
Transcriptome Analysis of Cyclooctasulfur Oxidation and Reduction by the Neutrophilic Chemolithoautotrophic Sulfurovum indicum from Deep-Sea Hydrothermal Ecosystems
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Chemical Analyses
2.3. Transcriptome Analysis
2.4. Quantitative Real-Time PCR Analyses
2.5. Experiment with Dialysis Membranes to Test Cell Contact with S0 Granules
2.6. Sequence and Phylogenetic Analyses
2.7. Statistical Analysis
3. Results
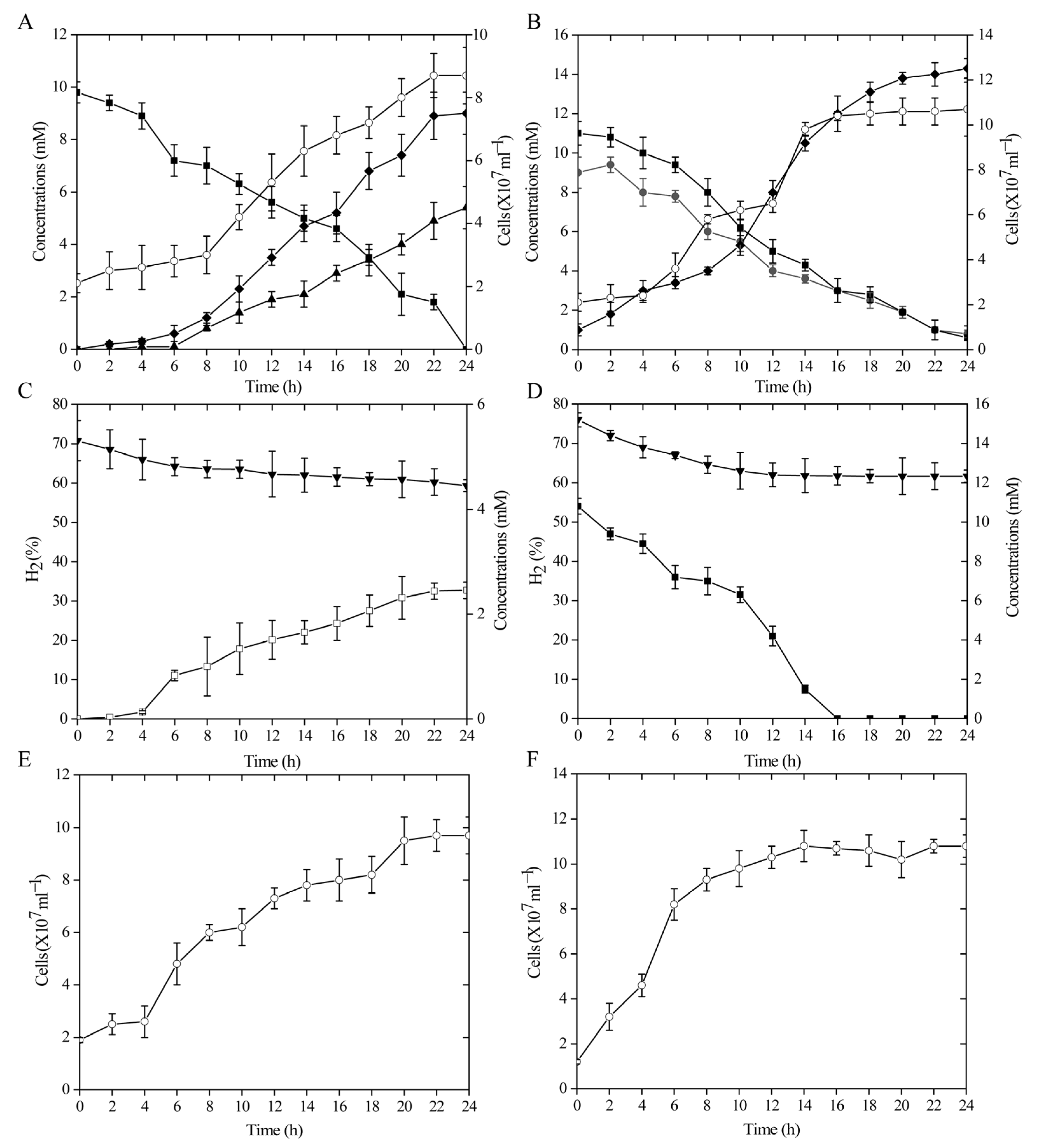
3.1. Growth Kinetics under Two Different Sulfur Oxidation Conditions
3.2. Growth Kinetics by Sulfur and Nitrate Reduction Coupled with Hydrogen Oxidization
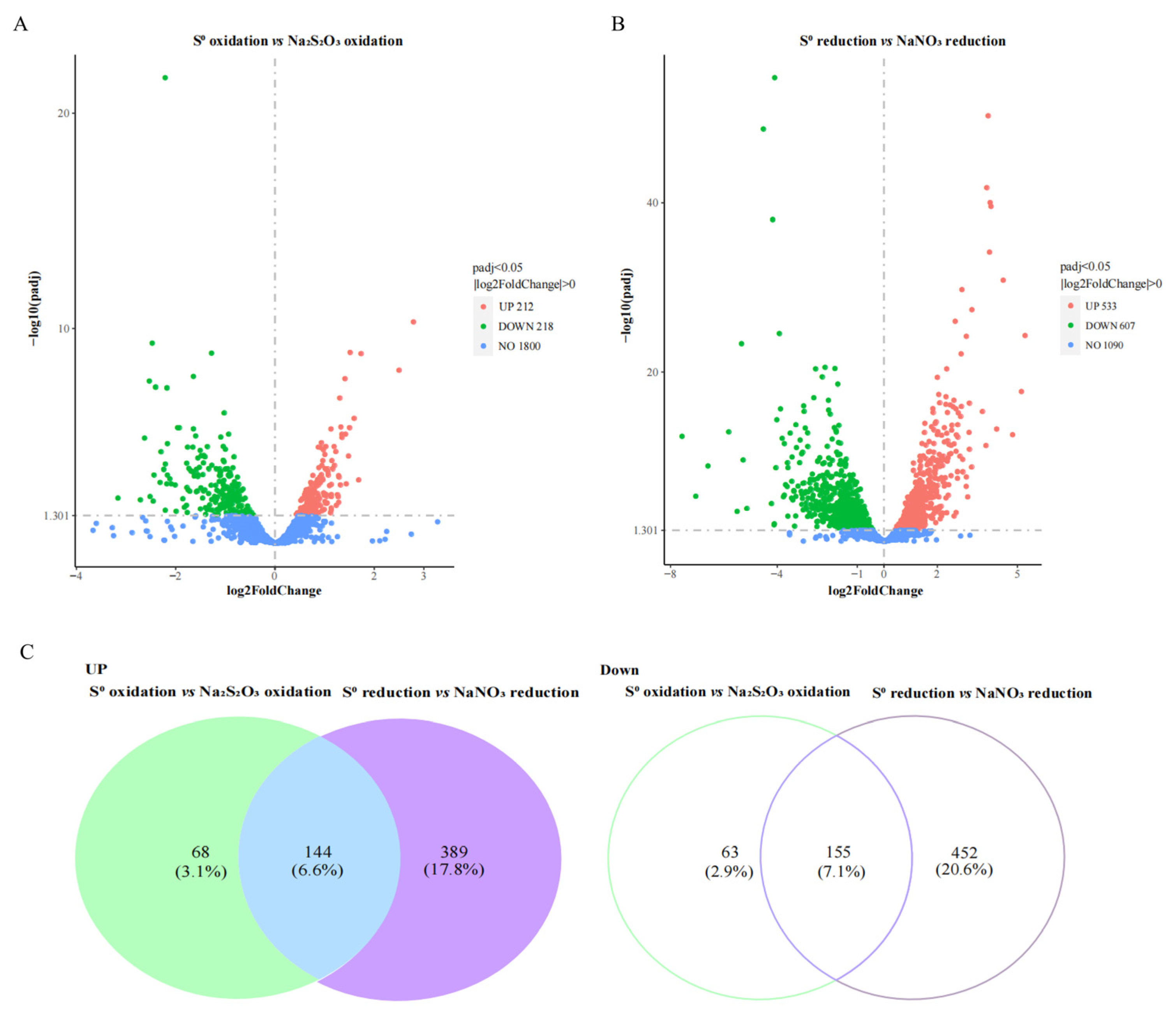
3.3. General Features of Transcriptomes for S0 Oxidation and Reduction Conditions
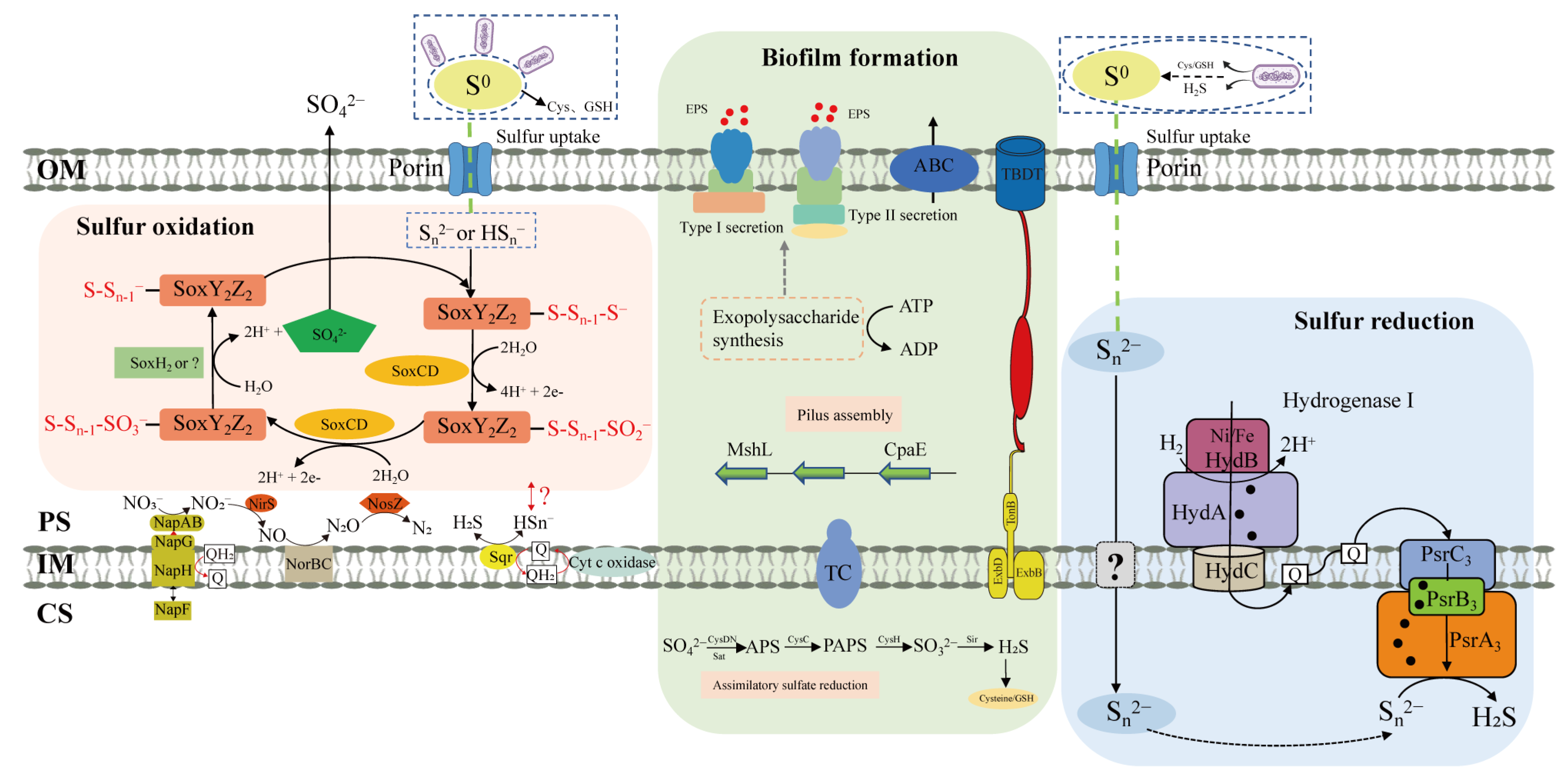
3.4. Genes Potentially Involved in S0 Activation and Their Differential Expression
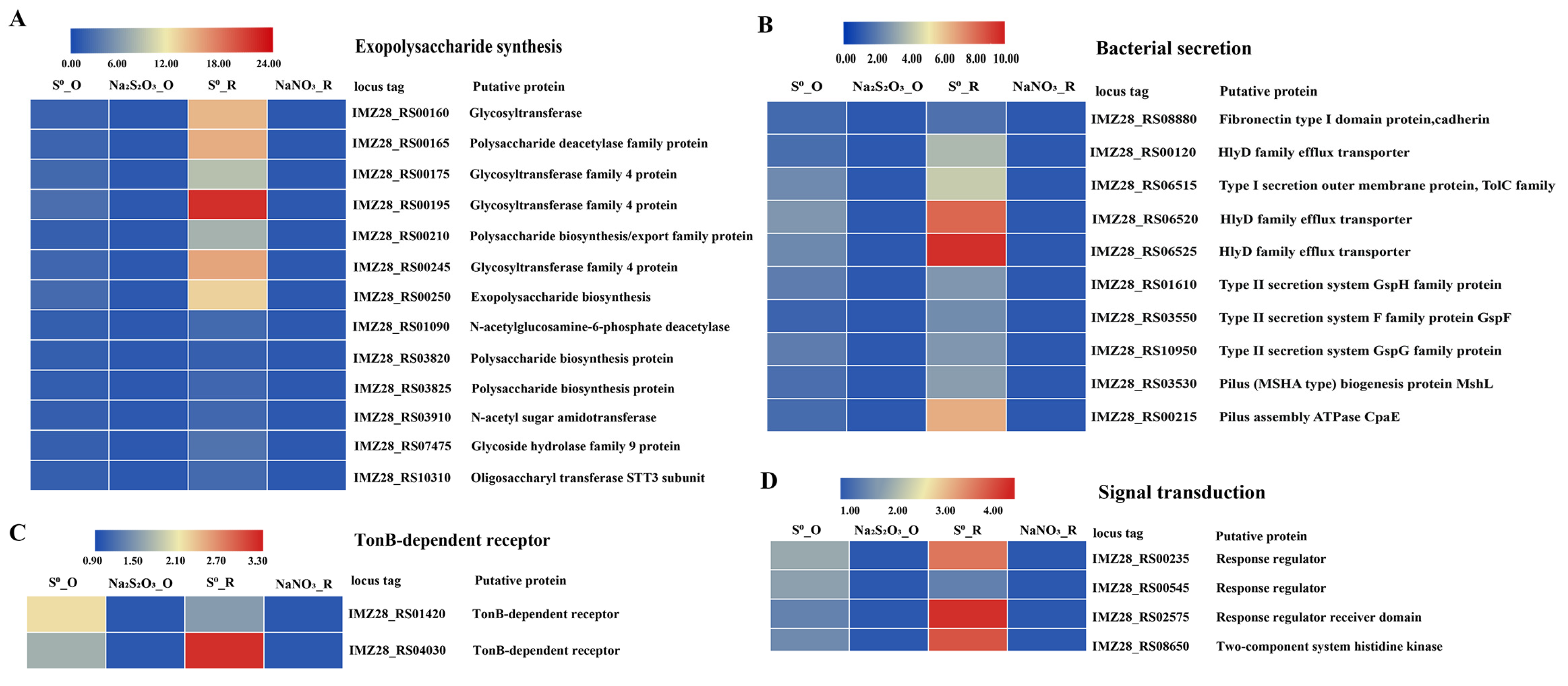
3.5. Genes Potentially Involved in S0 Uptake and Their Differential Expression
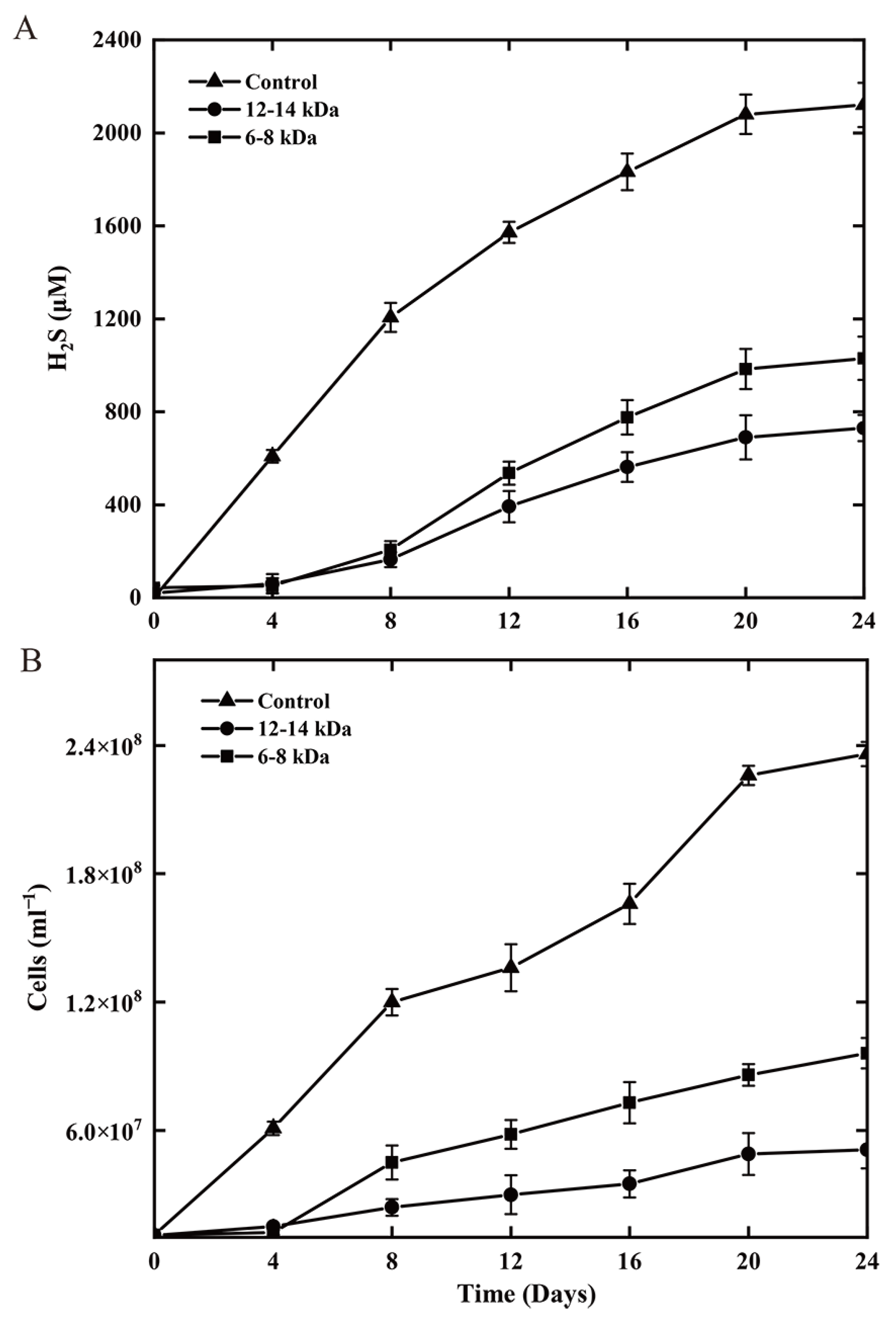
3.6. Evaluation of the Necessity for Direct Cell-S0 Contact Using Dialysis Bag Incubation
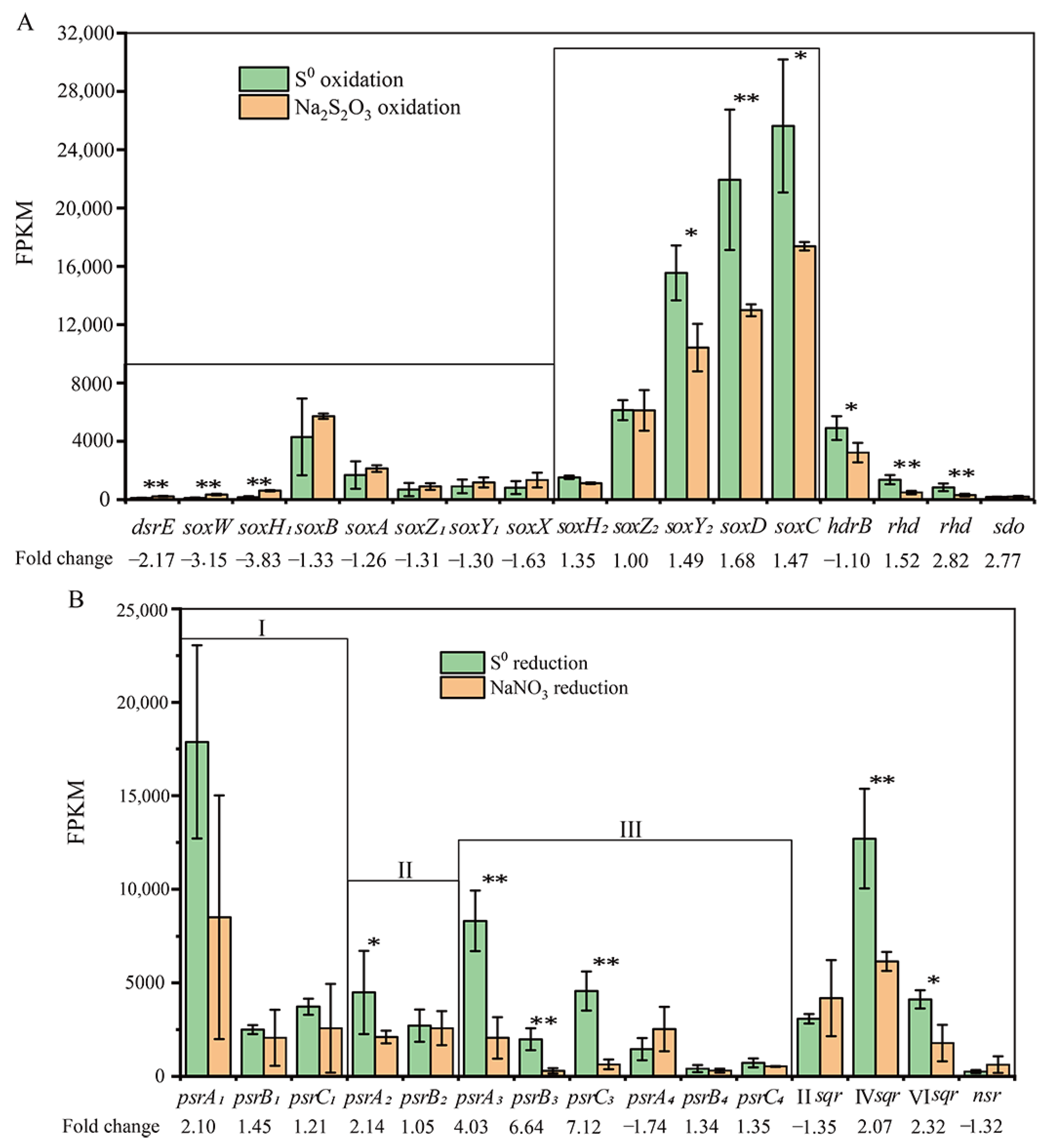
3.7. Expression of the Genes Involved in S0 Oxidation
3.8. Expression of the Genes Involved in S0 Reduction
3.9. qRT-PCR Analyses of Sulfur Oxidation and Reduction Gene Transcripts
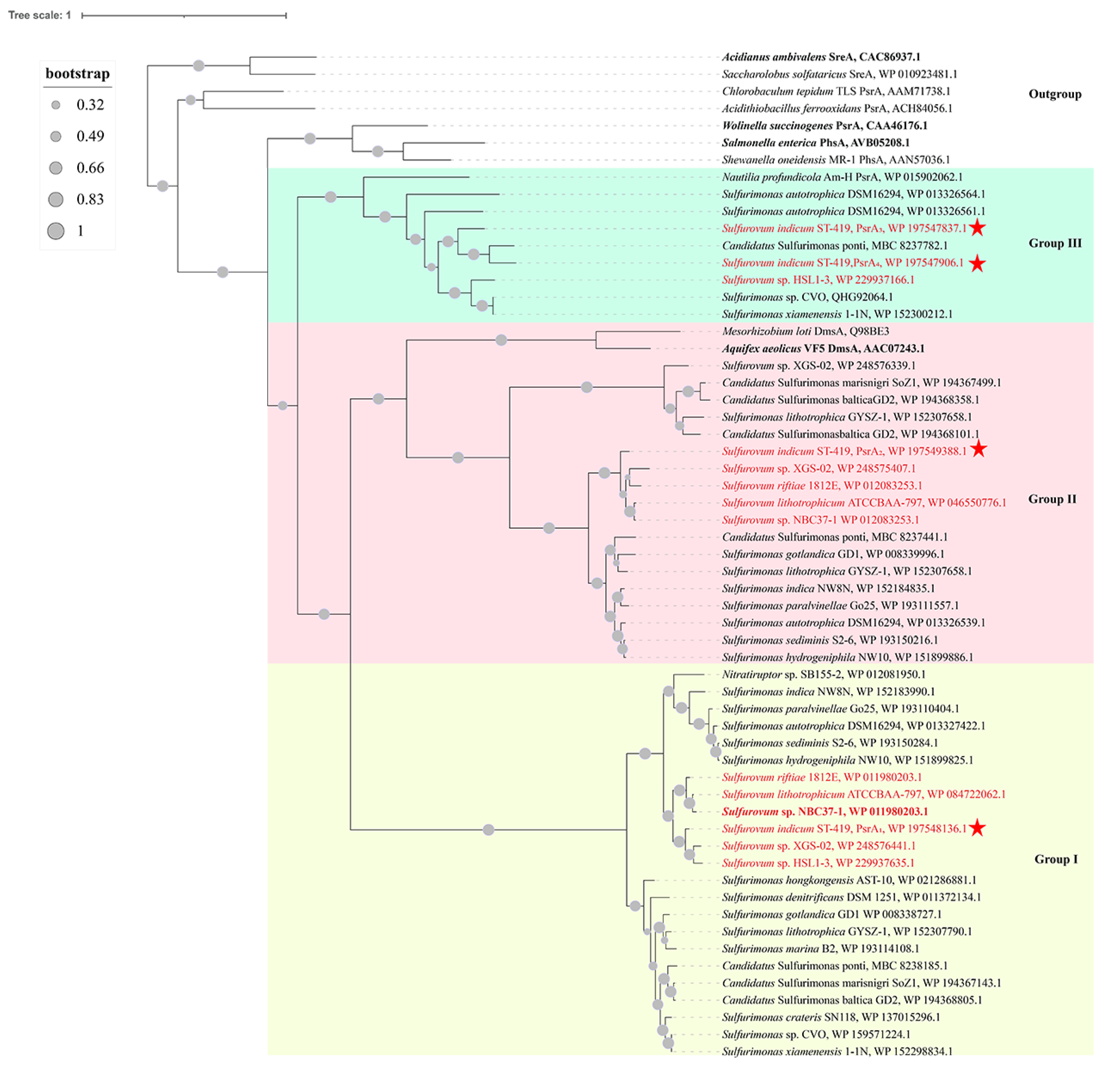
3.10. Phylogenetic and Sequence Analyses of Diverse Polysulfide Reductases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sievert, S.M.; Kiene, R.P.; Schulz-Vogt, H. The sulfur cycle. Oceanography 2007, 20, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Wasmund, K.; Mußmann, M.; Loy, A. The life sulfuric: Microbial ecology of sulfur cycling in marine sediments. Environ. Microbiol. Rep. 2017, 9, 323–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, C.G.; Bardischewsky, F.; Rother, D.; Quentmeier, A.; Fischer, J. Prokaryotic sulfur oxidation. Curr. Opin. Microbiol. 2005, 8, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Gregersen, L.H.; Bryant, D.A.; Frigaard, N.U. Mechanisms and evolution of oxidative sulfur metabolism in green sulfur bacteria. Front. Microbiol. 2011, 2, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sievert, S.M.; Vetriani, C. Chemoautotrophy at deep-sea vents: Past, present, and future. Oceanography 2012, 25, 218–233. [Google Scholar] [CrossRef]
- Meier, D.V.; Pjevac, P.; Bach, W.; Hourdez, S.; Girguis, P.R.; Vidoudez, C.; Amann, R.; Meyerdierks, A. Niche partitioning of diverse sulfur-oxidizing bacteria at hydrothermal vents. ISME J. 2017, 11, 1545–1558. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.D.; Wirsen, C.O. Microbiology and ecology of filamentous sulfur formation. Science 1997, 277, 1483–1485. [Google Scholar] [CrossRef]
- Engel, A.S.; Porter, M.L.; Stern, L.A.; Quinlan, S.; Bennett, P.C. Bacterial diversity and ecosystem function of filamentous microbial mats from aphotic (cave) sulfidic springs dominated by chemolithoautotrophic-Epsilonproteobacteria. FEMS Microbiol. Ecol. 2004, 51, 31–53. [Google Scholar] [CrossRef] [Green Version]
