

A Study on the Planarian Model Confirms the Antioxidant Properties of Tameron against X-ray- and Menadione-Induced Oxidative Stress


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Computer-Assisted Morphometry In Vivo
2.3. Whole-Mount Immunocytochemical Study of Planarian Stem Cell Mitotic Activity
2.4. Experimental Testing Substances
2.5. Planarian X-ray Irradiation
2.6. RT-PCR for Gene Expression Analysis
2.7. Cellular Antioxidant Activity (CAA) Assay in Planarians
2.8. Tameron-Mediated H2DCFDA Oxidation In Vitro
2.9. Statistical Data Analysis
2.10. Ethical Standards
3. Results
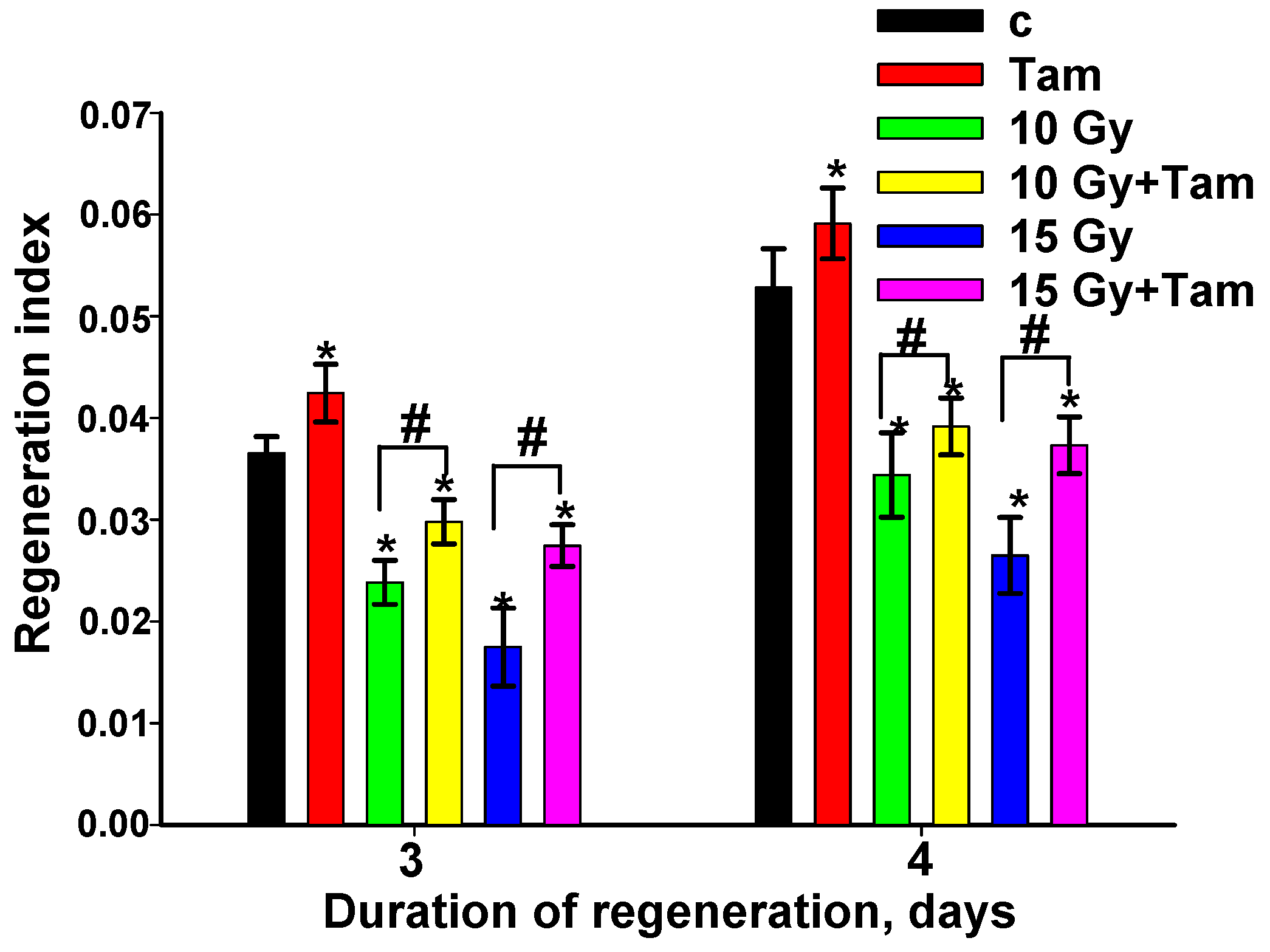
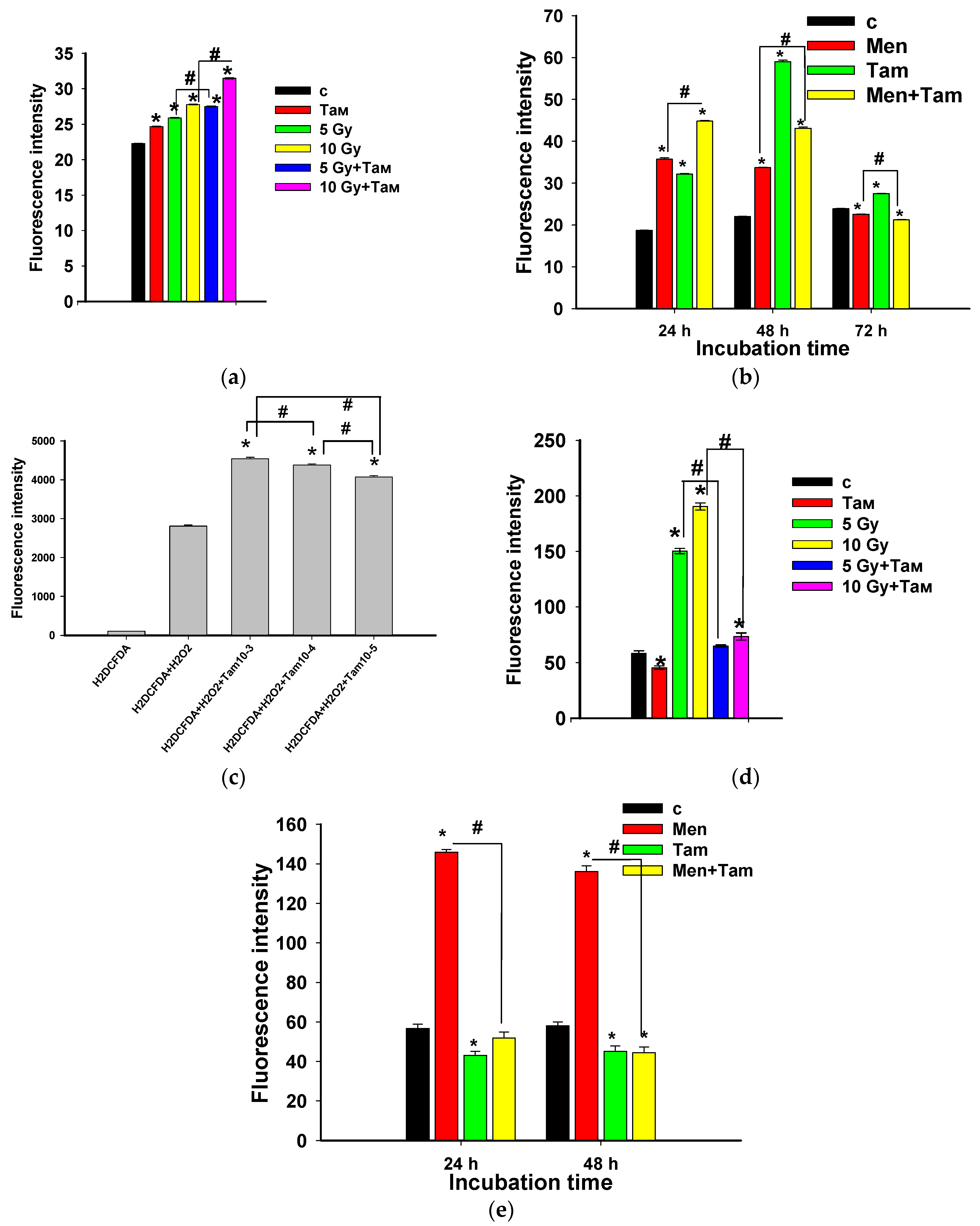
3.1. Tameron Acts as Radioprotector, Boosting the Planarian Regeneration
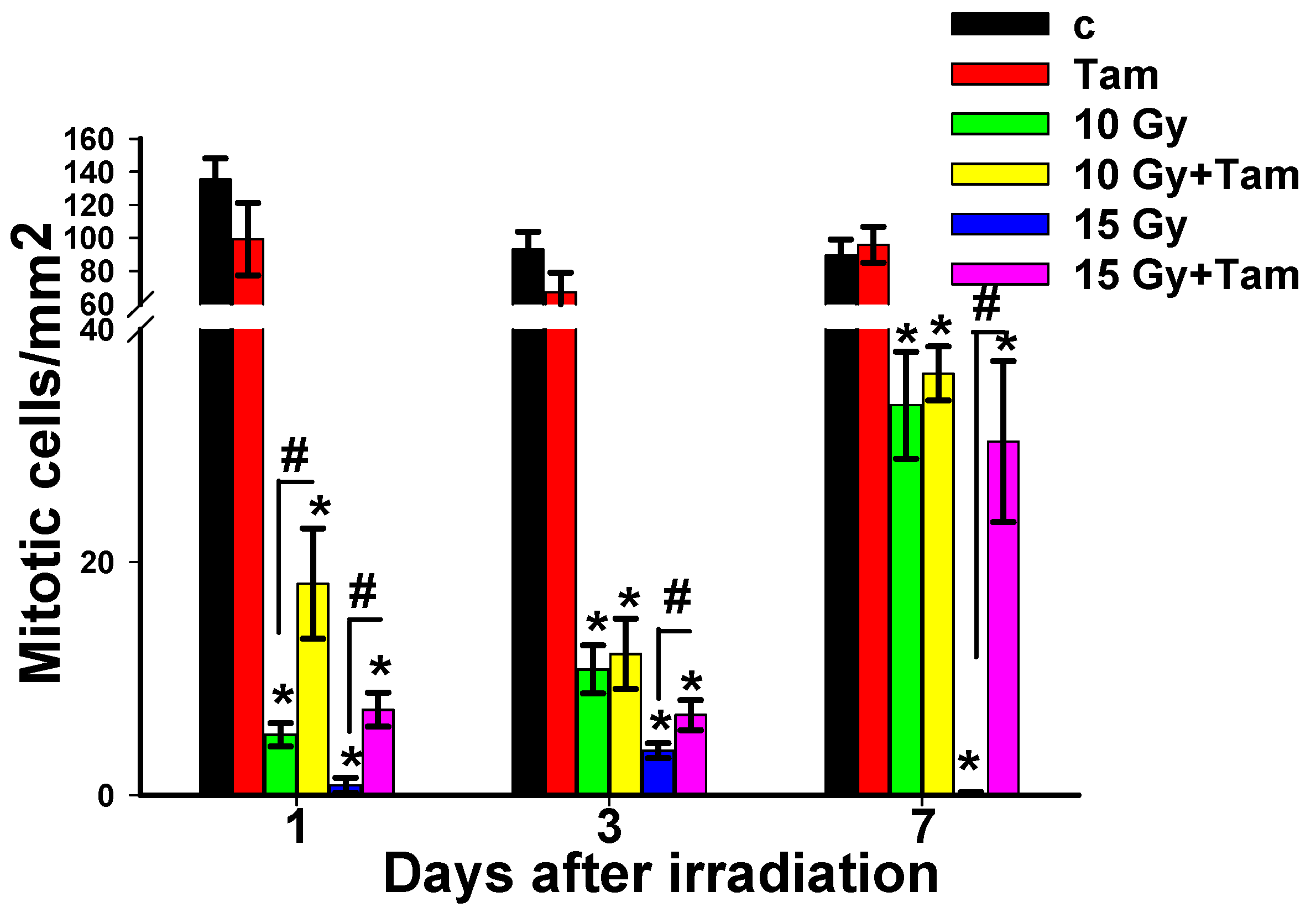
3.2. Tameron Increases the Survival of Mitotic-Active Planarian Neoblasts after X-ray Irradiation
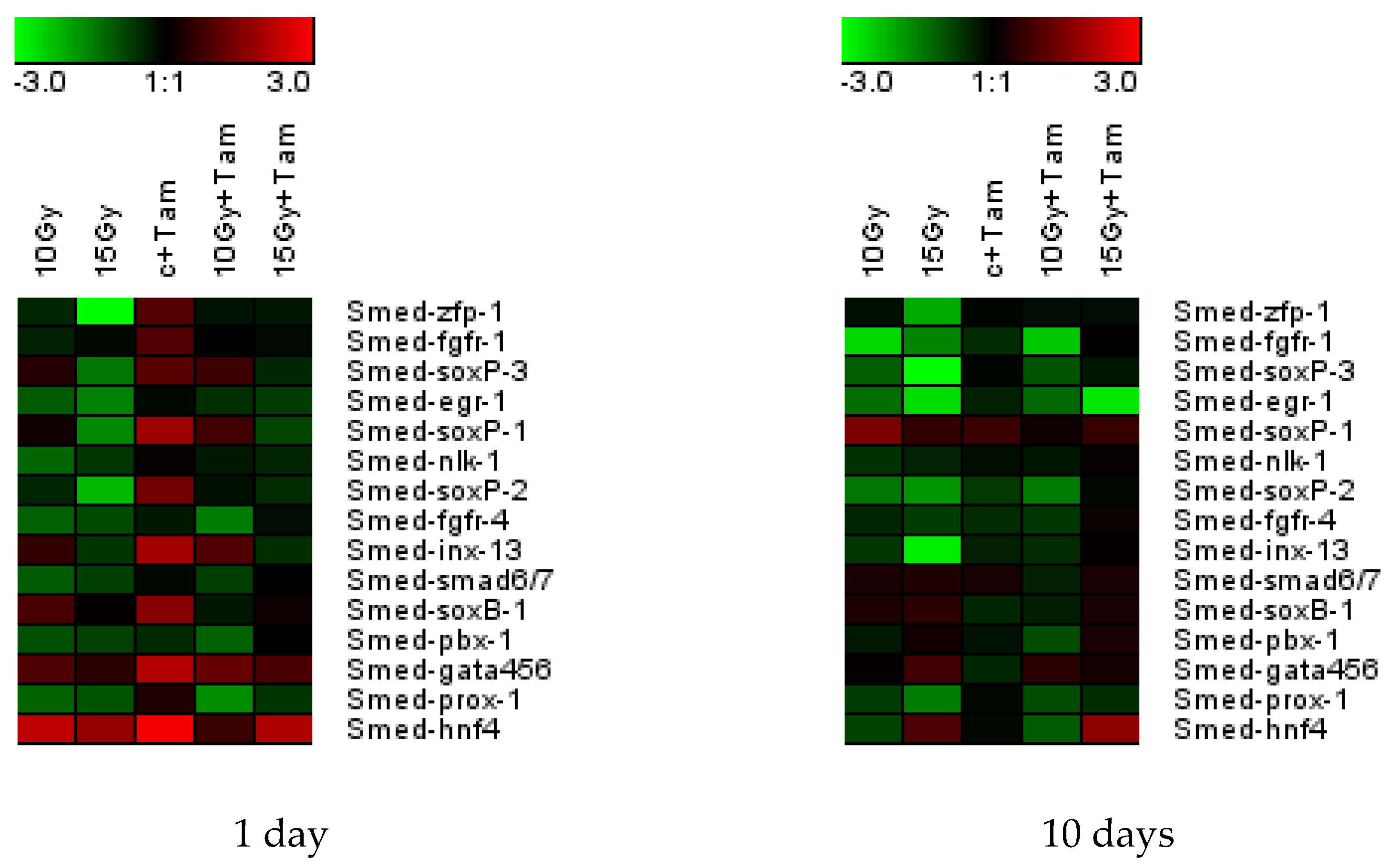
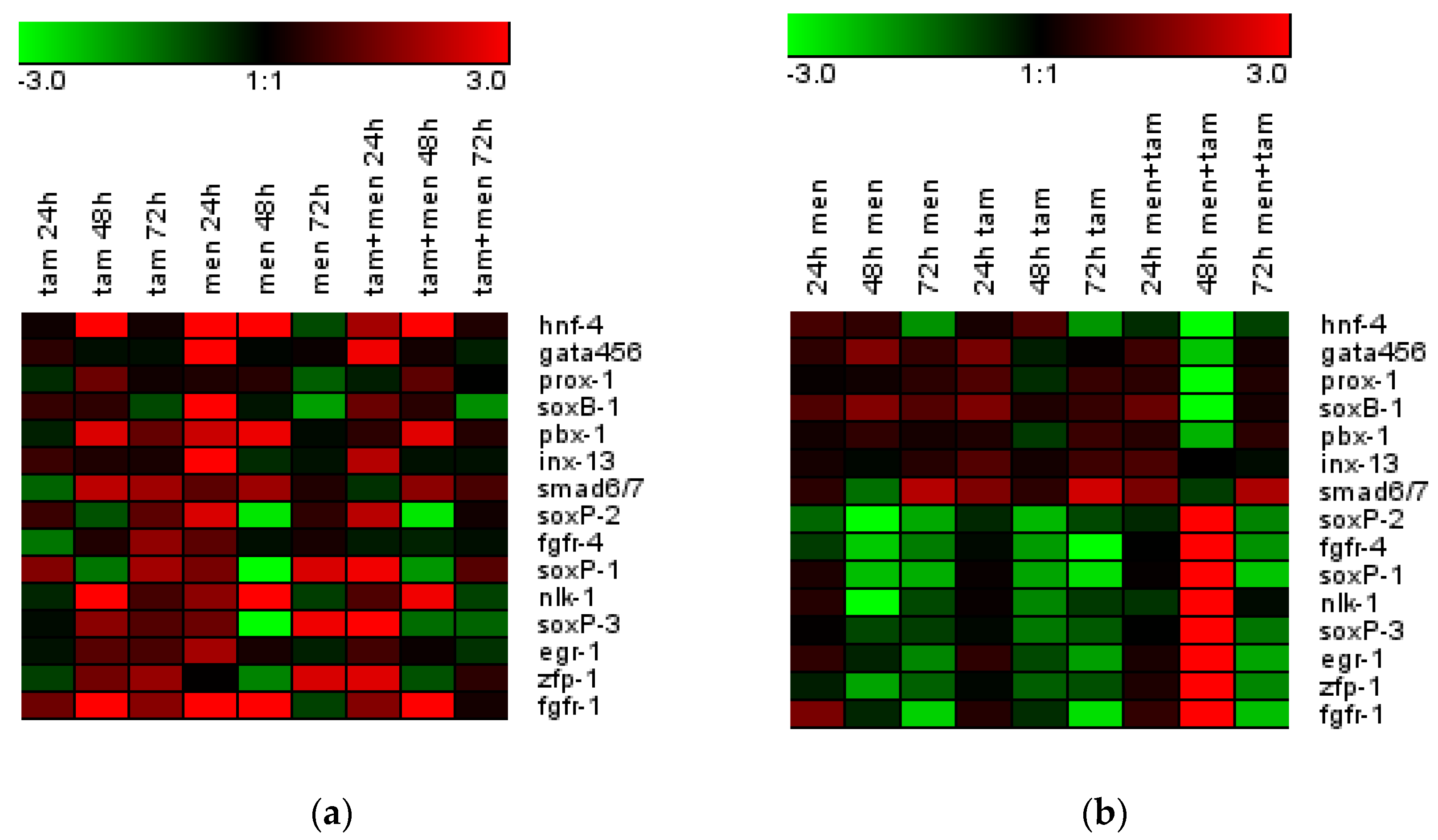
3.3. Tameron Saves the Expression of Neoblast Marker Genes after Sublethal Doses of X-ray Irradiation
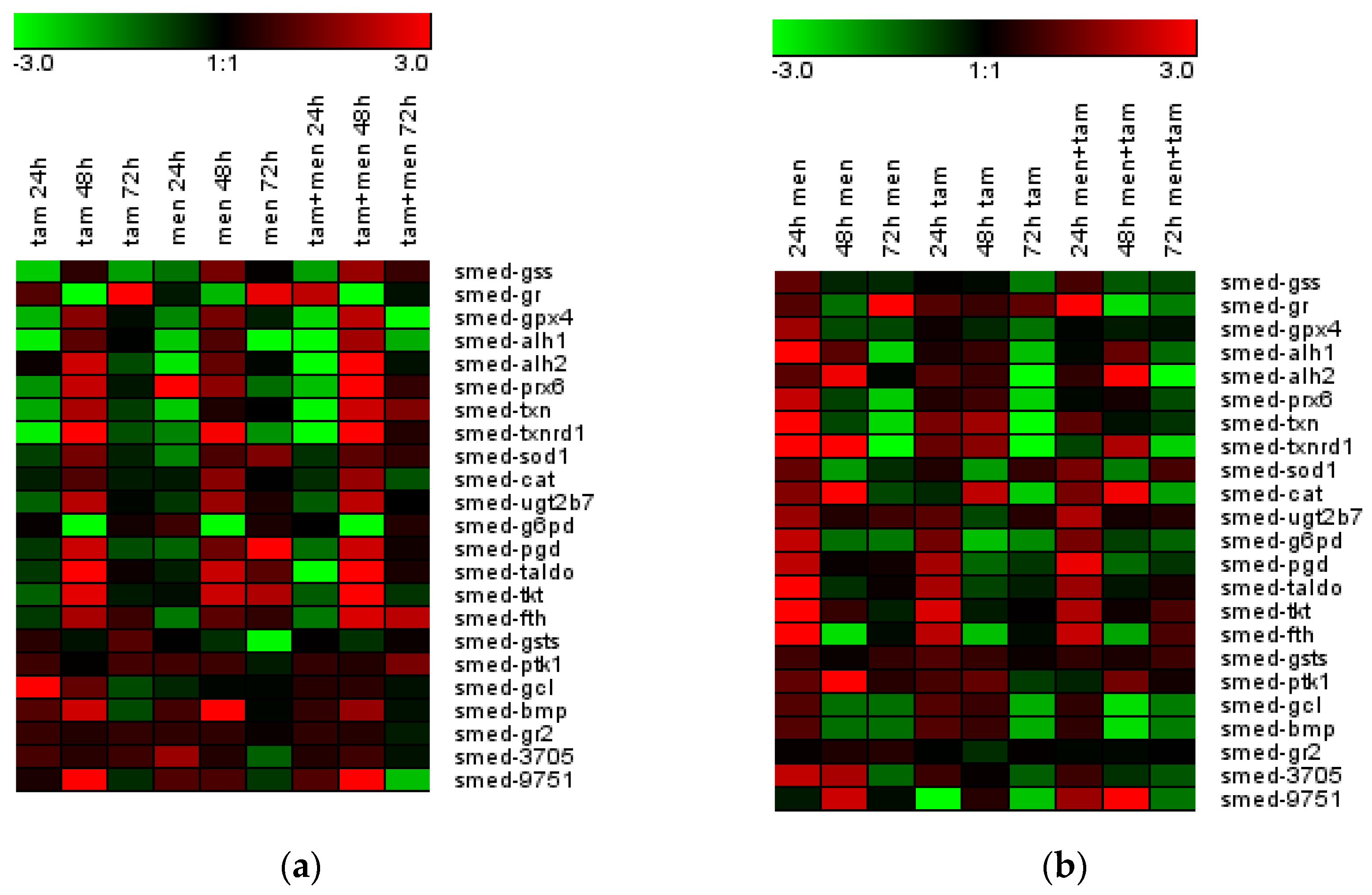
3.4. Tameron Modulates the Expression of Oxidative Stress Response Genes
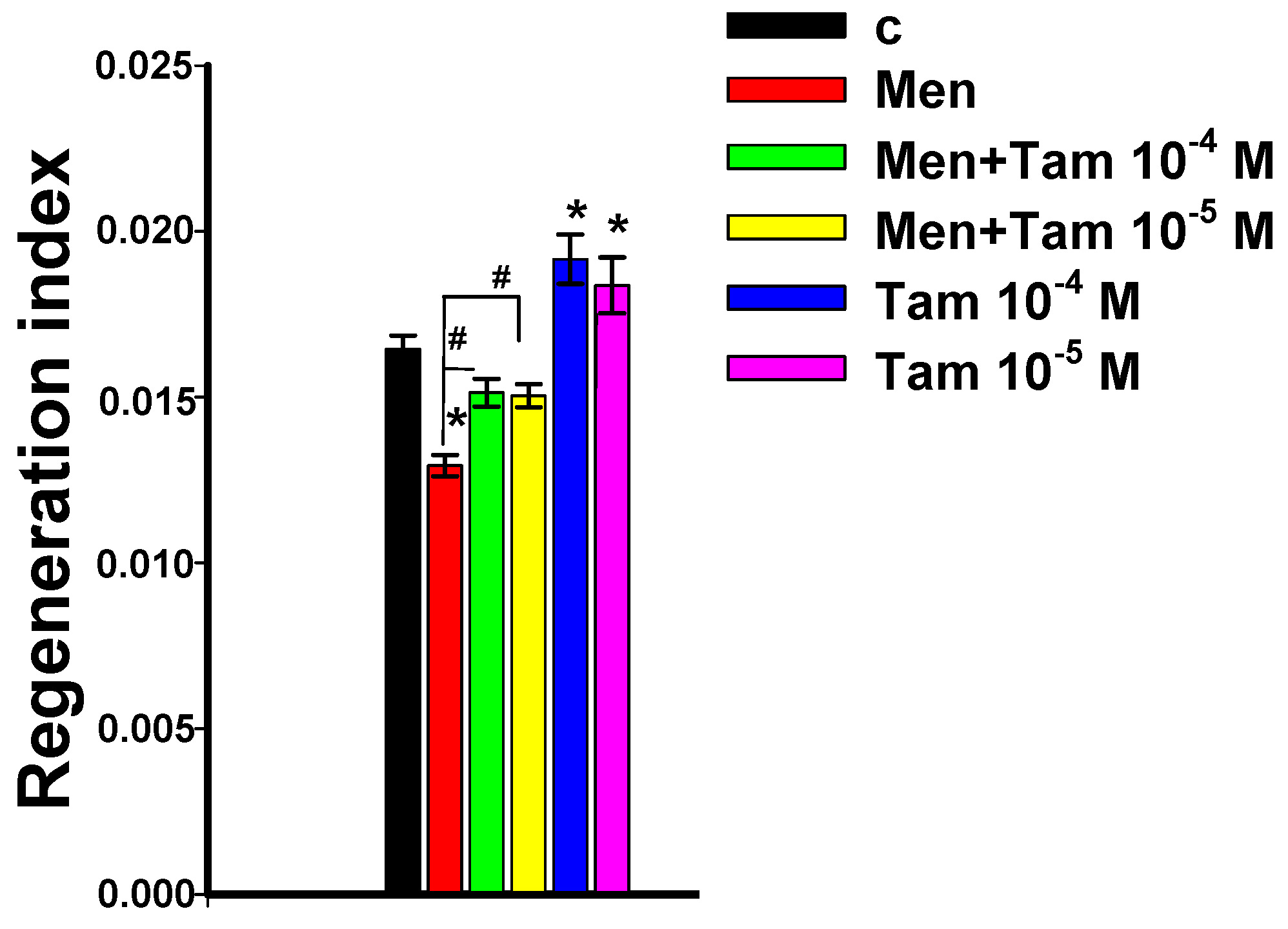
3.5. Tameron Boosts Planarian Regeneration after Menadione-Mediated Inhibition
3.6. Tameron Modulates the Expression of Neoblast Marker Genes after Menadione-Induced Oxidative Stress in Intact and Regenerating Planarians
3.7. Tameron Modulates the Expression of Oxidative Stress Response Genes after Menadione-Induced Oxidative Stress in Intact and Regenerating Planarians
