
The Rhodanese PspE Converts Thiosulfate to Cellular Sulfane Sulfur in Escherichia coli

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Culture Conditions, and Reagents
2.2. Construction of RHOD Mutants
2.3. Growth Curves
2.4. Protein Expression and Purification
2.5. Thiosulfate: GSH Sulfurtransferase Activity Assay
2.6. HPLC Analysis of Total Cellular Sulfane Sulfur
2.7. SSP4 Staining for Sulfane Sulfur Detection
2.8. GSSH Preparation and Detection
2.9. Lead Acetate Strips for the Detection of Hydrogen Sulfide
2.10. HPLC Analysis for Sulfide Detection
2.11. LC-MS/MS Analysis of PspE-Cys49
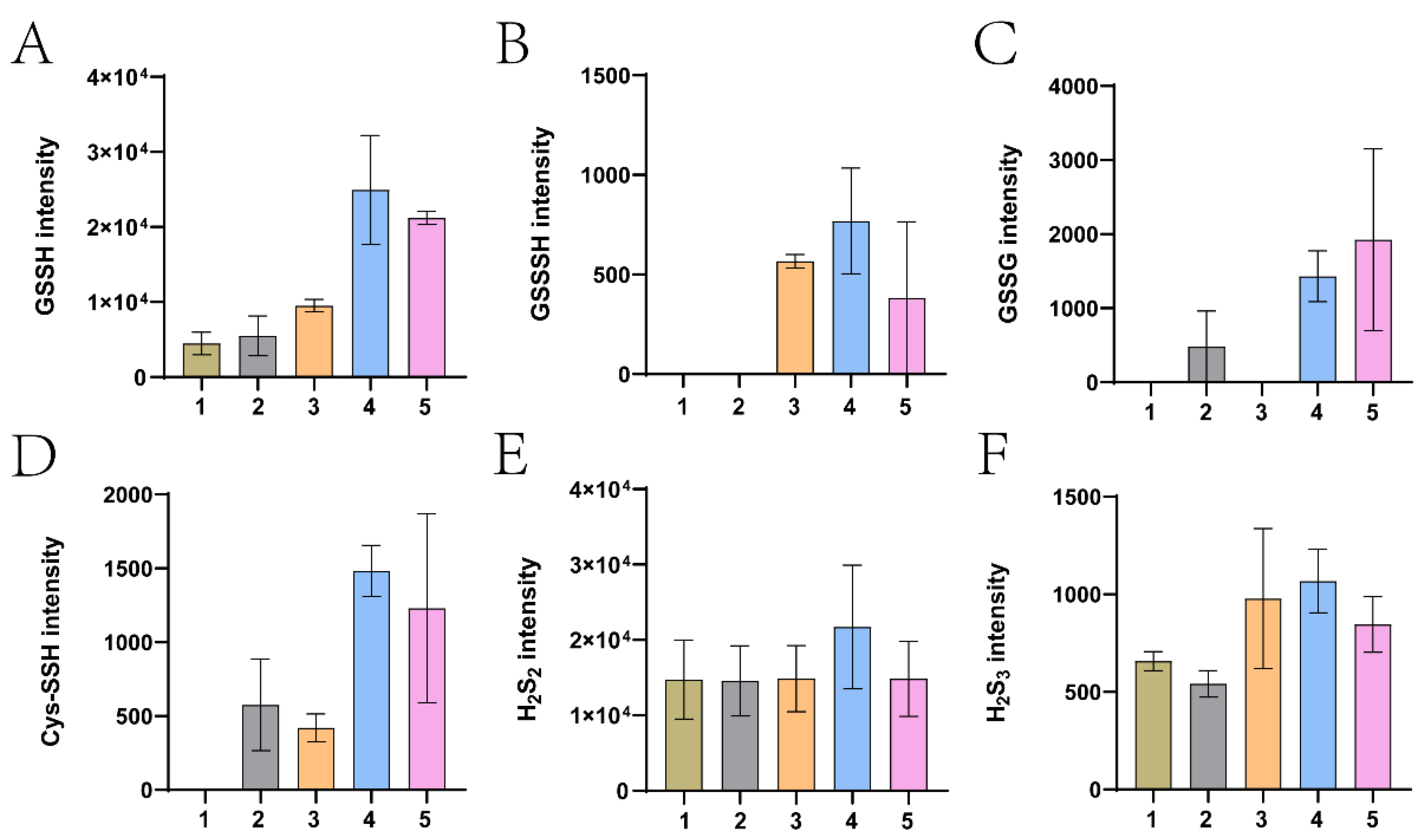
2.12. LC-MS Analysis of Cellular Sulfane Sulfur Species
3. Results
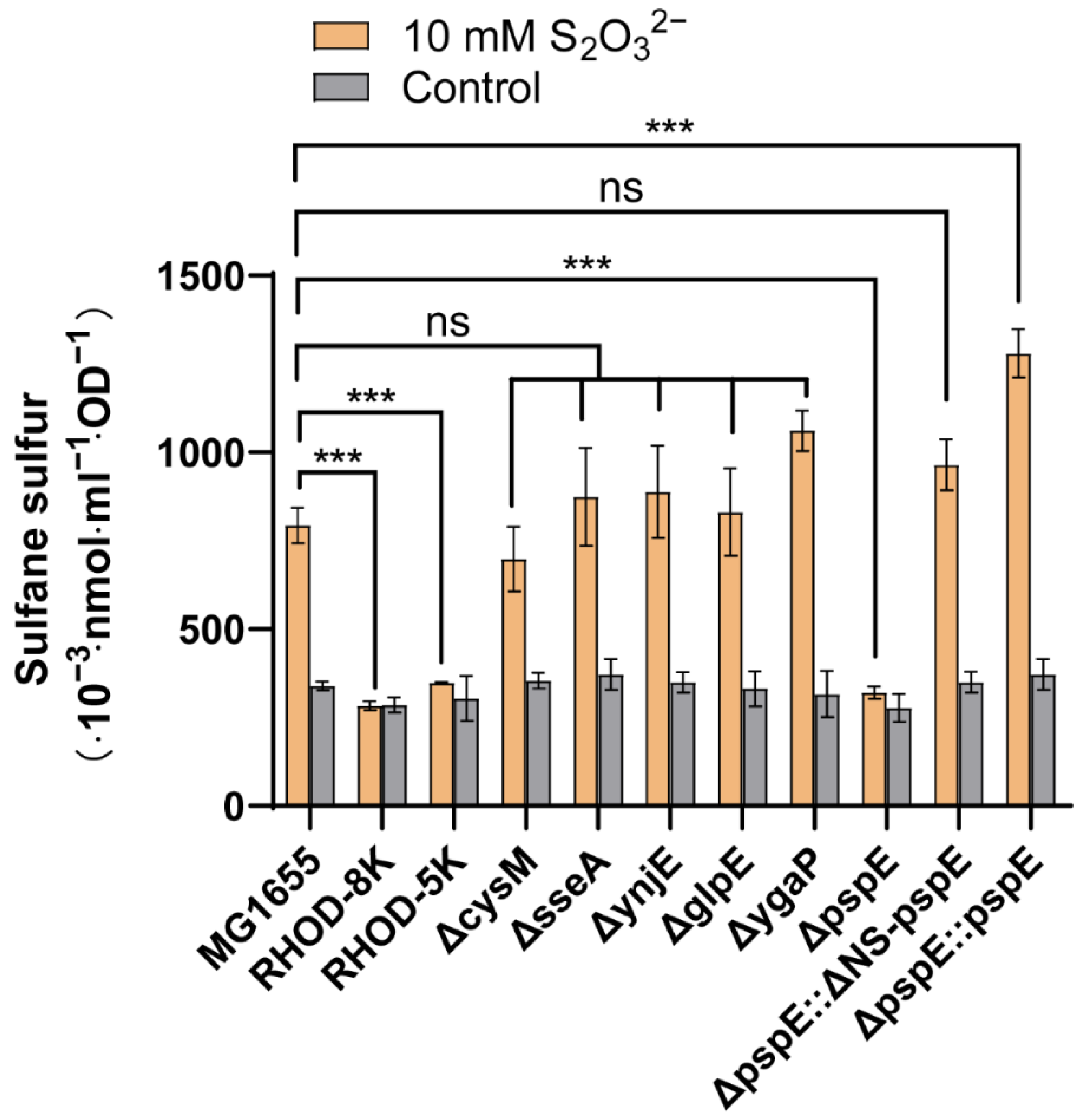
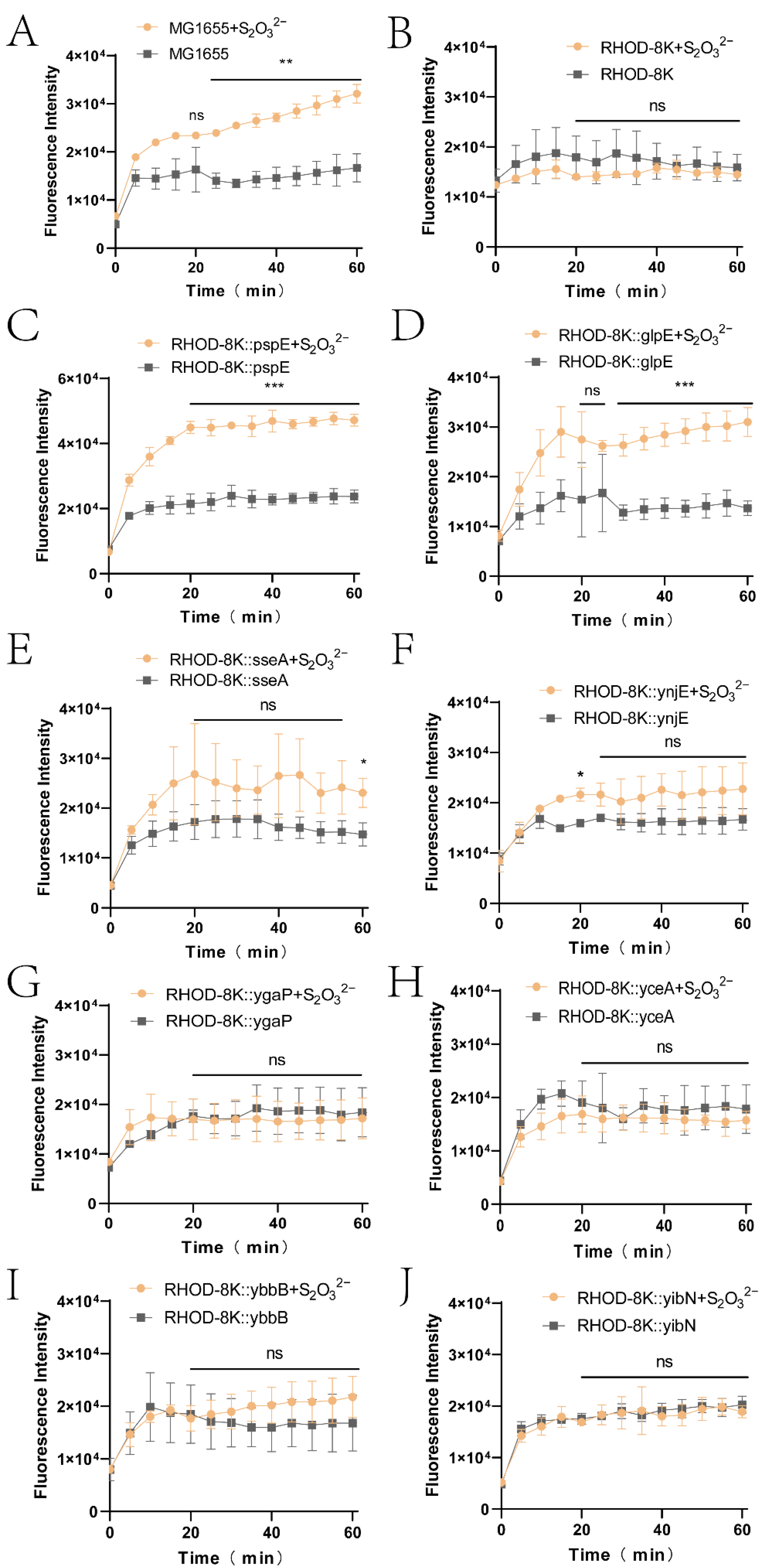
3.1. PspE was the Main Enzyme Generating Sulfane Sulfur from Thiosulfate
3.2. The Kinetics and Molecular Mechanism of PspE in Its Thiosulfate: Glutathione Sulfurtransferase Activity
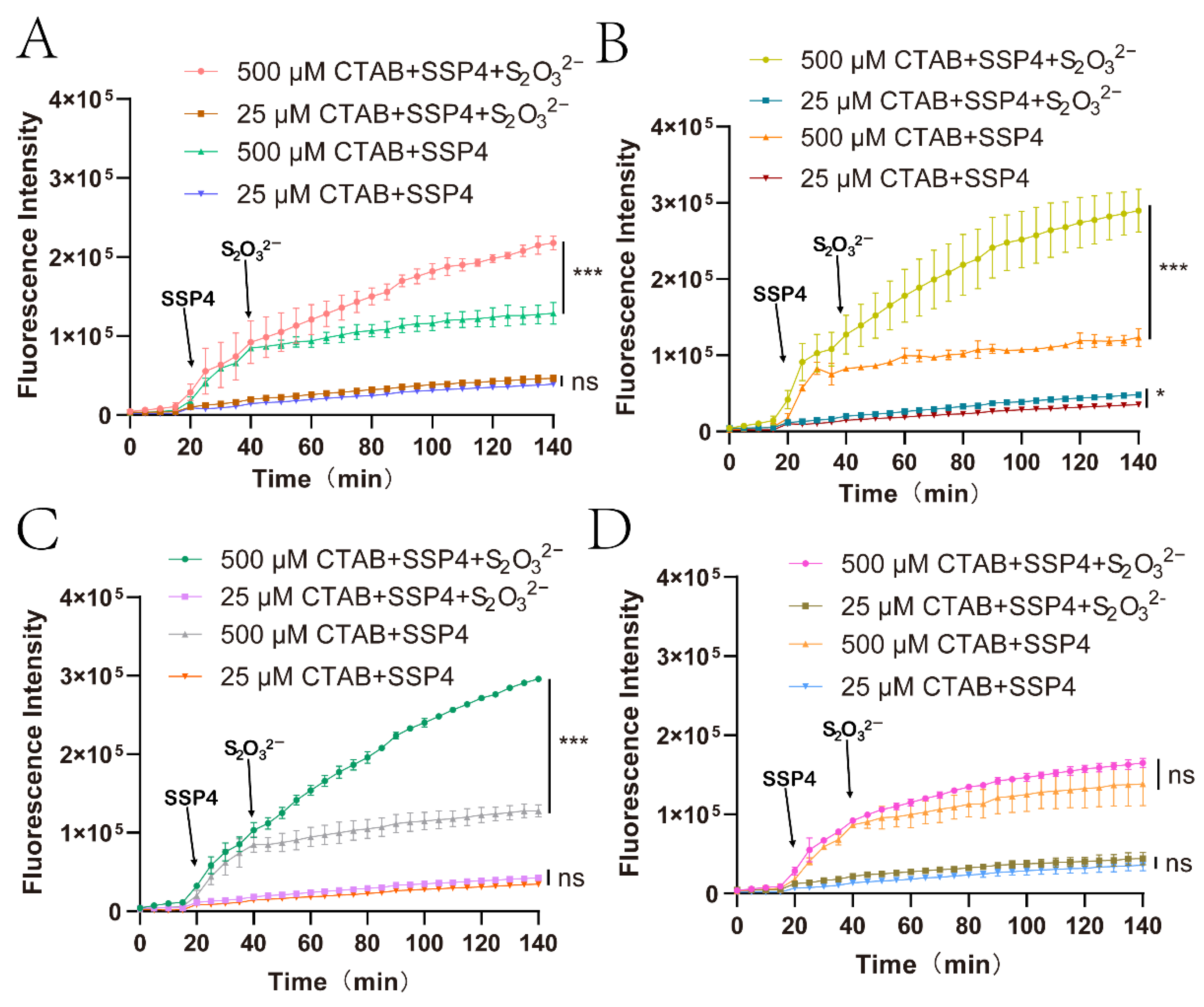
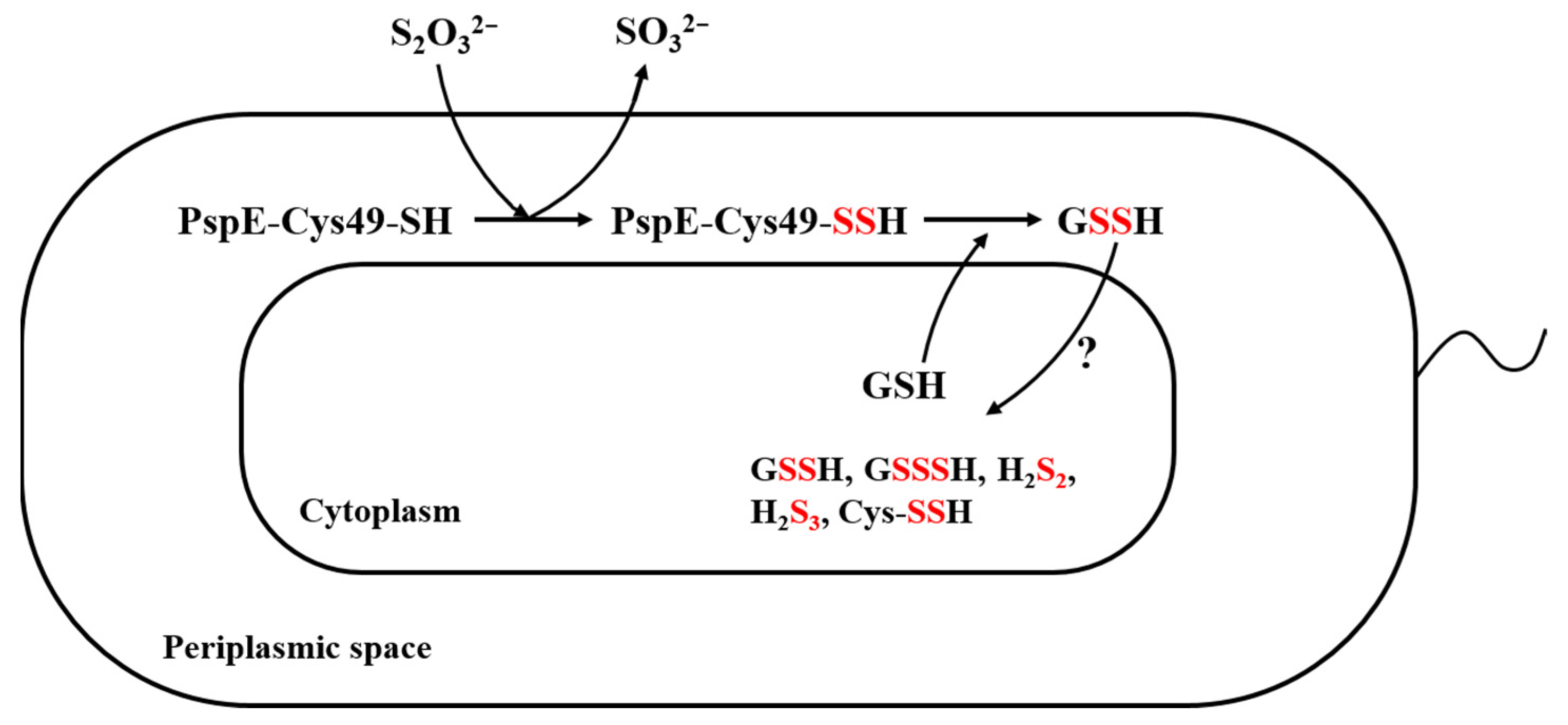
3.3. The Derived Sulfane Sulfur from Thiosulfate was Mainly Transported into the Cytoplasm
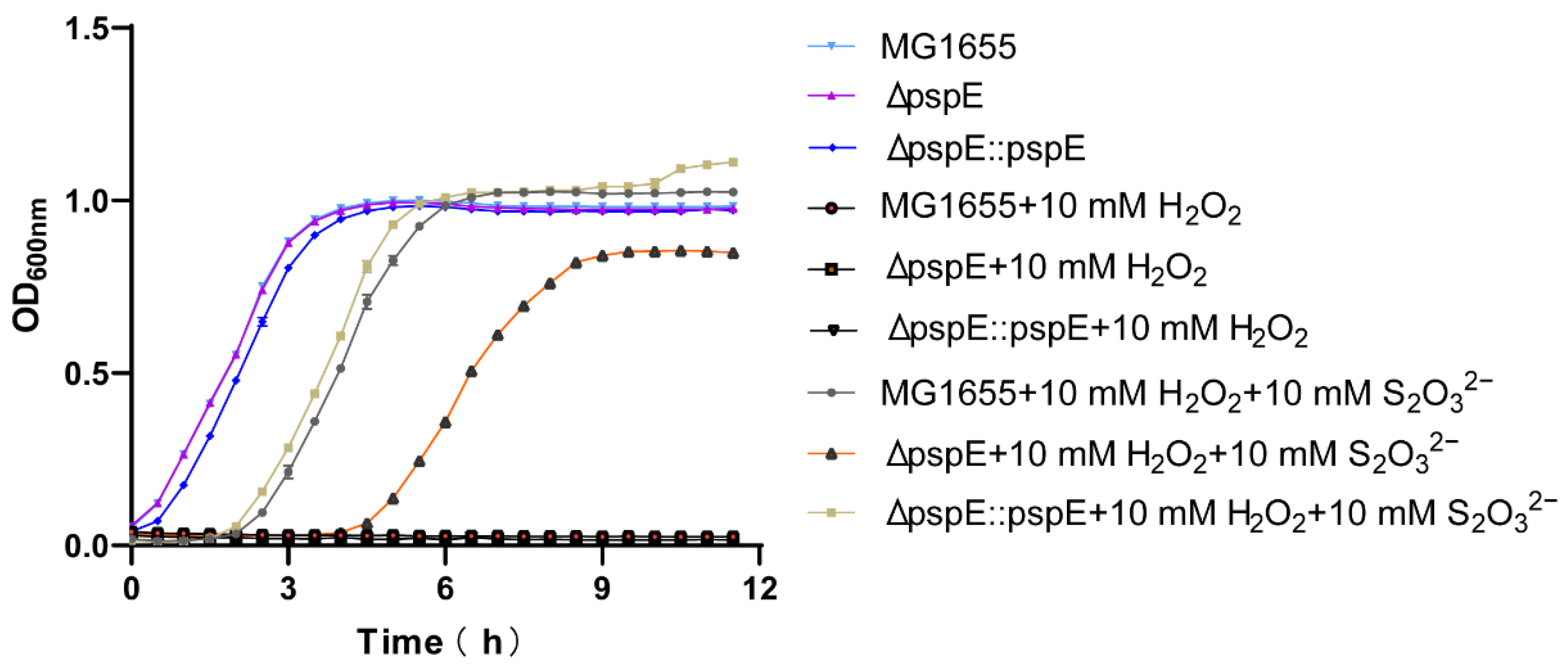
3.4. PspE was Responsible for Resisting H2O2 in the Presence of Thiosulfate in E. coli
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Takahashi, H.; Kopriva, S.; Giordano, M.; Saito, K.; Hell, R. Sulfur Assimilation in Photosynthetic Organisms: Molecular Functions and Regulations of Transporters and Assimilatory Enzymes. Annu. Rev. Plant Biol. 2011, 62, 157–184. [Google Scholar] [CrossRef] [PubMed]
- Fukuto, J.M.; Ignarro, L.J.; Nagy, P.; Wink, D.A.; Kevil, C.G.; Feelisch, M.; Cortese-Krott, M.M.; Bianco, C.L.; Kumagai, Y.; Hobbs, A.J.; et al. Biological hydropersulfides and related polysulfides—A new concept and perspective in redox biology. FEBS Lett. 2018, 592, 2140–2152. [Google Scholar] [CrossRef] [PubMed]
- Filipovic, M.R.; Zivanovic, J.; Alvarez, B.; Banerjee, R. Chemical Biology of H2S Signaling through Persulfidation. Chem. Rev. 2018, 118, 377–461. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.R.; Straub, K.D. The Role of Hydrogen Sulfide in Evolution and the Evolution of Hydrogen Sulfide in Metabolism and Signaling. Physiology 2016, 31, 60–72. [Google Scholar] [CrossRef]
- Ran, M.X.; Wang, T.Q.; Shao, M.; Chen, Z.G.; Liu, H.W.; Xia, Y.Z.; Xun, L.Y. Sensitive Method for Reliable Quantification of Sulfane Sulfur in Biological Samples. Anal. Chem. 2019, 91, 11981–11986. [Google Scholar] [CrossRef]
- Roy, B.; Shieh, M.; Ramush, G.; Xian, M. Organelle-Targeted Fluorescent Probes for Sulfane Sulfur Species. Antioxidants 2023, 12, 590. [Google Scholar] [CrossRef]
