

Stevioside Improves Antioxidant Capacity and Intestinal Barrier Function while Attenuating Inflammation and Apoptosis by Regulating the NF-κB/MAPK Pathways in Diquat-Induced Oxidative Stress of IPEC-J2 Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture and Treatments
2.3. Cell Viability Assay
2.4. Measurement of Cell Proliferation
2.5. Flow Cytometric Determination of Cell Apoptosis
2.6. Measurement of Intracellular ROS Production
2.7. Determination of Oxidative Stress Parameters
2.8. Detection of Cytokines
2.9. Quantitative Reverse-Transcription Polymerase Chain Reaction (qRT-PCR)
2.10. Western Blotting
2.11. Statistical Analysis
3. Results
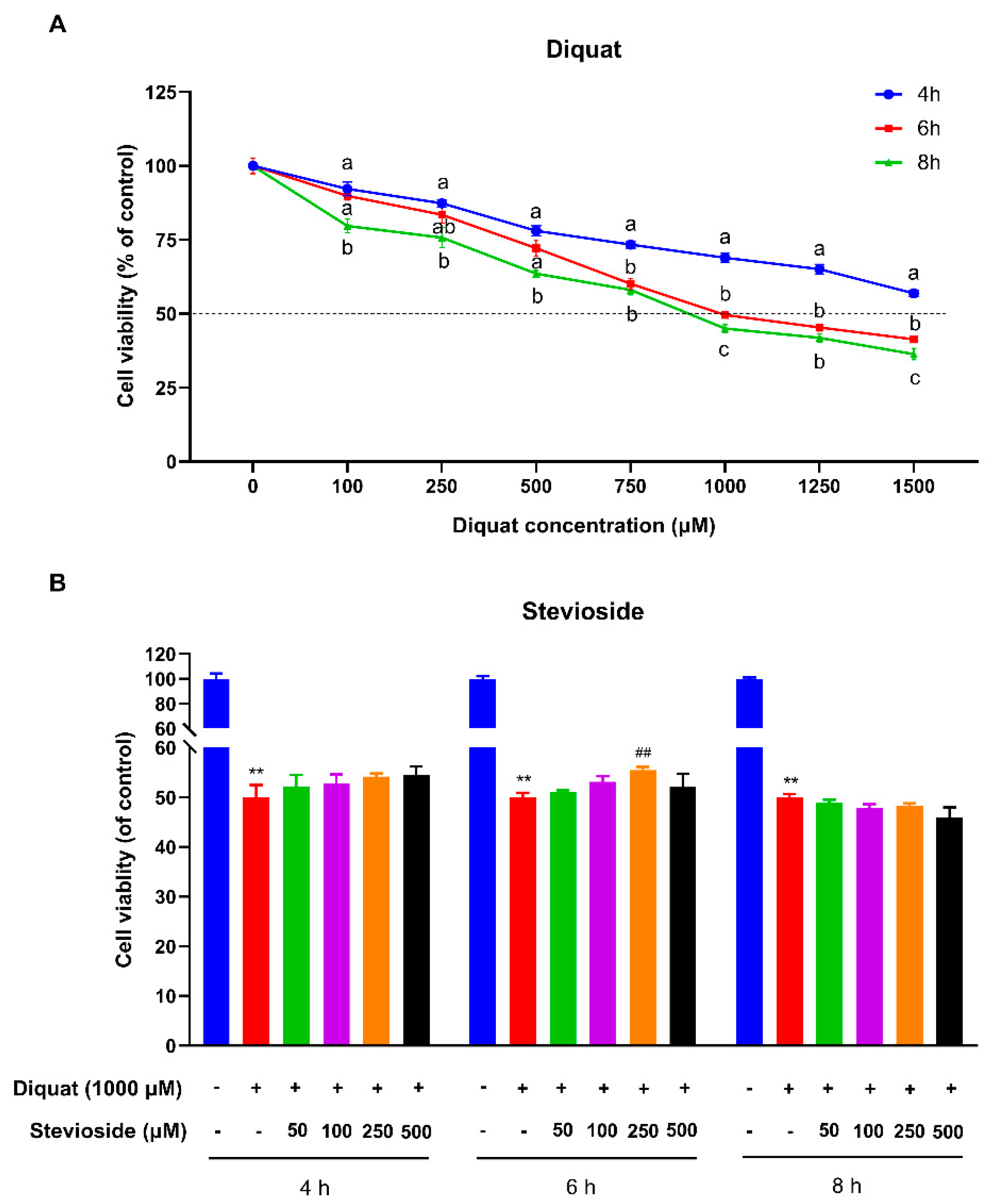
3.1. Stevioside Attenuated Diquat-Induced Cytotoxicity in IPEC-J2 Cells
3.2. Stevioside Promoted Cell Proliferation in Diquat-Treated IPEC-J2 Cells
3.3. Stevioside Alleviated Diquat-Induced Apoptosis in IPEC-J2 Cells
3.4. Stevioside Regulated Barrier Function in Diquat-Induced IPEC-J2 Cells
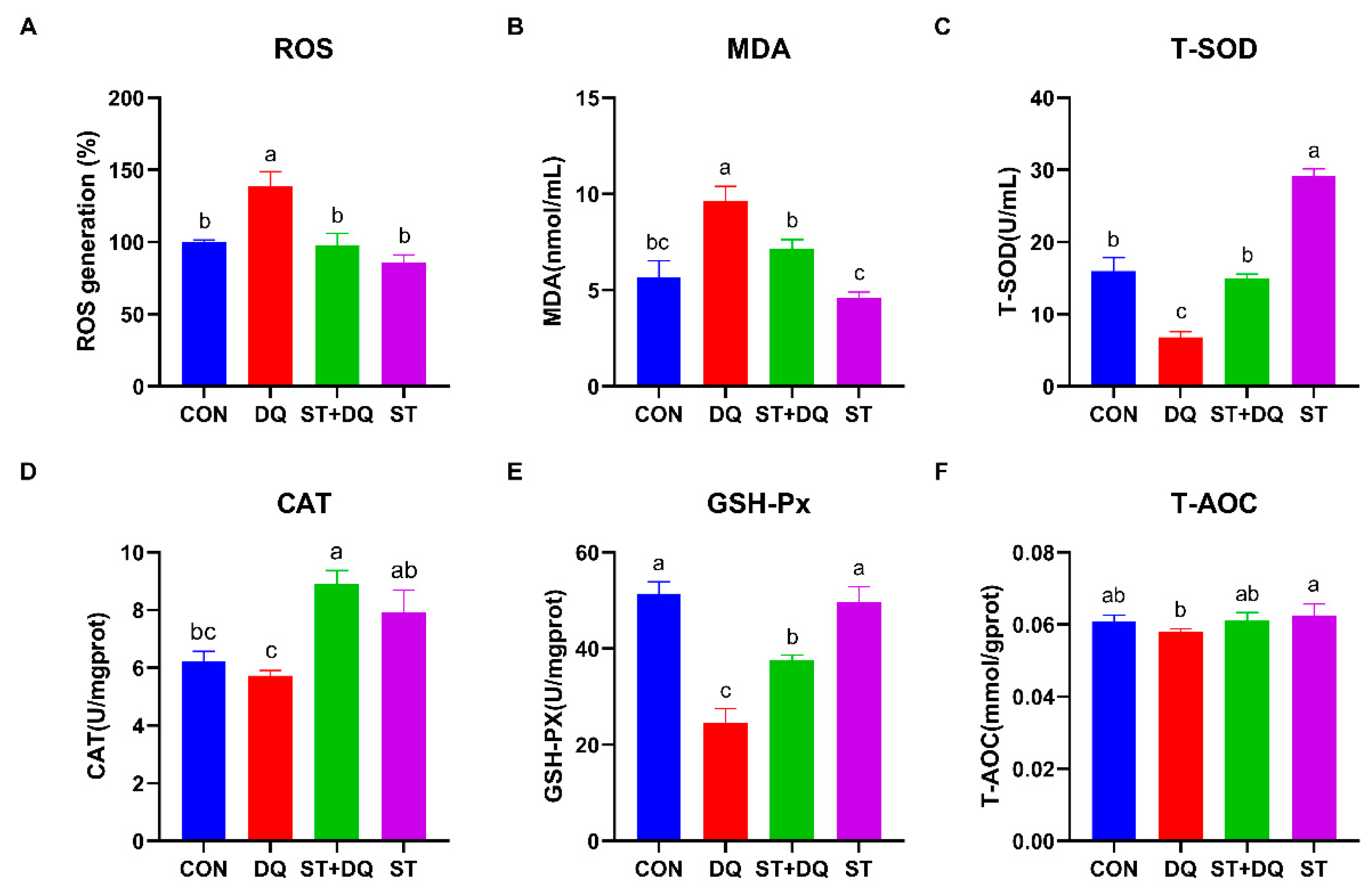
3.5. Stevioside Regulated the Cellular Redox State in Diquat-Induced IPEC-J2 Cells
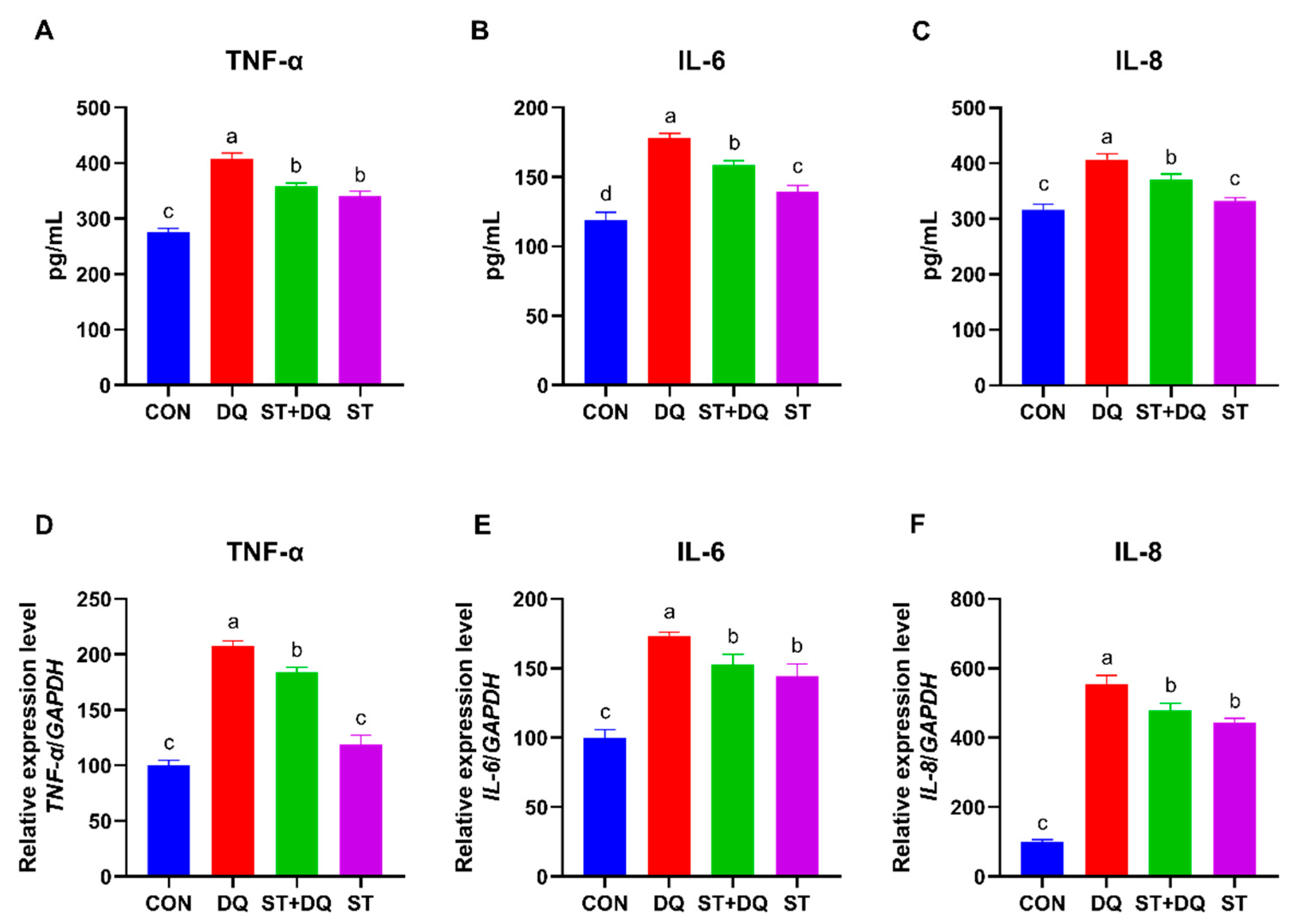
3.6. Effects of Stevioside on Inflammatory Cytokines
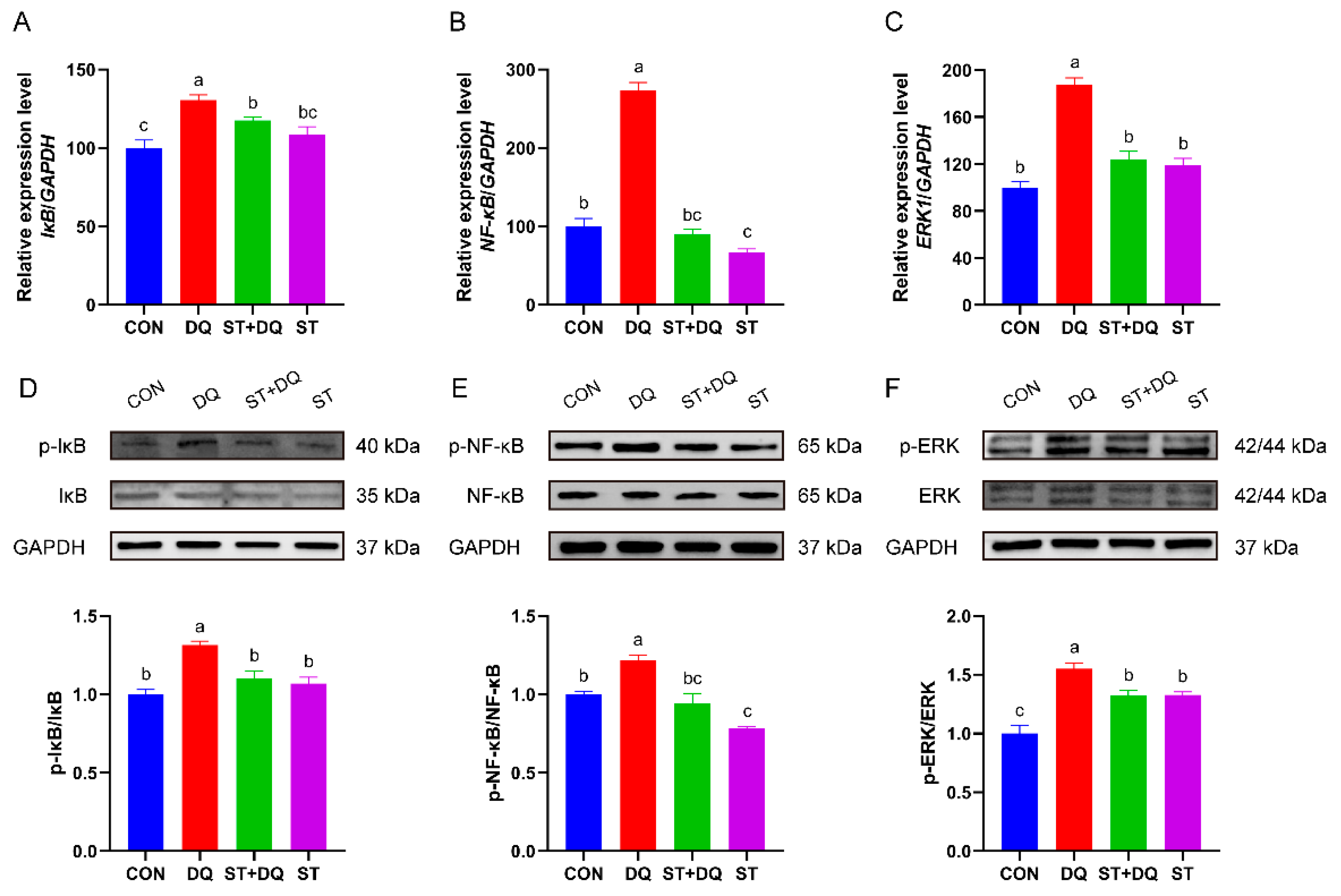
3.7. Stevioside Regulated NF-κB/MAPK Signaling Pathways in IPEC-J2 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, W.; Chen, G.; Yan, J.; Wang, X.; Zhu, Y.; Zhu, L. Single-cell sequencing analysis reveals gastric cancer microenvironment cells respond vastly different to oxidative stress. J. Transl. Med. 2022, 20, 250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Zhao, W.Y.; Li, B.; Li, W.Q.; Zhang, C.X.; Hou, X.C.; Jiang, J.; Dong, Y.Z. Ratiometric fluorescent probes for capturing endogenous hypochlorous acid in the lungs of mice. Chem. Sci. 2018, 9, 8207–8212. [Google Scholar] [CrossRef] [PubMed]
- Lauridsen, C. Effects of dietary fatty acids on gut health and function of pigs pre- and post-weaning. J. Anim. Sci. 2020, 98, skaa086. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.J.; Wang, L.X.; Yang, H.S.; Hu, A.; Yin, Y.L. Review: The roles and functions of glutamine on intestinal health and performance of weaning pigs. Animal 2019, 13, 2727–2735. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Xu, B.; Chen, Y.; Yang, W.; Xu, Y.; Huang, J.; Duo, T.; Mao, Y.; Zhou, G.; Yan, X.; et al. Dietary ellagic acid supplementation attenuates intestinal damage and oxidative stress by regulating gut microbiota in weanling piglets. Anim. Nutr. 2022, 11, 322–333. [Google Scholar] [CrossRef]
