


Differences in Physiological Responses of Two Tomato Genotypes to Combined Waterlogging and Cadmium Stresses

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Experimental Treatments
2.3. Measurements
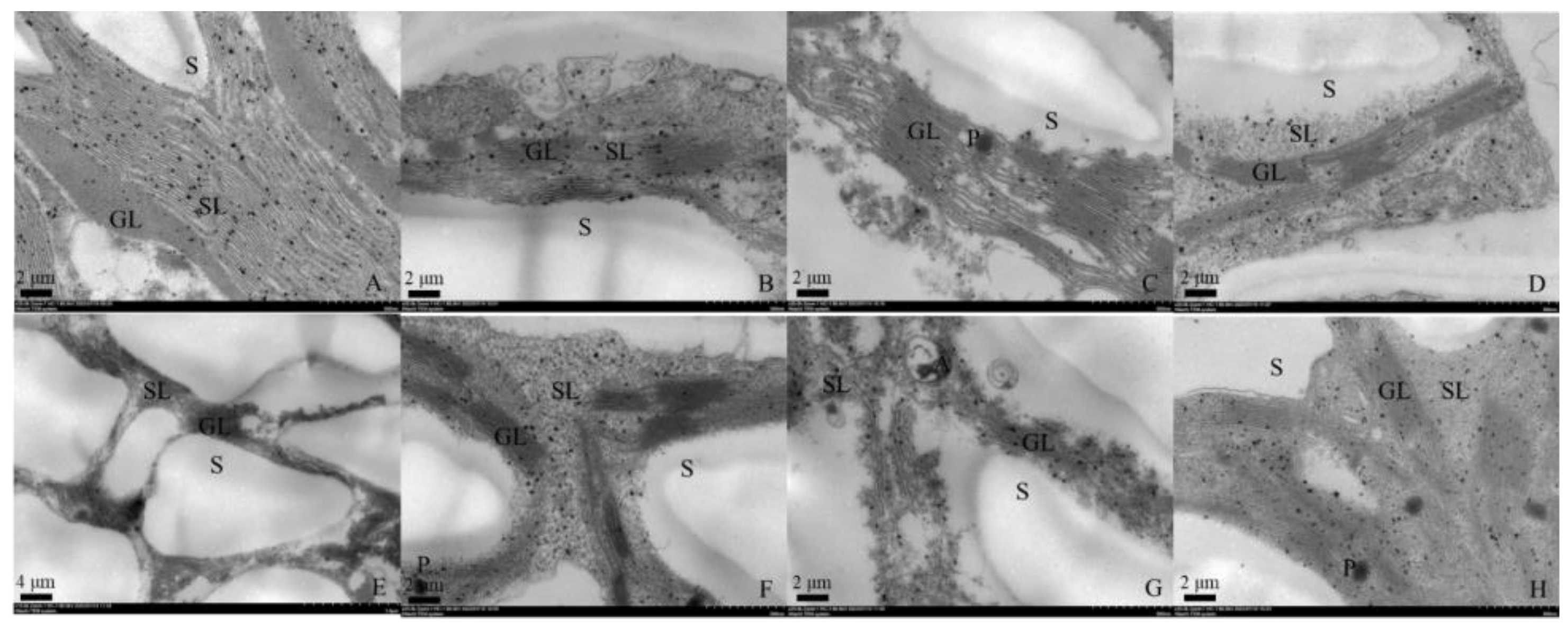
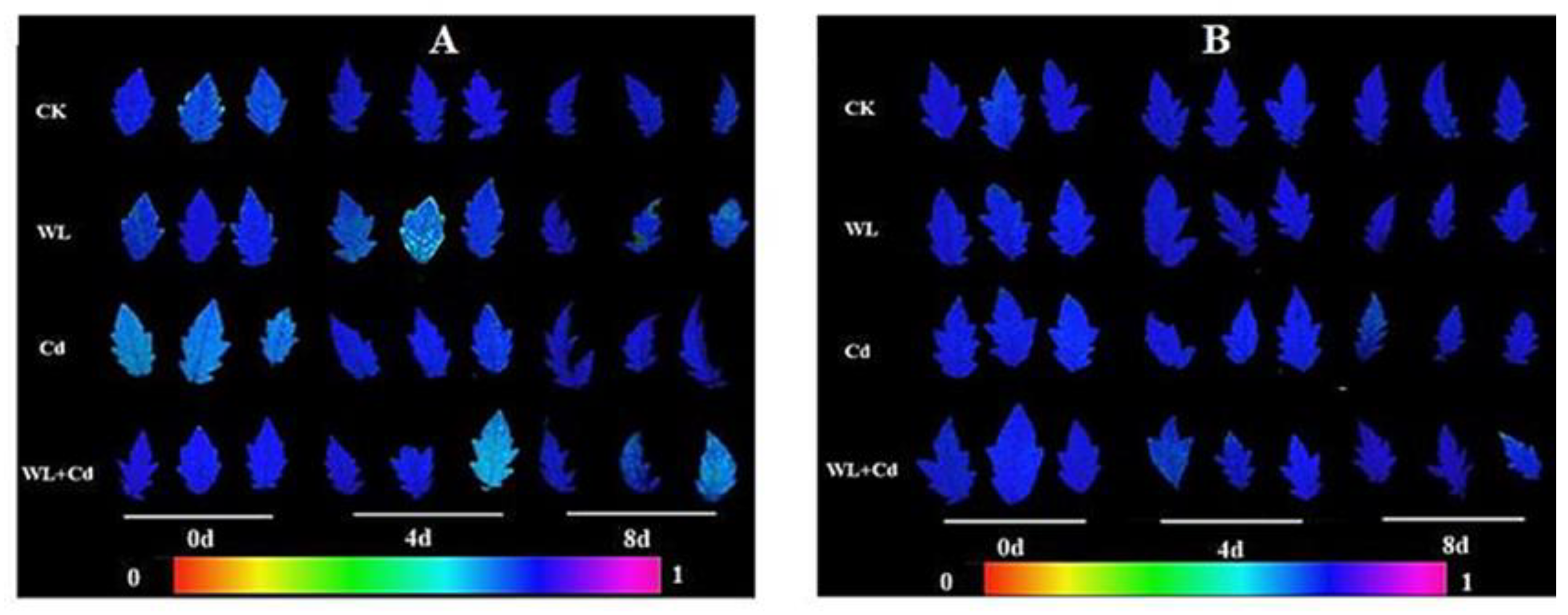
2.3.1. Chloroplast Ultrastructure and Chlorophyll Fluorescence
2.3.2. MDA, H2O2, Proline Content and O2·− Production Rate
2.3.3. Antioxidant Enzyme Activities
2.3.4. Plant Morphology and Biomass Accumulation
2.4. Data Analysis
3. Results
3.1. Chloroplast Ultrastructure Observation and Chlorophyll Fluorescence Measurements
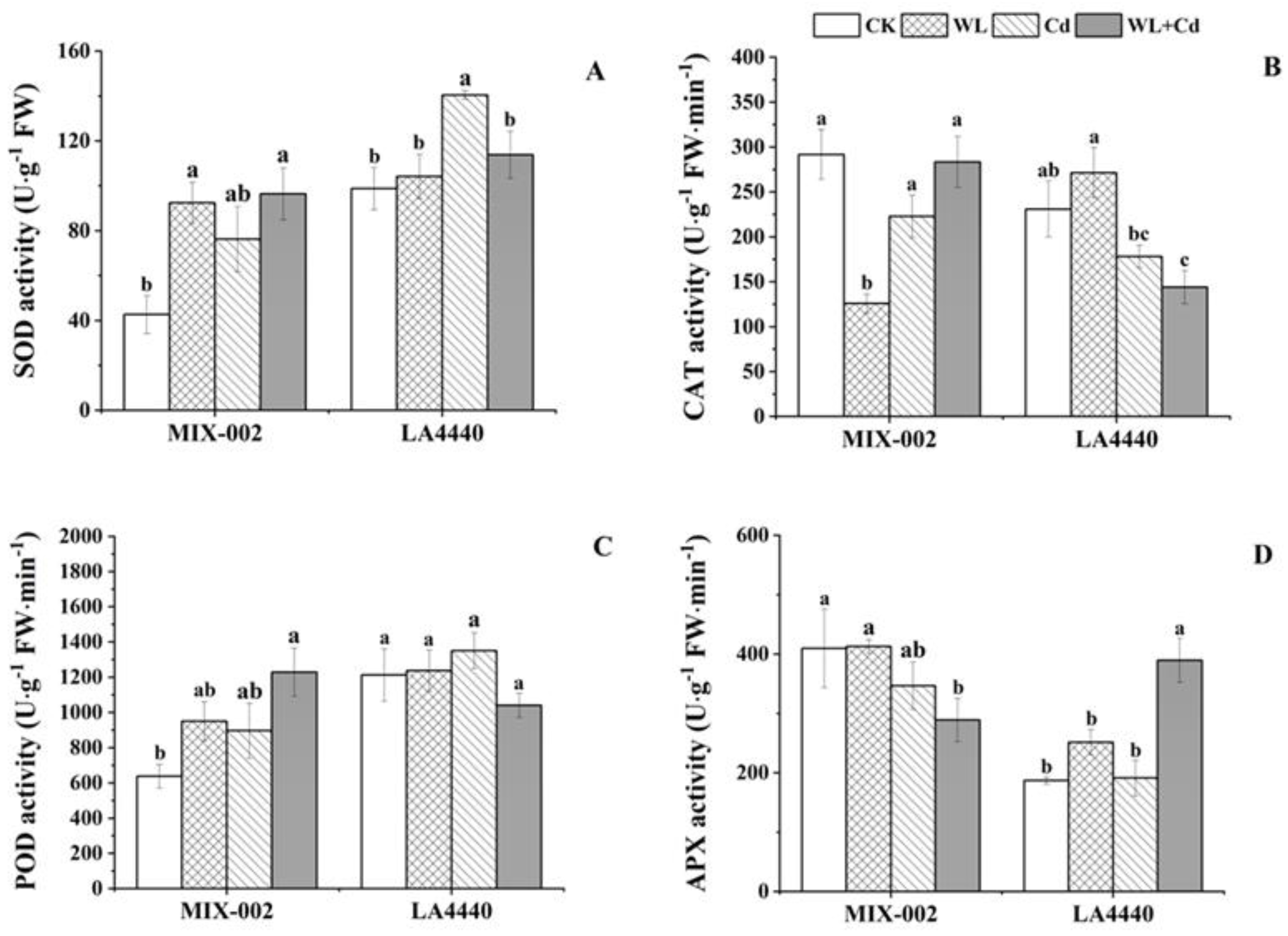
3.2. Key Regulators in Antioxidants System
3.3. Antioxidant Enzyme Activities
3.4. Biomass Accumulation
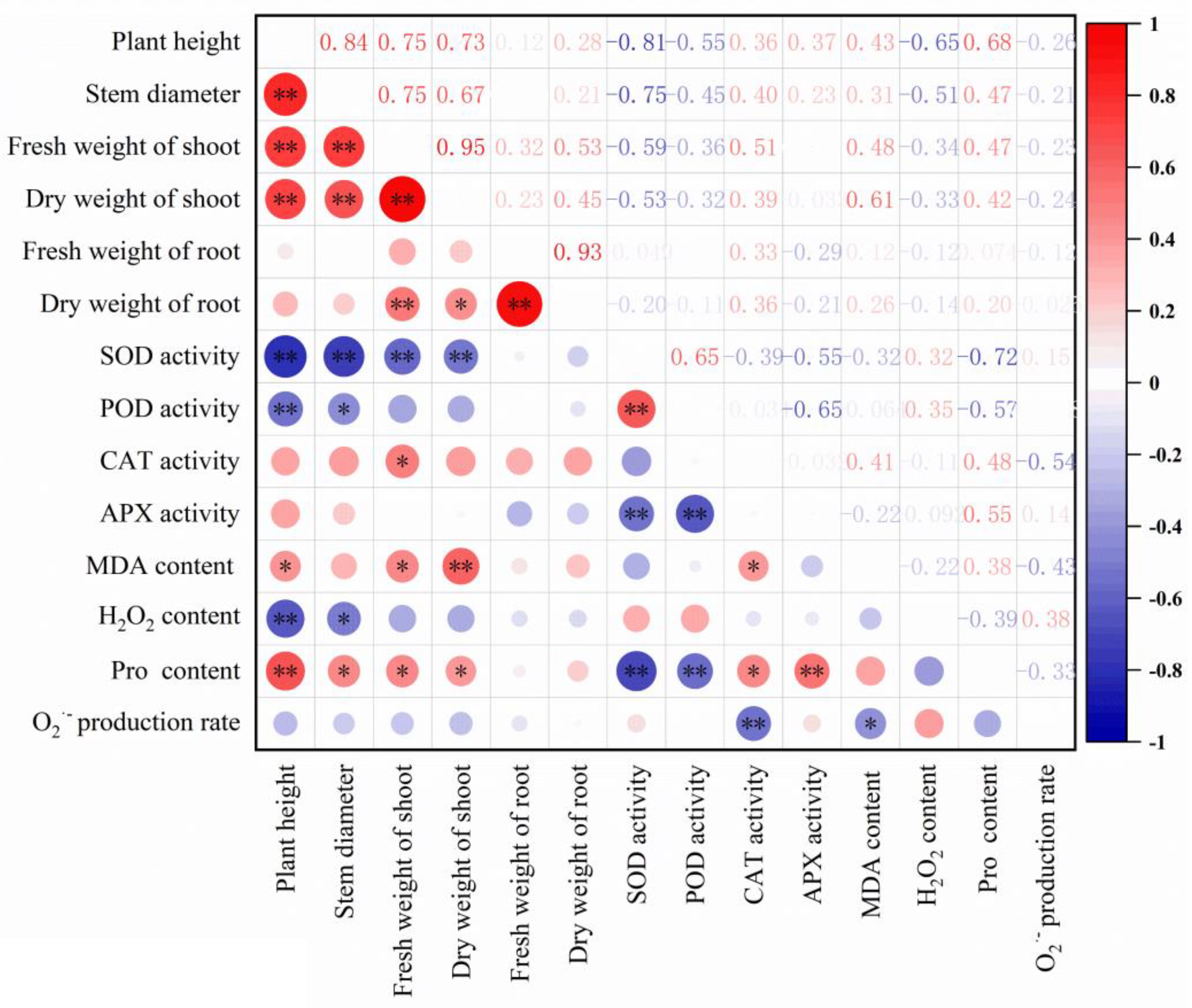
3.5. Effects of Individual Factors and Their Interactions on the Parameters
4. Discussion
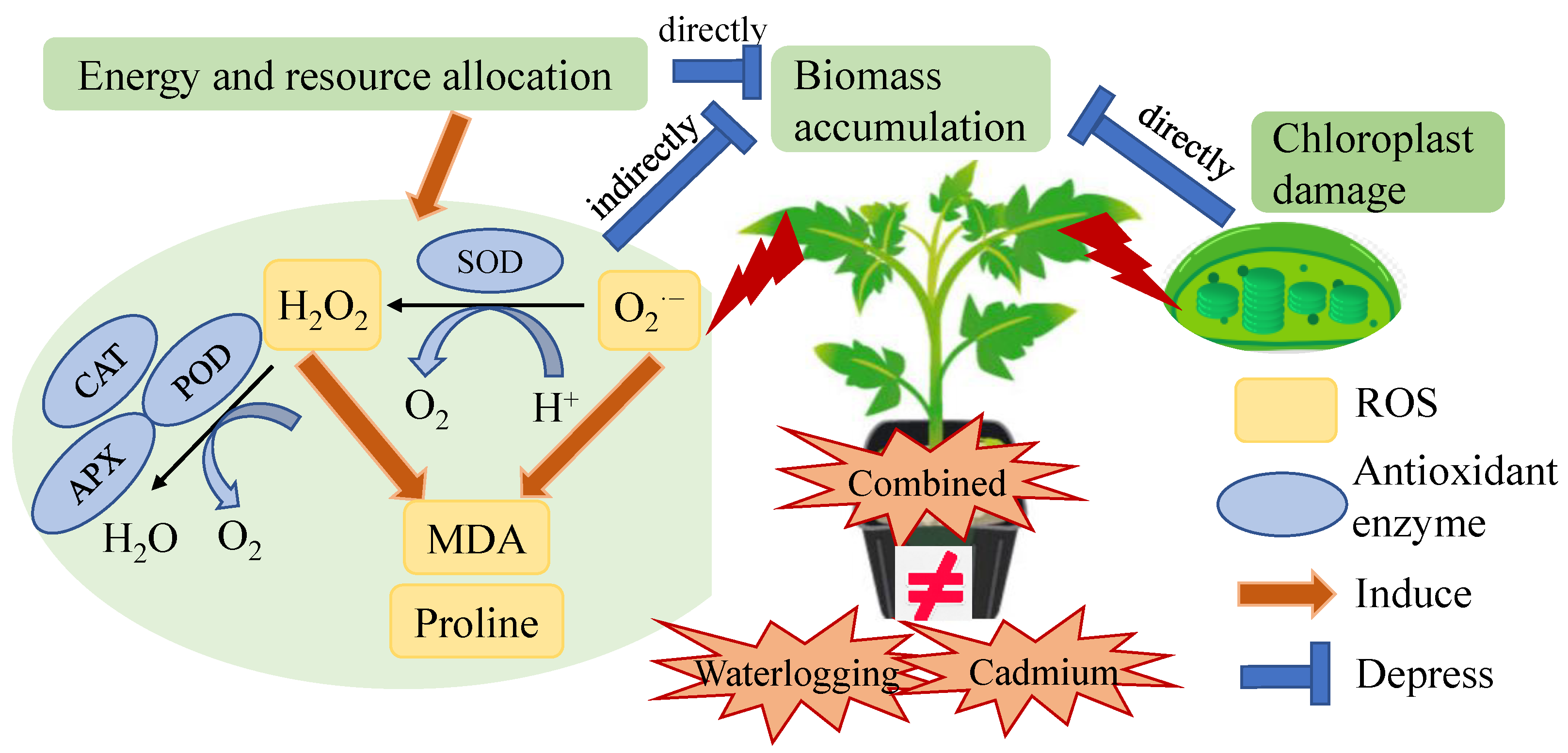
4.1. Damage of Combined Waterlogging and Cadmium Stress on Tomato Plants Was Not Accumulative
4.2. Two Tomato Genotypes under Combined Stress Exhibited Different Coordinate Regulation of Antioxidant Enzymes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wright, A.J.; de Kroon, H.; Visser, E.J.W.; Buchmann, T.; Ebeling, A.; Eisenhauer, N.; Fischer, C.; Hildebrandt, A.; Ravenek, J.M.; Roscher, C.; et al. Plants are less negatively affected by flooding when growing in species-rich plant communities. New Phytol. 2017, 213, 645–656. [Google Scholar] [CrossRef] [Green Version]