- Toohey, J.I.; Cooper, A.J. Thiosulfoxide (sulfane) sulfur: New chemistry and new regulatory roles in biology. Molecules 2014, 19, 12789–12813. [Google Scholar] [CrossRef]
- Shieh, M.; Ni, X.; Xu, S.; Lindahl, S.P.; Yang, M.; Matsunaga, T.; Flaumenhaft, R.; Akaike, T.; Xian, M. Shining a light on SSP4: A comprehensive analysis and biological applications for the detection of sulfane sulfurs. Redox Biol. 2022, 56, 102433. [Google Scholar] [CrossRef]
- Lau, N.; Pluth, M.D. Reactive sulfur species (RSS): Persulfides, polysulfides, potential, and problems. Curr. Opin. Chem. Biol. 2019, 49, 1–8. [Google Scholar] [CrossRef]
- Akaike, T.; Ida, T.; Wei, F.Y.; Nishida, M.; Kumagai, Y.; Alam, M.M.; Ihara, H.; Sawa, T.; Matsunaga, T.; Kasamatsu, S.; et al. Cysteinyl-tRNA synthetase governs cysteine polysulfidation and mitochondrial bioenergetics. Nat. Commun. 2017, 8, 1177. [Google Scholar] [CrossRef]
- Landry, A.P.; Moon, S.; Bonanata, J.; Cho, U.S.; Coitino, E.L.; Banerjee, R. Dismantling and Rebuilding the Trisulfide Cofactor Demonstrates Its Essential Role in Human Sulfide Quinone Oxidoreductase. J. Am. Chem. Soc. 2020, 142, 14295–14306. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, N.; Koike, S.; Nirasawa, T.; Kimura, H.; Ogasawara, Y. Alternative pathway of H(2)S and polysulfides production from sulfurated catalytic-cysteine of reaction intermediates of 3-mercaptopyruvate sulfurtransferase. Biochem. Biophys. Res. Commun. 2018, 496, 648–653. [Google Scholar] [CrossRef] [PubMed]
- Shatalin, K.; Nuthanakanti, A.; Kaushik, A.; Shishov, D.; Peselis, A.; Shamovsky, I.; Pani, B.; Lechpammer, M.; Vasilyev, N.; Shatalina, E.; et al. Inhibitors of bacterial H(2)S biogenesis targeting antibiotic resistance and tolerance. Science 2021, 372, 1169–1175. [Google Scholar] [CrossRef]
- Tiranti, V.; Viscomi, C.; Hildebrandt, T.; Di Meo, I.; Mineri, R.; Tiveron, C.; Levitt, M.D.; Prelle, A.; Fagiolari, G.; Rimoldi, M.; et al. Loss of ETHE1, a mitochondrial dioxygenase, causes fatal sulfide toxicity in ethylmalonic encephalopathy. Nat. Med. 2009, 15, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Pokorna, D.; Zabranska, J. Sulfur-oxidizing bacteria in environmental technology. Biotechnol. Adv. 2015, 33, 1246–1259. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.Z.; Lu, C.J.; Hou, N.K.; Xin, Y.F.; Liu, J.H.; Liu, H.L.; Xun, L.Y. Sulfide production and oxidation by heterotrophic bacteria under aerobic conditions. Isme J. 2017, 11, 2754–2766. [Google Scholar] [CrossRef]
- Xin, Y.F.; Gao, R.; Cui, F.F.; Lu, C.J.; Liu, H.L.; Liu, H.W.; Xia, Y.Z.; Xun, L.Y. The Heterotrophic Bacterium Cupriavidus pinatubonensis JMP134 Oxidizes Sulfide to Sulfate with Thiosulfate as a Key Intermediate. Appl. Environ. Microbiol. 2020, 86, e01835-20. [Google Scholar] [CrossRef]