- Wijtten, P.J.A.; van der Meulen, J.; Verstegen, M.W.A. Intestinal barrier function and absorption in pigs after weaning: A review. Brit. J. Nutr. 2011, 105, 967–981. [Google Scholar] [CrossRef]
- Kang, H.J.; Lee, H.N.; Hong, S.J.; Park, B.R.; Ameer, K.; Cho, J.Y.; Kim, Y.M. Synthesis and characteristics of a rebaudioside-A like compound as a potential non-caloric natural sweetener by Leuconostoc kimchii dextransucrase. Food Chem. 2022, 366, 130623. [Google Scholar] [CrossRef]
- Chatsudthipong, V.; Muanprasat, C. Stevioside and related compounds: Therapeutic benefits beyond sweetness. Pharmacol. Therapeut. 2009, 121, 41–54. [Google Scholar] [CrossRef]
- Ragone, M.I.; Bonazzola, P.; Colareda, G.A.; Lazarte, M.L.; Bruno, F.; Consolini, A.E. Cardioprotection of stevioside on stunned rat hearts: A mechano-energetical study. Phytomedicine 2017, 35, 18–26. [Google Scholar] [CrossRef]
- Casas-Grajales, S.; Ramos-Tovar, E.; Chavez-Estrada, E.; Alvarez-Suarez, D.; Hernandez-Aquino, E.; Reyes-Gordillo, K.; Cerda-Garcia-Roja, C.M.; Camacho, J.; Tsutsumi, V.; Lakshman, M.R.; et al. Antioxidant and immunomodulatory activity induced by stevioside in liver damage: In vivo, in vitro and in silico assays. Life Sci. 2019, 224, 187–196. [Google Scholar] [CrossRef]
- Wei, F.Y.; Zhu, H.; Li, N.; Yu, C.L.; Song, Z.B.; Wang, S.Y.; Sun, Y.; Zheng, L.H.; Wang, G.N.; Huang, Y.X.; et al. Stevioside Activates AMPK to Suppress Inflammation in Macrophages and Protects Mice from LPS-Induced Lethal Shock. Molecules 2021, 26, 858. [Google Scholar] [CrossRef] [PubMed]
- Sehar, I.; Kaul, A.; Bani, S.; Pal, H.C.; Saxena, A.K. Immune up regulatory response of a non-caloric natural sweetener, stevioside. Chem. Biol. Interact. 2008, 173, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.; Ma, X.; Liu, Y.L.; Qiao, S.Y.; Hou, Y.Q.; Zhang, G.L. Dietary modulation of endogenous host defense peptide synthesis as an alternative approach to in-feed antibiotics. Anim. Nutr. 2018, 4, 160–169. [Google Scholar] [CrossRef]
- Han, X.F.; Chen, C.X.; Zhang, X.L.; Wei, Y.Q.; Tang, S.X.; Wang, J.R.; Tan, Z.L.; Xu, L.W. Effects of Dietary Stevioside Supplementation on Feed Intake, Digestion, Ruminal Fermentation, and Blood Metabolites of Goats. Animals 2019, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Qi, L.; Wei, Q.; Shi, F. Maternal stevioside supplementation improves intestinal immune function of chicken offspring potentially via modulating gut microbiota and down-regulating the promoter methylation level of suppressor of cytokine signaling 1 (SOCS1). Anim. Nutr. 2022, 10, 329–346. [Google Scholar] [CrossRef]