3.8. Tameron Is an Effective ROS Scavenger in the CAA Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sindhi, V.; Gupta, V.; Sharma, K.; Bhatnagar, S.; Kumari, R.; Dhaka, N. Potential applications of antioxidants—A review. J. Pharm. Res. 2013, 7, 828–835. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.Z.; Ranjan, A.; Kaur, N.; Sur, S.; Tandon, V. Radioprotective Agents: Strategies and Translational Advances. Med. Res. Rev. 2016, 36, 461–493. [Google Scholar] [CrossRef] [PubMed]
- Azzam, E.I.; Jay-Gerin, J.P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef]
- Reisz, J.A.; Bansal, N.; Qian, J.; Zhao, W.; Furdui, C.M. Effects of ionizing radiation on biological molecules--mechanisms of damage and emerging methods of detection. Antioxid. Redox Signal. 2014, 21, 260–292. [Google Scholar] [CrossRef]
- Kouvaris, J.R.; Kouloulias, V.E.; Vlahos, L.J. Amifostine: The first selective-target and broad-spectrum radioprotector. Oncologist 2007, 12, 738–747. [Google Scholar] [CrossRef]
- Singh, V.K.; Seed, T.M. The efficacy and safety of amifostine for the acute radiation syndrome. Expert Opin. Drug Saf. 2019, 18, 1077–1090. [Google Scholar] [CrossRef]
- Du, J.; Zhang, P.; Cheng, Y.; Liu, R.; Liu, H.; Gao, F.; Shi, C.; Liu, C. General principles of developing novel radioprotective agents for nuclear emergency. Radiat. Med. Prot. 2020, 1, 120–126. [Google Scholar] [CrossRef]
- Rosen, E.M.; Day, R.; Singh, V.K. New approaches to radiation protection. Front. Oncol. 2014, 4, 381. [Google Scholar] [CrossRef]
- Labarrere, C.A.; Kassab, G.S. Glutathione: A Samsonian life-sustaining small molecule that protects against oxidative stress, ageing and damaging inflammation. Front. Nutr. 2022, 9, 1007816. [Google Scholar] [CrossRef]