- Xin, Y.F.; Liu, H.L.; Cui, F.F.; Liu, H.W.; Xun, L.Y. Recombinant Escherichia coli with sulfide: Quinone oxidoreductase and persulfide dioxygenase rapidly oxidises sulfide to sulfite and thiosulfate via a new pathway. Environ. Microbiol. 2016, 18, 5123–5136. [Google Scholar] [CrossRef]
- Shen, J.; Peng, H.; Zhang, Y.; Trinidad, J.C.; Giedroc, D.P. Staphylococcus aureus sqr Encodes a Type II Sulfide:Quinone Oxidoreductase and Impacts Reactive Sulfur Speciation in Cells. Biochemistry 2016, 55, 6524–6534. [Google Scholar] [CrossRef]
- Landry, A.P.; Ballou, D.P.; Banerjee, R. Hydrogen Sulfide Oxidation by Sulfide Quinone Oxidoreductase. Chembiochem 2021, 22, 949–960. [Google Scholar] [CrossRef]
- Zhang, X.; Xin, Y.P.; Chen, Z.G.; Xia, Y.Z.; Xun, L.Y.; Liu, H.W. Sulfide-quinone oxidoreductase is required for cysteine synthesis and indispensable to mitochondrial health. Redox Biol. 2021, 47, 102169. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.R.; Melideo, S.L.; Jorns, M.S. Role of Human Sulfide: Quinone Oxidoreductase in H2S Metabolism. Method Enzymol. 2015, 554, 255–270. [Google Scholar] [CrossRef]
- Shen, J.C.; Keithly, M.E.; Armstrong, R.N.; Higgins, K.A.; Edmonds, K.A.; Giedroc, D.P. Staphylococcus aureus CstB Is a Novel Multidomain Persulfide Dioxygenase-Sulfurtransferase Involved in Hydrogen Sulfide Detoxification. Biochemistry 2015, 54, 4542–4554. [Google Scholar] [CrossRef] [PubMed]
- Ran, M.X.; Li, Q.B.; Xin, Y.F.; Ma, S.H.; Zhao, R.; Wang, M.; Xun, L.Y.; Xia, Y.Z. Rhodaneses minimize the accumulation of cellular sulfane sulfur to avoid disulfide stress during sulfide oxidation in bacteria. Redox Biol. 2022, 53, 102345. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.L.; Xin, Y.F.; Xun, L.Y. Distribution, Diversity, and Activities of Sulfur Dioxygenases in Heterotrophic Bacteria. Appl. Environ. Microbiol. 2014, 80, 1799–1806. [Google Scholar] [CrossRef] [PubMed]
- Doka, E.; Pader, I.; Biro, A.; Johansson, K.; Cheng, Q.; Ballago, K.; Prigge, J.R.; Pastor-Flores, D.; Dick, T.P.; Schmidt, E.E.; et al. A novel persulfide detection method reveals protein persulfide- and polysulfide-reducing functions of thioredoxin and glutathione systems. Sci. Adv. 2016, 2, e1500968. [Google Scholar] [CrossRef]
- Wang, T.Q.; Ran, M.X.; Li, X.J.; Liu, Y.Q.; Xin, Y.F.; Liu, H.L.; Liu, H.W.; Xia, Y.Z.; Xun, L.Y. The Pathway of Sulfide Oxidation to Octasulfur Globules in the Cytoplasm of Aerobic Bacteria. Appl. Environ. Microbiol. 2022, 88, e0194121. [Google Scholar] [CrossRef]
- Powell, C.R.; Dillon, K.M.; Matson, J.B. A review of hydrogen sulfide (H2S) donors: Chemistry and potential therapeutic applications. Biochem. Pharmacol. 2018, 149, 110–123. [Google Scholar] [CrossRef]
- Levinn, C.M.; Cerda, M.M.; Pluth, M.D. Activatable Small-Molecule Hydrogen Sulfide Donors. Antioxid. Redox Signal. 2020, 32, 96–109. [Google Scholar] [CrossRef]