- De Ollas, C.; González-Guzmán, M.; Pitarch, Z.; Matus, J.T.; Candela, H.; Rambla, J.L.; Granell, A.; Gómez-Cadenas, A.; Arbona, V. Identification of ABA-mediated genetic and metabolic responses to soil flooding in tomato (Solanum lycopersicum L. Mill). Front. Plant Sci. 2021, 12, 613059. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Sharif, R.; Xu, X.; Chen, X. Mechanisms of waterlogging tolerance in plants: Research progress and prospects. Front. Plant Sci. 2021, 11, 627331. [Google Scholar] [CrossRef]
- Haider, F.U.; Liqun, C.; Coulter, J.A.; Cheema, S.A.; Wu, J.; Zhang, R.; Wenjun, M.; Farooq, M. Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicol. Environ. Saf. 2021, 211, 111887. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S.; Aarts, M.G.M.; Thomine, S.; Verbruggen, N. Plant science: The key to preventing slow cadmium poisoning. Trends Plant Sci. 2013, 18, 92–99. [Google Scholar] [CrossRef] [PubMed]
- El Rasafi, T.; Oukarroum, A.; Haddioui, A.; Song, H.; Kwon, E.E.; Bolan, N.S.; Tack, F.M.G.; Sebastian, A.; Prasad, M.N.V.; Rinklebe, J. Cadmium stress in plants: A critical review of the effects, mechanisms, and tolerance strategies. Crit. Rev. Environ. Sci. Technol. 2022, 52, 675–726. [Google Scholar] [CrossRef]