- Lavik, G.; Stührmann, T.; Brüchert, V.; Plas, A.V.; der Mohrholz, V.; Lam, P.; Mußmann, M.; Fuchs, B.M.; Amann, R.; Lass, U.; et al. Detoxification of sulphidic African shelf waters by blooming chemolithotrophs. Nature 2009, 457, 581. [Google Scholar] [CrossRef]
- Findlay, A.J.; Gartman, A.; MacDonald, D.J.; Hanson, T.E.; Shaw, T.J.; Luther, G.W. Distribution and size fractionation of elemental sulfur in aqueous environments: The Chesapeake Bay and mid-Atlantic ridge. Geochim. Cosmochim. Acta 2014, 142, 334–348. [Google Scholar] [CrossRef]
- Lau, G.E.; Cosmidis, J.; Grasby, S.E.; Trivedi, C.B.; Spear, J.R.; Templeton, A.S. Low-temperature formation and stabilization of rare allotropes of cyclooctasulfur (β-S8 and γ-S8) in the presence of organic carbon at a sulfur-rich glacial site in the Canadian High Arctic. Geochim. Cosmochim. Acta 2017, 200, 218–231. [Google Scholar] [CrossRef] [Green Version]
- Cai, R.; He, W.; Liu, R.; Zhang, J.; Zhang, X.; Sun, C. Deep-sea in situ insights into the formation of zero-valent sulfur driven by a bacterial thiosulfate oxidation pathway. mBio 2022, 13, e0014322. [Google Scholar] [CrossRef]
- Kafantaris, F.C.; Druschel, K.G. Kinetics of the nucleophilic dissolution of hydrophobic and hydrophilic elemental sulfur sols by sulfide. Geochim. Cosmochim. Acta 2019, 269, 554–565. [Google Scholar] [CrossRef]
- Steudel, R. Mechanism for the formation of elemental sulfur from aqueous sulfide in chemical and microbiological desulfurization processes. Ind. Eng. Chem. Res. 1996, 35, 1417–1423. [Google Scholar] [CrossRef]
- Zhang, L.; Qiu, Y.Y.; Zhou, Y.; Chen, G.H.; van Loosdrecht, M.C.M.; Jiang, F. Elemental sulfur as electron donor and/or acceptor: Mechanisms, applications and perspectives for biological water and wastewater treatment. Water. Res. 2021, 202, 117373. [Google Scholar] [CrossRef]
- Arredondo, R.; García, A.; Jerez, C. Partial removal of lipopolysaccharide from thiobacillus ferrooxidans affects its adhesion to solids. Appl. Environ. Microbiol. 1994, 6, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohwerder, T.; Sand, W. The sulfane sulfur of persulfides is the actual substrate of the sulfur-oxidizing enzymes from Acidithiobacillus and Acidiphilium spp. Microbiology 2003, 149, 1699–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangold, S.; Valdés, J.; Holmes, D.; Dopson, M. Sulfur metabolism in the extreme acidophile Acidithiobacillus caldus. Front. Microbiol. 2011, 2, 17. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Ren, Y.; Lin, J.; Liu, X.; Pang, X.; Lin, J. Acidithiobacillus caldus sulfur oxidation model based on transcriptome analysis between the wild type and sulfur oxygenase reductase defective mutant. PloS ONE 2012, 7, e39470. [Google Scholar] [CrossRef]
- Bouchal, P.; Zdrahal, Z.; Helanova, S.; Janiczek, O.; Hallberg, K.B.; Mandl, M. Proteomic and bioinformatic analysis of iron- and sulfur-oxidizing Acidithiobacillus ferrooxidans using immobilized pH gradients mass spectrometry. Proteomics 2010, 6, 4278–4285. [Google Scholar] [CrossRef]
- He, H.; Xia, J.; Huang, G.; Jiang, H.C.; Tao, X.X.; Zhao, Y.D.; He, W. Analysis of the elemental sulfur bio-oxidation by Acidithiobacillus ferrooxidans with sulfur K-edge XANES. World J. Microbiol. Biotechnol. 2010, 27, 1927–1931. [Google Scholar] [CrossRef]