- Ni, X.; Kelly, S.S.; Xu, S.; Xian, M. The Path to Controlled Delivery of Reactive Sulfur Species. Accounts Chem. Res. 2021, 54, 3968–3978. [Google Scholar] [CrossRef]
- Merz, T.; McCook, O.; Brucker, C.; Waller, C.; Calzia, E.; Radermacher, P.; Datzmann, T. H2S in Critical Illness-A New Horizon for Sodium Thiosulfate? Biomolecules 2022, 12, 543. [Google Scholar] [CrossRef] [PubMed]
- Petrikovics, I.; Budai, M.; Kovacs, K.; Thompson, D.E. Past, present and future of cyanide antagonism research: From the early remedies to the current therapies. World J. Methodol. 2015, 5, 88–100. [Google Scholar] [CrossRef]
- Yang, X.; Liu, Y.Q.; Xie, X.T.; Shi, W.; Si, J.Y.; Li, X.M.; Zhang, X.L.; Liu, B.C. Use of the optimized sodium thiosulfate regimen for the treatment of calciphylaxis in Chinese patients. Renal Fail. 2022, 44, 914–922. [Google Scholar] [CrossRef]
- Tsang, R.Y.; Al-Fayea, T.; Au, H.J. Cisplatin Overdose Toxicities and Management. Drug Saf. 2009, 32, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, T.M.; Grieshaber, M.K. Three enzymatic activities catalyze the oxidation of sulfide to thiosulfate in mammalian and invertebrate mitochondria. FEBS J. 2008, 275, 3352–3361. [Google Scholar] [CrossRef] [PubMed]
- Mikami, Y.; Shibuya, N.; Kimura, Y.; Nagahara, N.; Ogasawara, Y.; Kimura, H. Thioredoxin and dihydrolipoic acid are required for 3-mercaptopyruvate sulfurtransferase to produce hydrogen sulfide. Biochem. J. 2011, 439, 479–485. [Google Scholar] [CrossRef]
- Villarejo, M.; Westley, J. Mechanism of Rhodanese Catalysis of Thiosulfate-Lipoate Oxidation-Reduction. J. Biol. Chem. 1963, 238, 4016–4020. [Google Scholar] [CrossRef]
- Tai, C.H.; Nalabolu, S.R.; Jacobson, T.M.; Minter, D.E.; Cook, P.F. Kinetic mechanisms of the A and B isozymes of O-acetylserine sulfhydrylase from Salmonella typhimurium LT-2 using the natural and alternative reactants. Biochemistry 1993, 32, 6433–6442. [Google Scholar] [CrossRef]
- Li, K.; Xin, Y.F.; Xuan, G.H.; Zhao, R.; Liu, H.W.; Xia, Y.Z.; Xun, L.Y. Escherichia coli Uses Separate Enzymes to Produce H2S and Reactive Sulfane Sulfur From L-cysteine. Front. Microbiol. 2019, 10, 298. [Google Scholar] [CrossRef]
- Chen, Z.G.; Zhang, X.; Li, H.J.; Liu, H.W.; Xia, Y.Z.; Xun, L.Y. The Complete Pathway for Thiosulfate Utilization in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2018, 84, e01241-18. [Google Scholar] [CrossRef]
- Kruithof, P.D.; Lunev, S.; Aguilar Lozano, S.P.; de Assis Batista, F.; Al-Dahmani, Z.M.; Joles, J.A.; Dolga, A.M.; Groves, M.R.; van Goor, H. Unraveling the role of thiosulfate sulfurtransferase in metabolic diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165716. [Google Scholar] [CrossRef] [PubMed]