- Xue, H.; Li, J.; Xie, H.; Wang, Y. Review of Drug Repositioning Approaches and Resources. Int. J. Biol. Sci. 2018, 14, 1232–1244. [Google Scholar] [CrossRef] [PubMed]
- Shetty, A.K.; Attaluri, S.; Kodali, M.; Shuai, B.; Shetty, G.A.; Upadhya, D.; Hattiangady, B.; Madhu, L.N.; Upadhya, R.; Bates, A.; et al. Monosodium luminol reinstates redox homeostasis, improves cognition, mood and neurogenesis, and alleviates neuro- and systemic inflammation in a model of Gulf War Illness. Redox Biol. 2020, 28, 101389. [Google Scholar] [CrossRef] [PubMed]
- Scofield, V.L.; Yan, M.; Kuang, X.; Kim, S.J.; Crunk, D.; Wong, P.K. The drug monosodium luminol (GVT) preserves thymic epithelial cell cytoarchitecture and allows thymocyte survival in mice infected with the T cell-tropic, cytopathic retrovirus ts1. Immunol. Lett. 2009, 122, 159–169. [Google Scholar] [CrossRef]
- Jiang, Y.; Scofield, V.L.; Yan, M.; Qiang, W.; Liu, N.; Reid, A.J.; Lynn, W.S.; Wong, P.K. Retrovirus-induced oxidative stress with neuroimmunodegeneration is suppressed by antioxidant treatment with a refined monosodium alpha-luminol (Galavit). J. Virol. 2006, 80, 4557–4569. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.V.; Lungu, G.; Kuang, X.; Stoica, G.; Wong, P.K. Neuroprotective effects of the drug GVT (monosodium luminol) are mediated by the stabilization of Nrf2 in astrocytes. Neurochem. Int. 2010, 56, 780–788. [Google Scholar] [CrossRef]
- Schumann, S.; Kaiser, A.; Nicoletti, F. Immune-Modulating Drug MP1032 with SARS-CoV-2 Antiviral Activity In Vitro: A potential Multi-Target Approach for Prevention and Early Intervention Treatment of COVID-19. Int. J. Mol. Sci. 2020, 21, 8803. [Google Scholar] [CrossRef]
- Rink, J.C. Stem cell systems and regeneration in planaria. Dev. Genes Evol. 2013, 223, 67–84. [Google Scholar] [CrossRef]