- Zhan, Y.; Yang, M.; Zhang, S.; Zhao, D.; Duan, J.; Wang, W.; Yan, L. Iron and sulfur oxidation pathways of Acidithiobacillus ferrooxidans. World J. Microbiol. Biotechnol. 2019, 35, 60. [Google Scholar] [CrossRef] [PubMed]
- Boyd, E.S.; Druschel, G.K. Involvement of intermediate sulfur species in biological reduction of elemental sulfur under acidic, hydrothermal conditions. Appl. Environ. Microbiol. 2013, 79, 2061–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florentino, A.P.; Stams, A.J.; Sánchez-Andrea, I. Genome sequence of Desulfurella amilsii strain TR1 and comparative genomics of Desulfurellaceae family. Front. Microbiol. 2017, 8, 222. [Google Scholar] [CrossRef] [Green Version]
- Florentino, A.P.; Pereira, I.A.; Boeren, S.; van den Born, M.; Stams, A.J.; SánchezAndrea, I. Insight into the sulfur metabolism of Desulfurella amilsii by differential proteomics. Environ. Microbiol. 2019, 21, 209–225. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Takai, K. Sulfur metabolisms in epsilon- and gamma-proteobacteria in deep-sea hydrothermal fields. Front. Microbiol. 2011, 2, 192. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Lin, J.Q.; Liu, X.M.; Pang, X.; Zhang, C.J.; Yang, C.L.; Gao, X.Y.; Lin, C.M.; Li, Y.Q.; Li, Y. Sulfur oxidation in the acidophilic autotrophic Acidithiobacillus spp. Front. Microbiol. 2019, 9, 3290. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, R.; Xi, S.; Cai, R.; Sun, C. A novel bacterial thiosulfate oxidation pathway provides a new clue about the formation of zero-valent sulfur in deep sea. ISME J. 2020, 14, 2261–2274. [Google Scholar] [CrossRef]
- Yin, H.; Zhang, X.; Li, X.; He, Z.; Liang, Y.; Guo, X.; Hu, Q.; Xiao, Y.; Cong, J.; Ma, L. Whole-genome sequencing reveals novel insights into sulfur oxidation in the extremophile Acidithiobacillus thiooxidans. BMC Microbiol. 2014, 14, 179. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, C.G.; Rother, D.; Bardischewsky, F.; Quentmeier, A.; Fischer, J. Oxidation of reduced inorganic sulfur compounds by bacteria: Emergence of a common mechanism? Appl. Environ. Microbiol. 2001, 67, 2873–2882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabarczyk, D.B.; Berks, B.C. Intermediates in the Sox sulfur oxidation pathway are bound to a sulfane conjugate of the carrier protein SoxYZ. PloS ONE 2017, 12, e0173395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, K.; Adams, M. Sulfide dehydrogenase from the hyperthermophilic archaeon Pyrococcus furiosus: A new multifunctional enzyme involved in the reduction of elemental sulfur. J. Bacteriol. 1994, 176, 6509–6517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedderich, R.; Klimmek, O.; Kröger, A.; Dirmeier, R.; Keller, M.; Stetter, K.O. Anaerobic respiration with elemental sulfur and with disulfides. FEMS Microbiol. Rev. 1998, 22, 353–381. [Google Scholar] [CrossRef]
- Wang, S.; Jiang, L.; Hu, Q.; Liu, X.; Shao, Z. Elemental sulfur reduction by a deep-sea hydrothermal vent campylobacterium Sulfurimonas sp. NW10. Environ. Microbiol. 2021, 23, 965–979. [Google Scholar] [CrossRef]