- Jiang, J.L.; Qi, L.N.; Lv, Z.P.; Jin, S.; Wei, X.H.; Shi, F.X. Dietary Stevioside Supplementation Alleviates Lipopolysaccharide-Induced Intestinal Mucosal Damage through Anti-Inflammatory and Antioxidant Effects in Broiler Chickens. Antioxidants 2019, 8, 575. [Google Scholar] [CrossRef]
- Shin, Y.G.; Rathnayake, D.; Mun, H.S.; Dilawar, M.A.; Pov, S.; Yang, C.J. Sensory Attributes, Microbial Activity, Fatty Acid Composition and Meat Quality Traits of Hanwoo Cattle Fed a Diet Supplemented with Stevioside and Organic Selenium. Foods 2021, 10, 129. [Google Scholar] [CrossRef]
- Jiang, J.; Liu, S.; Jamal, T.; Ding, T.; Qi, L.; Lv, Z.; Yu, D.; Shi, F. Effects of dietary sweeteners supplementation on growth performance, serum biochemicals, and jejunal physiological functions of broiler chickens. Poult. Sci. 2020, 99, 3948–3958. [Google Scholar] [CrossRef]
- Xu, Q.L.; Liu, C.; Mo, X.J.; Chen, M.; Zhao, X.L.; Liu, M.Z.; Wang, S.B.; Zhou, B.; Zhao, C.X. Drinking Water Supplemented with Acidifiers Improves the Growth Performance of Weaned Pigs and Potentially Regulates Antioxidant Capacity, Immunity, and Gastrointestinal Microbiota Diversity. Antioxidants 2022, 11, 809. [Google Scholar] [CrossRef]
- Chen, J.L.; Chen, D.W.; Yu, B.; Luo, Y.H.; Zheng, P.; Mao, X.B.; Yu, J.; Luo, J.Q.; Huang, Z.Q.; Yan, H.; et al. Chlorogenic Acid Attenuates Oxidative Stress-Induced Intestinal Mucosa Disruption in Weaned Pigs. Front. Vet. Sci. 2022, 9, 806253. [Google Scholar] [CrossRef]
- Belle, N.M.; Ji, Y.B.; Herbine, K.; Wei, Y.; Park, J.; Zullo, K.; Hung, L.Y.; Srivatsa, S.; Young, T.; Oniskey, T.; et al. TFF3 interacts with LINGO2 to regulate EGFR activation for protection against colitis and gastrointestinal helminths. Nat. Commun. 2019, 10, 4408. [Google Scholar] [CrossRef] [PubMed]
- Brosnahan, A.J.; Brown, D.R. Porcine IPEC-J2 intestinal epithelial cells in microbiological investigations. Vet. Microbiol. 2012, 156, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Huang, S.B.; Wu, Z.F.; Huang, Z.H.; Hao, X.Y.; Zhang, L.M.; Hu, C.J.; Wei, J.F.; Deng, J.P.; Tan, C.Q. Maternal supply of cysteamine alleviates oxidative stress and enhances angiogenesis in porcine placenta. J. Anim. Sci. Biotechnol. 2021, 12, 91. [Google Scholar] [CrossRef]