- Kołton, A.; Kęska, K.; Czernicka, M. Selection of tomato and cucumber accessions for waterlogging sensitivity through morpho-physiological assessment at an early vegetative stage. Agronomy 2020, 10, 1490. [Google Scholar] [CrossRef]
- Ide, R.; Ichiki, A.; Suzuki, T.; Jitsuyama, Y. Analysis of yield reduction factors in processing tomatoes under waterlogging conditions. Sci. Hortic. 2022, 295, 110840. [Google Scholar] [CrossRef]
- Piotto, F.A.; Carvalho, M.E.A.; Souza, L.A.; Rabêlo, F.H.S.; Franco, M.R.; Batagin-Piotto, K.D.; Azevedo, R.A. Estimating tomato tolerance to heavy metal toxicity: Cadmium as study case. Environ. Sci. Pollut. Res. 2018, 25, 27535–27544. [Google Scholar] [CrossRef]
- Chu, Z.; Munir, S.; Zhao, G.; Hou, J.; Du, W.; Li, N.-Y.; Lu, Y.; Yu, Q.-h.; Shabala, S.; Ouyang, B. Linking phytohormones with growth, transport activity and metabolic responses to cadmium in tomato. Plant Growth Regul. 2020, 90, 557–569. [Google Scholar] [CrossRef]
- Cheng, Y.; Li, X.; Fang, M.; Ye, Q.; Li, Z.; Ahammed, G.J. Systemic H2O2 signaling mediates epigallocatechin-3-gallate-induced cadmium tolerance in tomato. J. Hazard. Mater. 2022, 438, 129511. [Google Scholar] [CrossRef] [PubMed]