- Takai, K.; Inagaki, F.; Nakagawa, S.; Hirayama, H.; Nunoura, T.; Sako, Y.; Nealson, K.H.; Horikoshi, K. Isolation and phylogenetic diversity of members of previously uncultivated ε-Proteobacteria in deep-sea hydrothermal fields. FEMS Microbiol. Lett. 2003, 218, 167–174. [Google Scholar] [PubMed] [Green Version]
- Vetriani, C.; Voordeckers, J.W.; Crespo-Medina, M.; O’Brien, C.E.; Giovannelli, D.; Lutz, R.A. Deep-sea hydrothermal vent Epsilonproteobacteria encode a conserved and widespread nitrate reduction pathway (Nap). ISME J. 2014, 8, 1510–1521. [Google Scholar] [CrossRef]
- Dick, G.J. The microbiomes of deep-sea hydrothermal vents: Distributed globally, shaped locally. Nat. Rev. Microbiol. 2019, 17, 271–283. [Google Scholar] [CrossRef]
- Inagaki, F.; Takai, K.; Nealson, K.H.; Horikoshi, K. Sulfurovum lithotrophicum gen. nov., sp. nov., a novel sulfur-oxidizing chemolithoautotroph within the ε-Proteobacteria isolated from Okinawa trough hydrothermal sediments. Int. J. Syst. Evol. Microbiol. 2004, 54, 1477–1482. [Google Scholar] [CrossRef] [Green Version]
- Reveillaud, J.; Reddington, E.; McDermott, J.; Algar, C.; Meyer, J.L.; Sylva, S.; Seewald, J.; German, C.R.; Huber, J.A. Subseafloor microbial communities in hydrogen-rich vent fluids from hydrothermal systems along the Mid-Cayman Rise. Environ. Microbiol. 2016, 18, 1970–1987. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.E.; Reveillaud, J.; Reddington, E.; Delmont, T.O.; Eren, A.M.; McDermott, J.M.; Seewald, J.S.; Huber, J.A. Genomic variation in microbial populations inhabiting the marine subseafloor at deep-sea hydrothermal vents. Nat. Commun. 2017, 8, 1114. [Google Scholar] [CrossRef] [Green Version]
- Watsuji, T.O.; Nakagawa, S.; Tsuchida, S.; Toki, T.; Hirota, A.; Tsunogai, U.; Takai, K. Diversity and function of epibiotic microbial communities on the galatheid crab, Shinkaia crosnieri. Microbes. Environ. 2010, 25, 288–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pjevac, P.; Kamyshny, A.; Dyksma, S.; Mußmann, M. Microbial consumption of zero-valence sulfur in marine benthic habitats. Environ. Microbiol. 2014, 16, 3416–3430. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Wang, S.; Li, D.; Shao, Z.; Lai, Q.; Wang, Y.; Wei, M.; Han, X.; Jiang, L. Sulfurovum indicum sp. nov. a novel hydrogen- and sulfur-oxidizing chemolithoautotroph isolated from a deep-sea hydrothermal plume in the northwestern indian ocean. Int. J. Syst. Evol. Microbiol. 2021, 71, 004748. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in realtime RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Lahme, S.; Enning, D.; Callbeck, C.M.; Vega, D.M.; Curtis, T.P.; Head, I.M.; Hubert, C.R.J. Metabolites of an oil field sulfide-oxidizing, nitrate-reducing Sulfurimonas sp. cause severe corrosion. Appl. Environ. Microbiol. 2019, 85, e01891-18. [Google Scholar] [CrossRef] [Green Version]
- Callbeck, C.M.; Lavik, G.; Ferdelman, T.G.; Fuchs, B.; Gruber-Vodicka, H.R.; Hach, P.F.; Littmann, S.; Schoffelen, N.J.; Kalvelage, T.; Thomsen, S.; et al. Oxygen minimum zone cryptic sulfur cycling sustained by offshore transport of key sulfur oxidizing bacteria. Nat. Commun. 2018, 9, 1729. [Google Scholar] [CrossRef] [Green Version]