- Novais, A.K.; Martel-Kennes, Y.; Roy, C.; Deschene, K.; Beaulieu, S.; Bergeron, N.; Laforest, J.P.; Lessard, M.; Matte, J.J.; Lapointe, J. Tissue-specific profiling reveals modulation of cellular and mitochondrial oxidative stress in normal- and low-birthweight piglets throughout the peri-weaning period. Animal 2020, 14, 1014–1024. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.D.; Bai, Y.S.; Li, J.B.; Ren, Z.S.; Li, J.P.; Zhang, J.; Shan, A.S. Lactobacillus rhamnosus GG ameliorates deoxynivalenol-induced kidney oxidative damage and mitochondrial injury in weaned piglets. Food Funct. 2022, 13, 3905–3916. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Huang, K.; Liu, J.Z.; Wu, S.; Shen, D.; Dai, P.Y.; Li, C.M. Fine particulate matter from pig house induced immune response by activating TLR4/MAPK/NF-kappa B pathway and NLRP3 inflammasome in alveolar macrophages. Chemosphere 2019, 236, 124373. [Google Scholar] [CrossRef]
- He, Y.L.; Sang, Z.; Zhuo, Y.S.; Wang, X.Y.; Guo, Z.H.; He, L.H.; Zeng, C.P.; Dai, H.C. Transport stress induces pig jejunum tissue oxidative damage and results in autophagy/mitophagy activation. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1521–1529. [Google Scholar] [CrossRef]
- Frame, C.A.; Johnson, E.; Kilburn, L.; Huff-Lonergan, E.; Kerr, B.J.; Serao, M.R. Impact of dietary oxidized protein on oxidative status and performance in growing pigs. J. Anim. Sci. 2020, 98, skaa097. [Google Scholar] [CrossRef]
- Suez, J.; Cohen, Y.; Valdes-Mas, R.; Mor, U.; Dori-Bachash, M.; Federici, S.; Zmora, N.; Leshem, A.; Heinemann, M.; Linevsky, R.; et al. Personalized microbiome-driven effects of non-nutritive sweeteners on human glucose tolerance. Cell 2022, 185, 3307–3328.e19. [Google Scholar] [CrossRef]
- Alavala, S.; Sangaraju, R.; Nalban, N.; Sahu, B.D.; Jerald, M.K.; Kilari, E.K.; Sistla, R. Stevioside, a diterpenoid glycoside, shows anti-inflammatory property against Dextran Sulphate Sodium-induced ulcerative colitis in mice. Eur. J. Pharmacol. 2019, 855, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Zhong, K.L.; Lu, M.Y.; Liu, F.; Mei, Y.; Zhang, X.J.; Zhang, H.; Zan, J.; Sun, X.O.; Tan, W. Isosteviol Sodium Protects Neural Cells Against Hypoxia-Induced Apoptosis Through Inhibiting MAPK and NF-kappa B Pathways. J. Stroke Cerebrovasc. Dis. 2019, 28, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Velesiotis, C.; Kanellakis, M.; Vynios, D.H. Steviol glycosides affect functional properties and macromolecular expression of breast cancer cells. IUBMB Life 2022, 74, 1012–1028. