1.32 ± 0.21 | 1.35 ± 0.17 | 1.02 ± 0.08 |
| Caspase-9 | 0.76 ± 0.10 B | 1.48 ± 0.23 A | 1.06 ± 0.06 AB |
| Cyto-C | 0.86 ± 0.04 b | 1.16 ± 0.08 a | 0.92 ± 0.06 b |
| 24 h | |||
| Bad | 0.90 ± 0.07 b | 1.68 ± 0.47 a | 1.01 ± 0.06 b |
| Bax | 0.72 ± 0.10 b | 1.09 ± 0.02 a | 0.97 ± 0.09 ab |
| Bcl-2 | 1.25 ± 0.05 A | 0.76 ± 0.12 B | 1.02 ± 0.05 AB |
| Caspase-3 | 0.86 ± 0.40 b | 1.75 ± 0.07 a | 1.08 ± 0.22 ab |
| Caspase-8 | 0.81 ± 0.27 | 1.82 ± 0.65 | 1.30 ± 0.23 |
| Caspase-9 | 0.79 ± 0.12 B | 2.24 ± 1.00 A | 1.05 ± 0.10 AB |
| Cyto-C | 0.88 ± 0.10 B | 1.35 ± 0.10 A | 0.97 ± 0.07 B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Sun, Y.; Song, W.; Shan, A. Vitamin E-Inhibited Phoxim-Induced Renal Oxidative Stress and Mitochondrial Apoptosis In Vivo and In Vitro of Piglets. Antioxidants 2023, 12, 2000. https://doi.org/10.3390/antiox12112000
Zhang J, Sun Y, Song W, Shan A. Vitamin E-Inhibited Phoxim-Induced Renal Oxidative Stress and Mitochondrial Apoptosis In Vivo and In Vitro of Piglets. Antioxidants. 2023; 12(11):2000. https://doi.org/10.3390/antiox12112000
Chicago/Turabian StyleZhang, Jing, Yuecheng Sun, Wentao Song, and Anshan Shan. 2023. "Vitamin E-Inhibited Phoxim-Induced Renal Oxidative Stress and Mitochondrial Apoptosis In Vivo and In Vitro of Piglets" Antioxidants 12, no. 11: 2000. https://doi.org/10.3390/antiox12112000
APA StyleZhang, J., Sun, Y., Song, W., & Shan, A. (2023). Vitamin E-Inhibited Phoxim-Induced Renal Oxidative Stress and Mitochondrial Apoptosis In Vivo and In Vitro of Piglets. Antioxidants, 12(11), 2000. https://doi.org/10.3390/antiox12112000