- Boyd, E.S.; Jackson, R.A.; Encarnacion, G.; Zahn, J.A.; Beard, T.; Leavitt, W.D.; Pi, Y.; Zhang, C.L.; Pearson, A.; Geesey, G.G. Isolation, characterization, and ecology of sulfur-respiring crenarchaea inhabiting acid-sulfate-chloride-containing geothermal springs in Yellowstone National Park. Appl. Environ. Microbiol. 2007, 73, 6669–6677. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.; Sha, Z.L.; Sun, C.M. Formation of cadmium sulfide nanoparticles mediates cadmium resistance and light utilization of the deep-sea bacterium Idiomarina sp. OT37-5b. Environ. Microbiol. 2021, 23, 934–948. [Google Scholar] [CrossRef]
- Amenabar, M.J.; Boyd, E.S. Mechanisms of mineral substrate acquisition in a thermoacidophile. Appl. Environ. Microbiol. 2018, 84, e00334-18. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2002, 2, 3. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boratyn, G.M.; Schäffer, A.A.; Agarwala, R.; Altschul, S.F.; Lipman, D.J.; Madden, T.L. Domain enhanced lookup time accelerated BLAST. Biol. Direct 2012, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Slobodkina, G.B.; Mardanov, A.V.; Ravin, N.V.; Frolova, A.A.; Chernyh, N.A.; Bonch-Osmolovskaya, E.A.; Slobodkin, A.I. Respiratory ammonification of nitrate coupled to anaerobic oxidation of elemental sulfur in deep-sea autotrophic thermophilic bacteria. Front. Microbiol. 2017, 8, 87. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Bogino, P.C.; Oliva, M.d.L.; Sorroche, F.G.; Giordano, W. The role of bacterial biofilms and surface components in plant-cacterial associations. Int. J. Mol. Sci. 2013, 14, 15838–15859. [Google Scholar] [CrossRef] [Green Version]
- Vozza, N.F.; Abdian, P.L.; Russo, D.M.; Mongiardini, E.J.; Lodeiro, A.R.; Molin, S.; Zorreguieta, A. A rhizobium leguminosarum CHDL- (cadherin-like-) lectin participates in assembly and remodeling of the biofilm matrix. Front. Microbiol. 2016, 7, 1608. [Google Scholar] [CrossRef] [Green Version]
- Craig, L.; Forest, K.T.; Maier, B. Type IV Pili: Dynamics, biophysics and functional consequences. Nat. Rev. Microbiol. 2019, 17, 429–440. [Google Scholar] [CrossRef]
- Sandeep, T.; Hancock, R.E.W. Involvement of two related porins, OprD and OpdP, in the uptake of arginine by Pseudomonas aeruginosa. FEMS Microbiol. Lett. 2006, 260, 23–29. [Google Scholar]
- Tommassen, J. Assembly of outer-membrane proteins in bacteria and mitochondria. Microbiology. 2010, 156, 2587–2596. [Google Scholar] [CrossRef] [Green Version]
- Götz, F.; Pjevac, P.; Markert, S.; McNichol, J.; Becher, D.; Schweder, T.; Mussmann, M.; Sievert, S.M. Transcriptomic and proteomic insight into the mechanism of cyclooctasulfur- versus thiosulfate-oxidation by the chemolithoautotroph Sulfurimonas denitrificans. Environ. Microbiol. 2019, 21, 244–258. [Google Scholar] [CrossRef] [Green Version]
- Buonfiglio, V.; Polidoro, M.; Flora, L.; Citro, G.; Valenti, P.; Orsi, N. Identification of two outer membrane proteins involved in the oxidation of sulphur compounds in Thiobacillus ferrooxidans. FEMS Microbiol. Rev. 1993, 11, 43–50. [Google Scholar] [CrossRef]