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.P.; Liu, B.; Wang, X.R.; Yu, Q.F.; Fang, R.J. Epidermal Growth Factor, through Alleviating Oxidative Stress, Protect IPEC-J2 Cells from Lipopolysaccharides-Induced Apoptosis. Int. J. Mol. Sci. 2018, 19, 848. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.W.; Shin, Y.J.; Luo, K.; Quan, Y.; Cui, S.; Ko, E.J.; Chung, B.H.; Yang, C.W. Ginseng increases Klotho expression by FoxO3-mediated manganese superoxide dismutase in a mouse model of tacrolimus-induced renal injury. Aging 2019, 11, 5548–5569. [Google Scholar] [CrossRef]
- Jin, Y.; Zhai, Z.; Jia, H.; Lai, J.; Si, X.; Wu, Z. Kaempferol attenuates diquat-induced oxidative damage and apoptosis in intestinal porcine epithelial cells. Food Funct. 2021, 12, 6889–6899. [Google Scholar] [CrossRef]
- Detterich, J.A.; Liu, H.L.; Suriany, S.; Kato, R.M.; Chalacheva, P.; Tedla, B.; Shah, P.M.; Khoo, M.C.; Wood, J.C.; Coates, T.D.; et al. Erythrocyte and plasma oxidative stress appears to be compensated in patients with sickle cell disease during a period of relative health, despite the presence of known oxidative agents. Free Radic. Biol. Med. 2019, 141, 408–415. [Google Scholar] [CrossRef]
- Vergauwen, H.; Prims, S.; Degroote, J.; Wang, W.; Casteleyn, C.; van Cruchten, S.; de Smet, S.; Michiels, J.; van Ginneken, C. In Vitro Investigation of Six Antioxidants for Pig Diets. Antioxidants 2016, 5, 41. [Google Scholar] [CrossRef]
- Gori, M.; Altomare, A.; Cocca, S.; Solida, E.; Ribolsi, M.; Carotti, S.; Rainer, A.; Francesconi, M.; Morini, S.; Cicala, M.; et al. Palmitic Acid Affects Intestinal Epithelial Barrier Integrity and Permeability In Vitro. Antioxidants 2020, 9, 417. [Google Scholar] [CrossRef]
- Kim, M.; Chung, K.S.; Hwang, S.J.; Yoon, Y.S.; Jang, Y.P.; Lee, J.K.; Lee, K.T. Protective Effect of Cicer arietinum L. (Chickpea) Ethanol Extract in the Dextran Sulfate Sodium-Induced Mouse Model of Ulcerative Colitis. Nutrients 2020, 12, 456. [Google Scholar] [CrossRef]
- Feng, R.; Ma, L.J.; Wang, M.; Liu, C.; Yang, R.; Su, H.; Yang, Y.; Wan, J.B. Oxidation of fish oil exacerbates alcoholic liver disease by enhancing intestinal dysbiosis in mice. Commun. Biol. 2020, 3, 481. [Google Scholar] [CrossRef]