- Reddien, P.W.; Sánchez Alvarado, A. Fundamentals of planarian regeneration. Annu. Rev. Cell Dev. Biol. 2004, 20, 725–757. [Google Scholar] [CrossRef]
- Roberts-Galbraith, R.H.; Newmark, P.A. On the organ trail: Insights into organ regeneration in the planarian. Curr. Opin. Genet. Dev. 2015, 32, 37–46. [Google Scholar] [CrossRef]
- Pagán, O.R. Planaria: An animal model that integrates development, regeneration and pharmacology. Int. J. Dev. Biol. 2017, 61, 519–529. [Google Scholar] [CrossRef]
- Pellettieri, J.; Sánchez Alvarado, A. Cell turnover and adult tissue homeostasis: From humans to planarians. Annu. Rev. Genet. 2007, 41, 83–105. [Google Scholar] [CrossRef] [PubMed]
- Salvetti, A.; Rossi, L.; Bonuccelli, L.; Lena, A.; Pugliesi, C.; Rainaldi, G.; Evangelista, M.; Gremigni, V. Adult stem cell plasticity: Neoblast repopulation in non-lethally irradiated planarians. Dev. Biol. 2009, 328, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Reddien, P.W.; Oviedo, N.J.; Jennings, J.R.; Jenkin, J.C.; Sánchez Alvarado, A. SMEDWI-2 is a PIWI-like protein that regulates planarian stem cells. Science 2005, 310, 1327–1330. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L.; Cassella, L.; Iacopetti, P.; Ghezzani, C.; Tana, L.; Gimenez, G.; Ghigo, E.; Salvetti, A. Insight into stem cell regulation from sub-lethally irradiated worms. Gene 2018, 662, 37–45. [Google Scholar] [CrossRef]
- Ermakov, A.M.; Kamenskikh, K.A.; Ermakova, O.N.; Blagodatsky, A.S.; Popov, A.L.; Ivanov, V.K. Planarians as an In Vivo Experimental Model for the Study of New Radioprotective Substances. Antioxidants 2021, 10, 1763. [Google Scholar] [CrossRef]