- Lahme, S.; Callbeck, C.M.; Eland, L.E.; Wipat, A.; Enning, D.; Head, I.M.; Hubert, C.R.J. Comparison of sulfide-oxidizing Sulfurimonas strains reveals a new mode of thiosulfate formation in subsurface environments. Environ. Microbiol. 2020, 22, 1784–1800. [Google Scholar] [CrossRef] [PubMed]
- Heide, V.D.T.; Poolman, B. ABC transporters: One, two or four extracytoplasmic substrate-binding sites? EMBO Rep. 2002, 3, 938–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulligan, C.; Fischer, M.; Thomas, G.H. Tripartite ATP-independent periplasmic (TRAP) transporters in bacteria and archaea. FEMS Microbiol. Rev. 2011, 35, 68–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagchi, A.; Ghosh, T.C. Structural insight into the interactions of SoxV, SoxW and SoxS in the process of transport of reductants during sulfur oxidation by the novel global sulfur oxidation reaction cycle. Biophys. Chem. 2006, 119, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Chen, H.; Ao, J.; Zhou, H.; Chen, X. Identification of differentially expressed genes in Sulfobacillus sp. TPY grown on either elemental sulphur or Fe2+. J. Gen. Appl. Microbiol. 2010, 56, 389–397. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Jiang, L.; Hu, Q.; Cui, L.; Zhu, B.; Fu, X.; Lai, Q.; Shao, Z.; Yang, S. Characterization of Sulfurimonas hydrogeniphila sp. nov., a novel bacterium predominant in deep-sea hydrothermal vents and comparative genomic analyses of the genus Sulfurimonas. Front. Microbiol. 2021, 12, 626705. [Google Scholar] [CrossRef]
- Yamamoto, M.; Nakagawa, S.; Shimamura, S.; Takai, K.; Horikoshi, K. Molecular characterization of inorganic sulfur-compound metabolism in the deep-sea epsilonproteobacterium Sulfurovum sp. NBC37-1. Environ. Microbiol. 2010, 12, 1144–1153. [Google Scholar] [CrossRef]
- Jelen, B.; Giovannelli, D.; Falkowski, P.G.; Vetriani, C. Elemental sulfur reduction in the deep-sea vent thermophile, Thermovibrio ammonificans. Environ. Microbiol. 2018, 20, 2301–2316. [Google Scholar] [CrossRef]
- Krafft, T.; Bokranz, M.; Klimmek, O.; Schröder, I.; Fahrenholz, F.; Kojro, E.; Kröger, A. Cloning and nucleotide sequence of the psrA gene of Wolinella succinogenes polysulphide reductase. Eur. J. Biochem. 1992, 206, 503–510. [Google Scholar] [CrossRef]
- Hinsley, A.P.; Berks, B.C. Specificity of respiratory pathways involved in the reduction of sulfur compounds by Salmonella enterica. Microbiology 2002, 148, 3631–3638. [Google Scholar] [CrossRef] [Green Version]
- Laska, S.; Lottspeich, F.; Kletzin, A. Membrane-bound hydrogenase and sulfur reductase of the hyperthermophilic and acidophilic archaeon Acidianus ambivalens. Microbiology 2003, 149, 2357–2371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guiral, M.; Tron, P.; Aubert, C.; Gloter, A.; Iobbi-Nivol, C.; Giudici-Orticoni, M.T. A membrane-bound multienzyme, hydrogen-oxidizing, and sulfur-reducing complex from the hyperthermophilic bacterium Aquifex aeolicus. J. Biol. Chem. 2005, 280, 42004–42015. [Google Scholar] [CrossRef] [Green Version]