- Zhang, X.J.; Gu, J.X.; Zhao, C.Y.; Hu, Y.Z.; Zhang, B.W.; Wang, J.; Lv, H.; Ji, X.M.; Wang, S. Sweeteners Maintain Epithelial Barrier Function through the miR-15b/RECK/MMP-9 Axis, Remodel Microbial Homeostasis, and Attenuate Dextran Sodium Sulfate-Induced Colitis in Mice. J. Agric. Food Chem. 2022, 70, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Kurtoglu, Y.E.; Navath, R.S.; Wang, B.; Kannan, S.; Romero, R.; Kannan, R.M. Poly(amidoamine) dendrimer-drug conjugates with disulfide linkages for intracellular drug delivery. Biomaterials 2009, 30, 2112–2121. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Zeng, Z.H.; Zhou, Y.H.; Wang, B.K.; Zou, P.; Wang, Q.; Ying, J.F.; Wang, F.; Li, X.; Xu, S.J.; et al. Bacillus amyloliquefaciens SC06 Induced AKT-FOXO Signaling Pathway-Mediated Autophagy to Alleviate Oxidative Stress in IPEC-J2 Cells. Antioxidants 2021, 10, 1545. [Google Scholar] [CrossRef]
- Lopez, V.; Perez, S.; Vinuesa, A.; Zorzetto, C.; Abian, O. Stevia rebaudiana ethanolic extract exerts better antioxidant properties and antiproliferative effects in tumour cells than its diterpene glycoside stevioside. Food Funct. 2016, 7, 2107–2113. [Google Scholar] [CrossRef]
- Jung, H.I.; Kong, M.S.; Lee, B.R.; Kim, T.H.; Chae, M.J.; Lee, E.J.; Jung, G.B.; Lee, C.H.; Sung, J.K.; Kim, Y.H. Exogenous Glutathione Increases Arsenic Translocation into Shoots and Alleviates Arsenic-Induced Oxidative Stress by Sustaining Ascorbate-Glutathione Homeostasis in Rice Seedlings. Front. Plant Sci. 2019, 10, 1089. [Google Scholar] [CrossRef]
- Sun, K.Y.; Sun, Y.T.; Li, H.Y.; Han, D.Y.; Bai, Y.T.; Zhao, R.; Guo, Z.J. Anti-Ageing Effect of Physalis alkekengi Ethyl Acetate Layer on a d-galactose-Induced Mouse Model through the Reduction of Cellular Senescence and Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 1836. [Google Scholar] [CrossRef]
- Salcher, S.; Hagenbuchner, J.; Geiger, K.; Seiter, M.A.; Rainer, J.; Kofler, R.; Hermann, M.; Kiechl-Kohlendorfer, U.; Ausserlechner, M.J.; Obexer, P. C10ORF10/DEPP, a transcriptional target of FOXO3, regulates ROS-sensitivity in human neuroblastoma. Mol. Cancer 2014, 13, 224. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, J.; Peng, J.; Wei, H.K. Oregano Essential Oil Induces SOD1 and GSH Expression through Nrf2 Activation and Alleviates Hydrogen Peroxide-Induced Oxidative Damage in IPEC-J2 Cells. Oxid. Med. Cell. Longev. 2016, 2016, 5987183. [Google Scholar] [CrossRef] [PubMed]
- Li, F.M.; Han, X.K.; Li, F.; Wang, R.; Wang, H.; Gao, Y.J.; Wang, X.J.; Fang, Z.Y.; Zhang, W.J.; Yao, S.; et al. LKB1 Inactivation Elicits a Redox Imbalance to Modulate Non-small Cell Lung Cancer Plasticity and Therapeutic Response. Cancer Cell 2015, 27, 698–711. [Google Scholar] [CrossRef]