- Westley, J. Rhodanese. Adv. Enzymol. Ramb. 1973, 39, 327–368. [Google Scholar]
- Cheng, H.; Donahue, J.L.; Battle, S.E.; Ray, W.K.; Larson, T.J. Biochemical and Genetic Characterization of PspE and GlpE, Two Single-domain Sulfurtransferases of Escherichia coli. Open Microbiol. J. 2008, 2, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Hanzelmann, P.; Dahl, J.U.; Kuper, J.; Urban, A.; Muller-Theissen, U.; Leimkuhler, S.; Schindelin, H. Crystal structure of YnjE from Escherichia coli, a sulfurtransferase with three rhodanese domains. Protein Sci. 2009, 18, 2480–2491. [Google Scholar] [CrossRef] [PubMed]
- Melideo, S.L.; Jackson, M.R.; Jorns, M.S. Biosynthesis of a Central Intermediate in Hydrogen Sulfide Metabolism by a Novel Human Sulfurtransferase and Its Yeast Ortholog. Biochemistry 2014, 53, 4739–4753. [Google Scholar] [CrossRef]
- Ploegman, J.H.; Drent, G.; Kalk, K.H.; Hol, W.G.; Heinrikson, R.L.; Keim, P.; Weng, L.; Russell, J. The covalent and tertiary structure of bovine liver rhodanese. Nature 1978, 273, 124–129. [Google Scholar] [CrossRef]
- Cartini, F.; Remelli, W.; Dos Santos, P.C.; Papenbrock, J.; Pagani, S.; Forlani, F. Mobilization of sulfane sulfur from cysteine desulfurases to the Azotobacter vinelandii sulfurtransferase RhdA. Amino Acids 2011, 41, 141–150. [Google Scholar] [CrossRef]
- Blattner, F.R.; Plunkett, G., 3rd; Bloch, C.A.; Perna, N.T.; Burland, V.; Riley, M.; Collado-Vides, J.; Glasner, J.D.; Rode, C.K.; Mayhew, G.F.; et al. The complete genome sequence of Escherichia coli K-12. Science 1997, 277, 1453–1462. [Google Scholar] [CrossRef]
- Yu, Q.L.; Ran, M.X.; Yang, Y.Q.; Liu, H.W.; Xun, L.Y.; Xia, Y.Z. Optimization of a Method for Detecting Intracellular Sulfane Sulfur Levels and Evaluation of Reagents That Affect the Levels in Escherichia coli. Antioxidants 2022, 11, 1292. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef]
- Li, M.; Wang, J.; Geng, Y.; Li, Y.; Wang, Q.; Liang, Q.; Qi, Q. A strategy of gene overexpression based on tandem repetitive promoters in Escherichia coli. Microb. Cell Factories 2012, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Kovach, M.E.; Elzer, P.H.; Hill, D.S.; Robertson, G.T.; Farris, M.A.; Roop, R.M., 2nd; Peterson, K.M. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Li, K.; Li, J.; Wang, T.; Gu, L.; Xun, L. T5 exonuclease-dependent assembly offers a low-cost method for efficient cloning and site-directed mutagenesis. Nucleic Acids Res. 2019, 47, e15. [Google Scholar] [CrossRef]
- Libiad, M.; Yadav, P.K.; Vitvitsky, V.; Martinov, M.; Banerjee, R. Organization of the human mitochondrial hydrogen sulfide oxidation pathway. J. Biol. Chem. 2014, 289, 30901–30910. [Google Scholar] [CrossRef]