- Goupil, L.S.; Ivry, S.L.; Hsieh, I.; Suzuki, B.M.; Craik, C.S.; O’Donoghue, A.J.; McKerrow, J.H. Cysteine and Aspartyl Proteases Contribute to Protein Digestion in the Gut of Freshwater Planaria. PLoS Negl. Trop. Dis. 2016, 10, e0004893. [Google Scholar] [CrossRef]
- Ermakov, A.M.; Ermakova, O.N.; Maevskiĭ, E.I. A role of some intracellular signaling cascades in planarian regeneration activated under irradiation with low-temperature argon plasma. Biofizika 2014, 59, 552–557. [Google Scholar] [CrossRef]
- Newmark, P.A.; Sánchez Alvarado, A. Bromodeoxyuridine specifically labels the regenerative stem cells of planarians. Dev. Biol. 2000, 220, 142–153. [Google Scholar] [CrossRef]
- Ermakova, O.N.; Ermakov, A.M.; Tiras Kh, P.; Lednev, V.V. Melatonin effect on the regeneration of the flatworm Girardia tigrina. Ontogenez 2009, 40, 466–469. [Google Scholar] [CrossRef]
- Ermakov, A.; Popov, A.; Ermakova, O.; Ivanova, O.; Baranchikov, A.; Kamenskikh, K.; Shekunova, T.; Shcherbakov, A.; Popova, N.; Ivanov, V. The first inorganic mitogens: Cerium oxide and cerium fluoride nanoparticles stimulate planarian regeneration via neoblastic activation. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 104, 109924. [Google Scholar] [CrossRef]
- Ermakov, A.M.; Ermakova, O.N.; Afanasyeva, V.A.; Popov, A.L. Dose-Dependent Effects of Cold Atmospheric Argon Plasma on the Mesenchymal Stem and Osteosarcoma Cells In Vitro. Int. J. Mol. Sci. 2021, 22, 6797. [Google Scholar] [CrossRef]