- Elkelish, A.A.; Alhaithloul, H.A.S.; Qari, S.H.; Soliman, M.H.; Hasanuzzaman, M. Pretreatment with Trichoderma harzianum alleviates waterlogging-induced growth alterations in tomato seedlings by modulating physiological, biochemical, and molecular mechanisms. Environ. Exp. Bot. 2020, 171, 103946. [Google Scholar] [CrossRef]
- Gil, C.; Boluda, R.; Ramos, J. Determination and evaluation of cadmium, lead and nickel in greenhouse soils of Almería (Spain). Chemosphere 2004, 55, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, R.; Iqbal, M.; Ashraf, M.A.; Hussain, I.; Shafiq, F.; Yousaf, A.; Zaheer, A. Glycine betaine counteracts the inhibitory effects of waterlogging on growth, photosynthetic pigments, oxidative defence system, nutrient composition, and fruit quality in tomato. J. Hortic. Sci. Biotechnol. 2018, 93, 385–391. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Nadarajah, K.K. ROS homeostasis in abiotic stress tolerance in plants. Int. J. Mol. Sci. 2020, 21, 5208. [Google Scholar] [CrossRef]
- Barickman, T.C.; Simpson, C.; Sams, C.E. Waterlogging causes early modification in the physiological performance, carotenoids, chlorophylls, proline, and soluble sugars of cucumber plants. Plants 2019, 8, 160. [Google Scholar] [CrossRef] [Green Version]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Yu, X.; Song, X.; Rosenqvist, E.; Wan, H.; Ottosen, C.O. Salinity, waterlogging and elevated [CO2] induced interactive and complicated responses in cultivated and wild tomato. J. Exp. Bot. 2022, 73, 5252–5263. [Google Scholar] [CrossRef]