- Adams, H.; Teertstra, W.; Koster, M.; Tommassen, J. PspE (phage-shock protein E) of Escherichia coli is a rhodanese. FEBS Lett. 2002, 518, 173–176. [Google Scholar] [CrossRef]
- Volkmer, B.; Heinemann, M. Condition-Dependent Cell Volume and Concentration of Escherichia coli to Facilitate Data Conversion for Systems Biology Modeling. PLoS ONE 2011, 6, e23126. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yang, Y.; Liu, M.; Liu, H.; Liu, H.; Xia, Y.; Xun, L. Elemental Sulfur Inhibits Yeast Growth via Producing Toxic Sulfide and Causing Disulfide Stress. Antioxidants 2022, 11, 576. [Google Scholar] [CrossRef]
- Spallarossa, A.; Donahue, J.L.; Larson, T.J.; Bolognesi, M.; Bordo, D. Escherichia coli GlpE is a prototype sulfurtransferase for the single-domain rhodanese homology superfamily. Structure 2001, 9, 1117–1125. [Google Scholar] [CrossRef]
- Alt, E.; Leipold, F.; Milatovic, D.; Lehmann, G.; Heinz, S.; Schomig, A. Hydrogen peroxide for prevention of bacterial growth on polymer biomaterials. Ann. Thorac. Surg. 1999, 68, 2123–2128. [Google Scholar] [CrossRef]
- Rao, S.P.; Dobariya, P.; Bellamkonda, H.; More, S.S. Role of 3-Mercaptopyruvate Sulfurtransferase (3-MST) in Physiology and Disease. Antioxidants 2023, 12, 603. [Google Scholar] [CrossRef]
- Colnaghi, R.; Cassinelli, G.; Drummond, M.; Forlani, F.; Pagani, S. Properties of the Escherichia coli rhodanese-like protein SseA: Contribution of the active-site residue Ser240 to sulfur donor recognition. FEBS Lett. 2001, 500, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, M.D.; Ahmed, F.; Lacourciere, G.M.; Lauhon, C.T.; Stadtman, T.C.; Larson, T.J. Functional diversity of the rhodanese homology domain: The Escherichia coli ybbB gene encodes a selenophosphate-dependent tRNA 2-selenouridine synthase. J. Biol. Chem. 2004, 279, 1801–1809. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhou, P.; He, Y.; Yu, L.; Xiong, Y.; Tian, C.L.; Wu, F.M. Fast conformational exchange between the sulfur-free and persulfide-bound rhodanese domain of E. coli YgaP. Biochem. Biophys. Res. Commun. 2014, 452, 817–821. [Google Scholar] [CrossRef] [PubMed]
- Jacquoilleot, S.; Sheffield, D.; Olayanju, A.; Sison-Young, R.; Kitteringham, N.R.; Naisbitt, D.J.; Aleksic, M. Glutathione metabolism in the HaCaT cell line as a model for the detoxification of the model sensitisers 2,4-dinitrohalobenzenes in human skin. Toxicol. Lett. 2015, 237, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Pittman, M.S.; Robinson, H.C.; Poole, R.K. A bacterial glutathione transporter (Escherichia coli CydDC) exports reductant to the periplasm. J. Biol. Chem. 2005, 280, 32254–32261. [Google Scholar] [CrossRef] [PubMed]