- van Wolfswinkel, J.C.; Wagner, D.E.; Reddien, P.W. Single-cell analysis reveals functionally distinct classes within the planarian stem cell compartment. Cell Stem Cell 2014, 15, 326–339. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Yamamoto, M. Molecular mechanisms activating the Nrf2-Keap1 pathway of antioxidant gene regulation. Antioxid. Redox Signal. 2005, 7, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Sturn, A.; Quackenbush, J.; Trajanoski, Z. Genesis: Cluster analysis of microarray data. Bioinformatics 2002, 18, 207–208. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.G.; Noh, W.J.; Kim, H.; Lee, M.Y. Generation of reactive oxygen species contributes to the development of carbon black cytotoxicity to vascular cells. Toxicol. Res. 2011, 27, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Guedelhoefer, O.C.T.; Sánchez Alvarado, A. Amputation induces stem cell mobilization to sites of injury during planarian regeneration. Development 2012, 139, 3510–3520. [Google Scholar] [CrossRef] [PubMed]
- Hassan, G.S. Menadione. Profiles Drug Subst. Excip. Relat. Methodol. 2013, 38, 227–313. [Google Scholar] [CrossRef]
- Abe, K.; Saito, H. Menadione toxicity in cultured rat cortical astrocytes. Jpn. J. Pharmacol. 1996, 72, 299–306. [Google Scholar] [CrossRef]
- Cifra, M.; Pospíšil, P. Ultra-weak photon emission from biological samples: Definition, mechanisms, properties, detection and applications. J. Photochem. Photobiol. B Biol. 2014, 139, 2–10. [Google Scholar] [CrossRef]
- Liu, Z.; Deng, P.; Liu, S.; Bian, Y. Is Nuclear Factor Erythroid 2-Related Factor 2 a Target for the Intervention of Cytokine Storms? Antioxidants 2023, 12, 172. [Google Scholar] [CrossRef]
- Qiang, W.; Cahill, J.M.; Liu, J.; Kuang, X.; Liu, N.; Scofield, V.L.; Voorhees, J.R.; Reid, A.J.; Yan, M.; Lynn, W.S.; et al. Activation of transcription factor Nrf-2 and its downstream targets in response to moloney murine leukemia virus ts1-induced thiol depletion and oxidative stress in astrocytes. J. Virol. 2004, 78, 11926–11938. [Google Scholar] [CrossRef] [PubMed]