- Liu, S.; Xiong, Y.; Cao, S.; Wen, X.; Xiao, H.; Li, Y.; Chi, L.; He, D.; Jiang, Z.; Wang, L. Dietary Stevia Residue Extract Supplementation Improves Antioxidant Capacity and Intestinal Microbial Composition of Weaned Piglets. Antioxidants 2022, 11, 2016. [Google Scholar] [CrossRef] [PubMed]
- Seyedsadjadi, N.; Berg, J.; Bilgin, A.A.; Grant, R. A Pilot Study Providing Evidence for a Relationship between a Composite Lifestyle Score and Risk of Higher Carotid Intima-Media Thickness: Is There a Link to Oxidative Stress? Oxid. Med. Cell. Longev. 2018, 2018, 4504079. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.X.; Wei, Y.J.; Shah, M.K.; Wang, X.Y.; Lin, J.T.; Wan, P.; Cui, L.; Yin, Q.Q. Effects of Compound Active Peptides on Protecting Liver and Intestinal Epithelial Cells from Damages and Preventing Hyperglycemia. Oxid. Med. Cell. Longev. 2020, 2020, 3183104. [Google Scholar] [CrossRef]
- Boonkaewwan, C.; Burodom, A. Anti-inflammatory and immunomodulatory activities of stevioside and steviol on colonic epithelial cells. J. Sci. Food Agric. 2013, 93, 3820–3825. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yang, P.; Sifa, D.; Wen, Z. Effect of dietary stevioside supplementation on growth performance, nutrient digestibility, serum parameters, and intestinal microflora in broilers. Food Funct. 2019, 10, 2340–2346. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Jiang, L.Y.; Long, X.; Fu, C.P.; Wang, X.D.; Wu, X.D.; Liu, Z.L.; Zhu, F.; Shi, J.D.; Li, S.Q. Bone-marrow-derived mesenchymal stem cells inhibit gastric aspiration lung injury and inflammation in rats. J. Cell. Mol. Med. 2016, 20, 1706–1717. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Al-Alwan, L.; Alshakfa, S.; Audusseau, S.; Mogas, A.K.; Chouiali, F.; Nair, P.; Baglole, C.J.; Hamid, Q.; Eidelman, D.H. Upregulation of IL-17A/F from human lung tissue explants with cigarette smoke exposure: Implications for COPD. Respir. Res. 2014, 15, 145. [Google Scholar] [CrossRef]
- Shi, C.; Zhan, L.; Wu, Y.; Li, Z.; Li, J.; Li, Y.; Wei, J.; Zhang, Y.; Li, L. Kaji-Ichigoside F1 and Rosamultin Protect Vascular Endothelial Cells against Hypoxia-Induced Apoptosis via the PI3K/AKT or ERK1/2 Signaling Pathway. Oxid. Med. Cell. Longev. 2020, 2020, 6837982. [Google Scholar] [CrossRef]
- Fengyang, L.; Yunhe, F.; Bo, L.; Zhicheng, L.; Depeng, L.; Dejie, L.; Wen, Z.; Yongguo, C.; Naisheng, Z.; Xichen, Z.; et al. Stevioside suppressed inflammatory cytokine secretion by downregulation of NF-kappaB and MAPK signaling pathways in LPS-stimulated RAW264.7 cells. Inflammation 2012, 35, 1669–1675. [Google Scholar] [CrossRef]