- Takai, K.; Campbell, B.J.; Cary, S.C.; Suzuki, M.; Oida, H.; Nunoura, T.; Hirayama, H.; Nakagawa, S.; Suzuki, Y.; Inagaki, F.; et al. Enzymatic and genetic characterization of carbon and energy metabolisms by deep-sea hydrothermal chemolithoautotrophic isolates of Epsilonproteobacteria. Appl. Environ. Microbiol. 2005, 71, 7310–7320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Jiang, L.; Xie, S.; Alain, K.; Wang, Z.; Wang, J.; Liu, D.; Shao, Z. Disproportionation of inorganic sulfur compounds by mesophilic chemolithoautotrophic Campylobacterota. Msystems 2022, 21, e0095422. [Google Scholar] [CrossRef] [PubMed]
- Franz, B.; Lichtenberg, H.; Hormes, J.; Modrow, H.; Dahl, C.; Prange, A. Utilization of solid ‘elemental’ sulfur by the phototrophic purple sulfur bacterium Allochromatium vinosum: A sulfur K-edge X-ray absorption spectroscopy study. Microbiology 2007, 153, 1268–1274. [Google Scholar] [CrossRef] [Green Version]
- Blumentals, I.I.; Itoh, M.; Olson, G.J.; Kelly, R.M. Role of polysulfides in reduction of elemental sulfur by the hyperthermophilic archaebacterium Pyrococcus furiosus. Appl. Environ. Microbiol. 1990, 56, 1255–1262. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Pedersen, K.; Edlund, J.; Eriksson, L.; Åström, M.; Andersson, A.F.; Bertilsson, S.; Dopson, M. Potential for hydrogen-oxidizing chemolithoautotrophic and diazotrophic populations to initiate biofilm formation in oligotrophic, deep terrestrial subsurface waters. Microbiome 2017, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Kerou, M.; Offre, P.; Valledor, L.; Abby, S.S.; Melcher, M.; Nagler, M.; Weckwerth, W.; Schleper, C. Proteomics and comparative genomics of Nitrososphaera viennensis reveal the core genome and adaptations of archaeal ammonia oxidizers. Proc. Natl. Acad. Sci. USA 2016, 113, E7937–E7946. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Bellenberg, S.; Neu, T.R.; Sand, W.; Vera, M. The Biofilm Lifestyle of Acidophilic Metal/Sulfur-Oxidizing Microorganisms. In Biotechnology of Extremophiles: Grand Challenges in Biology and Biotechnology; Rampelotto, P., Ed.; Springer: Cham, Switzerland, 2016; Volume 1. [Google Scholar]
- Zhang, R.; Neu, T.R.; Li, Q.; Blanchard, V.; Zhang, Y.; Schippers, A.; Sand, W. Insight into interactions of thermoacidophilic archaea with elemental sulfur: Biofilm dynamics and EPS analysis. Front. Microbiol. 2019, 10, 896. [Google Scholar] [CrossRef] [Green Version]
- Svensson, S.L.; Pryjma, M.; Gaynor, E.C. Flagella-mediated adhesion and extracellular DNA release contribute to biofilm formation and stress tolerance of Campylobacter jejuni. PLoS ONE 2014, 9, e106063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarrell, K.F.; Stark, M.; Nair, D.B.; Chong, J.P.J. Flagella and pili are both necessary for efficient attachment of Methanococcus maripaludis to surfaces. FEMS Microbiol. Lett. 2011, 319, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Harneit, K.G.; Ksel, A.; Kock, D.; Klock, J.H.; Gehrke, T.; Sand, W. Adhesion to metal sulfide surfaces by cells of Acidithiobacillus ferrooxidans, Acidithiobacillus thiooxidans and Leptospirillum ferrooxidans. Hydrometallurgy 2006, 83, 245–254. [Google Scholar] [CrossRef]
- Zhu, H.; Tian, B.; Liu, W.; Zhang, S.; Cao, C.; Zhang, Y.; Zou, W. A three-stage culture process for improved exopolysaccharide production by tremella fuciformis. Bioresour. Technol. 2012, 116, 526–528. [Google Scholar] [CrossRef]