- Hosoki, R.; Matsuki, N.; Kimura, H. The possible role of hydrogen sulfide as an endogenous smooth muscle relaxant in synergy with nitric oxide. Biochem. Biophys. Res. Commun. 1997, 237, 527–531. [Google Scholar] [CrossRef]
- Aminzadeh, M.A.; Vaziri, N.D. Downregulation of the renal and hepatic hydrogen sulfide (H2S)-producing enzymes and capacity in chronic kidney disease. Nephrol. Dial. Transplant. 2012, 27, 498–504. [Google Scholar] [CrossRef]
- Yang, G.D.; Wu, L.Y.; Jiang, B.; Yang, W.; Qi, J.S.; Cao, K.; Meng, Q.H.; Mustafa, A.K.; Mu, W.T.; Zhang, S.M.; et al. H2S as a physiologic vasorelaxant: Hypertension in mice with deletion of cystathionine gamma-lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef]
- Lv, B.Y.; Chen, S.L.N.; Tang, C.S.; Jin, H.F.; Du, J.B.; Huang, Y.Q. Hydrogen sulfide and vascular regulation—An update. J. Adv. Res. 2021, 27, 85–97. [Google Scholar] [CrossRef]
- Zhao, W.M.; Zhang, J.; Lu, Y.J.; Wang, R. The vasorelaxant effect of H2S as a novel endogenous gaseous K-ATP channel opener. Embo J. 2001, 20, 6008–6016. [Google Scholar] [CrossRef]
- Mueller, E.G. Trafficking in persulfides: Delivering sulfur in biosynthetic pathways. Nat. Chem. Biol. 2006, 2, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.K.; Gadalla, M.M.; Sen, N.; Kim, S.; Mu, W.T.; Gazi, S.K.; Barrow, R.K.; Yang, G.D.; Wang, R.; Snyder, S.H. H2S Signals through Protein S-Sulfhydration. Sci. Signal. 2009, 2, ra72. [Google Scholar] [CrossRef] [PubMed]
- Walsh, B.J.C.; Costa, S.S.; Edmonds, K.A.; Trinidad, J.C.; Issoglio, F.M.; Brito, J.A.; Giedroc, D.P. Metabolic and Structural Insights into Hydrogen Sulfide Mis-Regulation in Enterococcus faecalis. Antioxidants 2022, 11, 1607. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, J.; Lu, C.; Xia, Y.; Xin, Y.; Liu, H.; Xun, L.; Liu, H. FisR activates sigma(54) -dependent transcription of sulfide-oxidizing genes in Cupriavidus pinatubonensis JMP134. Mol. Microbiol. 2017, 105, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.W.; Xuan, G.H.; Liu, H.W.; Xia, Y.Z.; Xun, L.Y. Sulfane Sulfur Is a Strong Inducer of the Multiple Antibiotic Resistance Regulator MarR in Escherichia coli. Antioxidants 2021, 10, 1778. [Google Scholar] [CrossRef]
- Xuan, G.H.; Lu, C.J.; Xu, H.W.; Chen, Z.G.; Li, K.; Liu, H.L.; Liu, H.W.; Xia, Y.Z.; Xun, L.Y. Sulfane Sulfur is an intrinsic signal activating MexR-regulated antibiotic resistance in Pseudomonas aeruginosa. Mol. Microbiol. 2020, 114, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Biggs, T.D.; Xian, M. Hydrogen sulfide (H2S) releasing agents: Chemistry and biological applications. Chem. Commun. 2014, 50, 11788–11805. [Google Scholar] [CrossRef]
- Li, Z.; Xia, H.; Sharp, T.E., 3rd; LaPenna, K.B.; Elrod, J.W.; Casin, K.M.; Liu, K.; Calvert, J.W.; Chau, V.Q.; Salloum, F.N.; et al. Mitochondrial H(2)S Regulates BCAA Catabolism in Heart Failure. Circ. Res. 2022, 131, 222–235. [Google Scholar] [CrossRef]