- Kim, S.M.; Park, E.J.; Lee, H.J. Nuciferine attenuates lipopolysaccharide-stimulated inflammatory responses by inhibiting p38 MAPK/ATF2 signaling pathways. Inflammopharmacology 2022, 30, 2373–2383. [Google Scholar] [CrossRef]
- Wang, C.; Shi, C.; Su, W.; Jin, M.; Xu, B.; Hao, L.; Zhang, Y.; Lu, Z.; Wang, F.; Wang, Y.; et al. Dynamics of the Physicochemical Characteristics, Microbiota, and Metabolic Functions of Soybean Meal and Corn Mixed Substrates during Two-Stage Solid-State Fermentation. mSystems 2020, 5, e00501-19. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Li, X.; An, X.R.; Liu, W.; Yuan, T. Masticadienonic acid from Chios mastic gum mitigates colitis in mice via modulating inflammatory response, gut barrier integrity and microbiota. Phytomedicine 2023, 108, 154518. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Pastor-Villaescusa, B.; Rueda-Robles, A.; Abadia-Molina, F.; Ruiz-Ojeda, F.J. Plausible Biological Interactions of Low- and Non-Calorie Sweeteners with the Intestinal Microbiota: An Update of Recent Studies. Nutrients 2020, 12, 1153. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Q.; Liu, M.; Chao, X.; Zhang, C.; Yang, H.; Chen, J.; Zhou, B. Stevioside Improves Antioxidant Capacity and Intestinal Barrier Function while Attenuating Inflammation and Apoptosis by Regulating the NF-κB/MAPK Pathways in Diquat-Induced Oxidative Stress of IPEC-J2 Cells. Antioxidants 2023, 12, 1070. https://doi.org/10.3390/antiox12051070
Xu Q, Liu M, Chao X, Zhang C, Yang H, Chen J, Zhou B. Stevioside Improves Antioxidant Capacity and Intestinal Barrier Function while Attenuating Inflammation and Apoptosis by Regulating the NF-κB/MAPK Pathways in Diquat-Induced Oxidative Stress of IPEC-J2 Cells. Antioxidants. 2023; 12(5):1070. https://doi.org/10.3390/antiox12051070
Chicago/Turabian StyleXu, Qinglei, Mingzheng Liu, Xiaohuan Chao, Chunlei Zhang, Huan Yang, Jiahao Chen, and Bo Zhou. 2023. "Stevioside Improves Antioxidant Capacity and Intestinal Barrier Function while Attenuating Inflammation and Apoptosis by Regulating the NF-κB/MAPK Pathways in Diquat-Induced Oxidative Stress of IPEC-J2 Cells" Antioxidants 12, no. 5: 1070. https://doi.org/10.3390/antiox12051070
APA StyleXu, Q., Liu, M., Chao, X., Zhang, C., Yang, H., Chen, J., & Zhou, B. (2023). Stevioside Improves Antioxidant Capacity and Intestinal Barrier Function while Attenuating Inflammation and Apoptosis by Regulating the NF-κB/MAPK Pathways in Diquat-Induced Oxidative Stress of IPEC-J2 Cells. Antioxidants, 12(5), 1070. https://doi.org/10.3390/antiox12051070