- Manchur, M.A.; Kikumoto, M.; Kanao, T.; Takada, J.; Kamimura, K. Characterization of an OmpA-like outer membrane protein of the acidophilic iron-oxidizing bacterium, Acidithiobacillus ferrooxidans. Extremophiles 2011, 15, 403–410. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, P.; Guiliani, N.; Valenzuela, L.; Beard, S.; Jerez, C.A. Differential protein expression during growth of Acidithiobacillus ferrooxidans on ferrous iron, sulfur compounds, or metal sulfides. Appl. Environ. Microbiol. 2004, 70, 4491–4498. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.; Sun, C. Cadmium sulfide nanoparticle biomineralization and biofilm formation mediate cadmium resistance of the deep-sea bacterium Pseudoalteromonas sp. MT33b. Environ. Microbiol. Rep. 2021, 13, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Ares, A.; Sakai, S.; Sasaki, T.; Mitarai, S.; Nunoura, T. Sequestration and efflux largely account for Cd and Cu resistance in the deep sea Epsilonproteobacterium, Nitratiruptor sp. SB155-2. Environ. Microbiol. 2021, 16255, 1462–2920. [Google Scholar]
- Quatrini, R.; Jedlicki, E.; Holmes, D.S. Genomic insights into the iron uptake mechanisms of the biomining microorganism Acidithiobacillus ferrooxidans. J. Ind. Microbiol. Biotechnol. 2005, 32, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Hensen, D.; Sperling, D.; Trüper, H.G.; Brune, D.C.; Dahl, C. Thiosulphate oxidation in the phototrophic sulphur bacterium Allochromatium vinosum. Mol. Microbiol. 2006, 62, 794–810. [Google Scholar] [CrossRef]
- Pjevac, P.; Meier, D.V.; Markert, S.; Hentschker, C.; Schweder, T.; Becher, D.; Gruber-Vodicka, H.R.; Richter, M.; Bach, W.; Amann, R.; et al. Metaproteogenomic profiling of microbial communities colonizing actively venting hydrothermal chimneys. Front. Microbiol. 2018, 9, 680. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Jiang, L.; Cui, L.; Alain, K.; Xie, S.; Shao, Z. Transcriptome Analysis of Cyclooctasulfur Oxidation and Reduction by the Neutrophilic Chemolithoautotrophic Sulfurovum indicum from Deep-Sea Hydrothermal Ecosystems. Antioxidants 2023, 12, 627. https://doi.org/10.3390/antiox12030627
Wang S, Jiang L, Cui L, Alain K, Xie S, Shao Z. Transcriptome Analysis of Cyclooctasulfur Oxidation and Reduction by the Neutrophilic Chemolithoautotrophic Sulfurovum indicum from Deep-Sea Hydrothermal Ecosystems. Antioxidants. 2023; 12(3):627. https://doi.org/10.3390/antiox12030627
Chicago/Turabian StyleWang, Shasha, Lijing Jiang, Liang Cui, Karine Alain, Shaobin Xie, and Zongze Shao. 2023. "Transcriptome Analysis of Cyclooctasulfur Oxidation and Reduction by the Neutrophilic Chemolithoautotrophic Sulfurovum indicum from Deep-Sea Hydrothermal Ecosystems" Antioxidants 12, no. 3: 627. https://doi.org/10.3390/antiox12030627
APA StyleWang, S., Jiang, L., Cui, L., Alain, K., Xie, S., & Shao, Z. (2023). Transcriptome Analysis of Cyclooctasulfur Oxidation and Reduction by the Neutrophilic Chemolithoautotrophic Sulfurovum indicum from Deep-Sea Hydrothermal Ecosystems. Antioxidants, 12(3), 627. https://doi.org/10.3390/antiox12030627