- Ondrasek, G.; Romić, D.; Tanaskovik, V.; Savić, R.; Rathod, S.; Horvatinec, J.; Rengel, Z. Humates mitigate Cd uptake in the absence of NaCl salinity, but combined application of humates and NaCl enhances Cd mobility & phyto-accumulation. Sci. Total Environ. 2022, 847, 157649. [Google Scholar]
- Kumar, G.N.M.; Knowles, N.R. Changes in lipid peroxidation and lipolytic and free-radical scavenging enzyme activities during aging and sprouting of potato (Solanum tuberosum) seed-tubers. Plant Physiol. 1993, 102, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarty, D.; Datta, S.K. Micropropagation of gerbera: Lipid peroxidation and antioxidant enzyme activities during acclimatization process. Acta Physiol. Plant. 2008, 30, 325–331. [Google Scholar] [CrossRef]
- Ábrahám, E.; Hourton-Cabassa, C.; Erdei, L.; Szabados, L. Methods for determination of proline in plants. In Plant Stress Tolerance: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2010; pp. 317–331. [Google Scholar]
- Ke, D.; Sun, G.; Wang, Z. Effects of superoxide radicals on ACC synthase activity in chilling-stressed etiolated mungbean seedlings. Plant Growth Regul. 2007, 51, 83–91. [Google Scholar] [CrossRef]