- Qiang, W.; Kuang, X.; Liu, J.; Liu, N.; Scofield, V.L.; Reid, A.J.; Jiang, Y.; Stoica, G.; Lynn, W.S.; Wong, P.K. Astrocytes survive chronic infection and cytopathic effects of the ts1 mutant of the retrovirus Moloney murine leukemia virus by upregulation of antioxidant defenses. J. Virol. 2006, 80, 3273–3284. [Google Scholar] [CrossRef] [PubMed]
- Warabi, E.; Takabe, W.; Minami, T.; Inoue, K.; Itoh, K.; Yamamoto, M.; Ishii, T.; Kodama, T.; Noguchi, N. Shear stress stabilizes NF-E2-related factor 2 and induces antioxidant genes in endothelial cells: Role of reactive oxygen/nitrogen species. Free Radic. Biol. Med. 2007, 42, 260–269. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Chen, M.G.; Ma, Q. Activation of Nrf2 in defense against cadmium-induced oxidative stress. Chem. Res. Toxicol. 2008, 21, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Calkins, M.J.; Chan, K.; Kan, Y.W.; Johnson, J.A. Identification of the NF-E2-related factor-2-dependent genes conferring protection against oxidative stress in primary cortical astrocytes using oligonucleotide microarray analysis. J. Biol. Chem. 2003, 278, 12029–12038. [Google Scholar] [CrossRef] [PubMed]
- Deger, Y.; Dede, S.; Belge, A.; Mert, N.; Kahraman, T.; Alkan, M. Effects of X-ray radiation on lipid peroxidation and antioxidant systems in rabbits treated with antioxidant compounds. Biol. Trace Elem. Res. 2003, 94, 149–156. [Google Scholar] [CrossRef]