- Kondo, K.; Bhushan, S.; King, A.L.; Prabhu, S.D.; Hamid, T.; Koenig, S.; Murohara, T.; Predmore, B.L.; Gojon, G.; Gojon, G.; et al. H2S Protects Against Pressure Overload-Induced Heart Failure via Upregulation of Endothelial Nitric Oxide Synthase. Circulation 2013, 127, 1116–1127. [Google Scholar] [CrossRef]
- Snijder, P.M.; Frenay, A.R.; de Boer, R.A.; Pasch, A.; Hillebrands, J.L.; Leuvenink, H.G.D.; van Goor, H. Exogenous administration of thiosulfate, a donor of hydrogen sulfide, attenuates angiotensin II-induced hypertensive heart disease in rats. Brit. J. Pharmacol. 2015, 172, 1494–1504. [Google Scholar] [CrossRef]
- Li, H.; Liu, H.; Chen, Z.; Zhao, R.; Wang, Q.; Ran, M.; Xia, Y.; Hu, X.; Liu, J.; Xian, M.; et al. Using resonance synchronous spectroscopy to characterize the reactivity and electrophilicity of biologically relevant sulfane sulfur. Redox Biol. 2019, 24, 101179. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.K.; Gadalla, M.M.; Snyder, S.H. Signaling by Gasotransmitters. Sci. Signal. 2009, 2, re2. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RHODs | Km (GSH) mM | Km (S2O32−) mM | Vmax nmol·min−1·mg−1 | kcat s−1 | kcat/Km (GSH) M−1 s−1 | kcat/Km (S2O32−) M−1 s−1 |
|---|---|---|---|---|---|---|
| ΔNS-PspE | 28.9 ± 3.0 | 7.8 ± 1.6 | 212.7 ± 13.9 | 0.04 | 1.3 | 4.8 |
| GlpE | 14.3 ± 0.1 | 33.6 ± 7.0 | 109.1 ± 12.2 | 0.02 | 1.7 | 0.7 |
| ΔNS-YnjE | - | - | - | - | - | - |
| YceA | - | - | - | - | - | - |
| Sequence | Protein | Modification | Intensity | Percentage (%) |
|---|---|---|---|---|
| IATAVPDKNDTVKVYCNAGR | ΔNS-PspE + S2O32− | Cys-SH | 7,288,433,560 | 98.9 |
| Cys-SSH | 79,902,321 | 1.1 | ||
| ΔNS-PspE | Cys-SH | 121,956,774 | 100 | |
| Cys-SSH | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Q.; Ran, M.; Xin, Y.; Liu, H.; Liu, H.; Xia, Y.; Xun, L. The Rhodanese PspE Converts Thiosulfate to Cellular Sulfane Sulfur in Escherichia coli. Antioxidants 2023, 12, 1127. https://doi.org/10.3390/antiox12051127
Yu Q, Ran M, Xin Y, Liu H, Liu H, Xia Y, Xun L. The Rhodanese PspE Converts Thiosulfate to Cellular Sulfane Sulfur in Escherichia coli. Antioxidants. 2023; 12(5):1127. https://doi.org/10.3390/antiox12051127
Chicago/Turabian StyleYu, Qiaoli, Mingxue Ran, Yuping Xin, Huaiwei Liu, Honglei Liu, Yongzhen Xia, and Luying Xun. 2023. "The Rhodanese PspE Converts Thiosulfate to Cellular Sulfane Sulfur in Escherichia coli" Antioxidants 12, no. 5: 1127. https://doi.org/10.3390/antiox12051127
APA StyleYu, Q., Ran, M., Xin, Y., Liu, H., Liu, H., Xia, Y., & Xun, L. (2023). The Rhodanese PspE Converts Thiosulfate to Cellular Sulfane Sulfur in Escherichia coli. Antioxidants, 12(5), 1127. https://doi.org/10.3390/antiox12051127