- Zhou, W.; Zhao, D.; Lin, X. Effects of waterlogging on nitrogen accumulation and alleviation of waterlogging damage by application of nitrogen fertilizer and mixtalol in winter rape (Brassica napus L.). J. Plant Growth Regul. 1997, 16, 47–53. [Google Scholar] [CrossRef]
- Munoz-Munoz, J.L.; García-Molina, F.; García-Ruíz, P.A.; Arribas, E.; Tudela, J.; García-Cánovas, F.; Rodríguez-López, J.N. Enzymatic and chemical oxidation of trihydroxylated phenols. Food Chem. 2009, 113, 435–444. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Method. Enzymol. 1984, 105, 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Chen, Y.; Wang, H.; Hu, W.; Wang, S.; Wang, Y.; Snider, J.L.; Zhou, Z. Combined elevated temperature and soil waterlogging stresses inhibit cell elongation by altering osmolyte composition of the developing cotton (Gossypium hirsutum L.) fiber. Plant Sci. 2017, 256, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, U.; Hussain, S.; Hussain, S.; Ali, B.; Ashraf, U.; Zamir, S.i.; Al-Robai, S.A.; Alzahrani, F.O.; Hano, C.; El-Esawi, M.A. Morpho-physio-biochemical and molecular responses of maize hybrids to salinity and waterlogging during stress and recovery phase. Plants 2021, 10, 1345. [Google Scholar] [CrossRef]