- Pirotte, N.; Stevens, A.-S.; Fraguas, S.; Plusquin, M.; Van Roten, A.; Van Belleghem, F.; Paesen, R.; Ameloot, M.; Cebrià, F.; Artois, T.; et al. Reactive Oxygen Species in Planarian Regeneration: An Upstream Necessity for Correct Patterning and Brain Formation. Oxidative Med. Cell. Longev. 2015, 2015, 392476. [Google Scholar] [CrossRef]
- Dikalov, S.I.; Harrison, D.G. Methods for detection of mitochondrial and cellular reactive oxygen species. Antioxid. Redox Signal. 2014, 20, 372–382. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsarkova, E.; Filippova, K.; Afanasyeva, V.; Ermakova, O.; Kolotova, A.; Blagodatski, A.; Ermakov, A. A Study on the Planarian Model Confirms the Antioxidant Properties of Tameron against X-ray- and Menadione-Induced Oxidative Stress. Antioxidants 2023, 12, 953. https://doi.org/10.3390/antiox12040953
Tsarkova E, Filippova K, Afanasyeva V, Ermakova O, Kolotova A, Blagodatski A, Ermakov A. A Study on the Planarian Model Confirms the Antioxidant Properties of Tameron against X-ray- and Menadione-Induced Oxidative Stress. Antioxidants. 2023; 12(4):953. https://doi.org/10.3390/antiox12040953
Chicago/Turabian StyleTsarkova, Elena, Kristina Filippova, Vera Afanasyeva, Olga Ermakova, Anastasia Kolotova, Artem Blagodatski, and Artem Ermakov. 2023. "A Study on the Planarian Model Confirms the Antioxidant Properties of Tameron against X-ray- and Menadione-Induced Oxidative Stress" Antioxidants 12, no. 4: 953. https://doi.org/10.3390/antiox12040953
APA StyleTsarkova, E., Filippova, K., Afanasyeva, V., Ermakova, O., Kolotova, A., Blagodatski, A., & Ermakov, A. (2023). A Study on the Planarian Model Confirms the Antioxidant Properties of Tameron against X-ray- and Menadione-Induced Oxidative Stress. Antioxidants, 12(4), 953. https://doi.org/10.3390/antiox12040953