- Altaf, A.; Zhu, X.; Zhu, M.; Quan, M.; Irshad, S.; Xu, D.; Aleem, M.; Zhang, X.; Gull, S.; Li, F.; et al. Effects of environmental stresses (heat, salt, waterlogging) on grain yield and associated traits of wheat under application of sulfur-coated urea. Agronomy 2021, 11, 2340. [Google Scholar] [CrossRef]
- Sousa, B.; Rodrigues, F.; Soares, C.; Martins, M.; Azenha, M.A.; Lino-Neto, T.; Santos, C.; Cunha, A.; Fidalgo, F. Impact of combined heat and salt stresses on tomato plants—Insights into nutrient uptake and redox homeostasis. Antioxidants 2022, 11, 478. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Yu, X.; Zhao, T.; Ottosen, C.O.; Rosenqvist, E.; Wu, Z. Physiological analysis and transcriptome sequencing reveal the effects of combined cold and drought on tomato leaf. BMC Plant Biol. 2019, 19, 377. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Kong, L.; Yu, X.; Ottosen, C.O.; Zhao, T.; Jiang, F.; Wu, Z. Oxidative damage and antioxidant mechanism in tomatoes responding to drought and heat stress. Acta Physiol. Plant. 2019, 41, 20. [Google Scholar] [CrossRef]
- Lopes, N.G.; Kloss, R.B.; Dos Santos, I.C.; Souza, V.L.; Prasad, M.N.; Mangabeira, P.A.; França, M.G. Soil flooding and its outcome on cadmium and nutrient uptake affect photosynthetic activity in Inga laurina plants. Ecotoxicology 2023, 32, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Dong, F.; Yang, Y.; Zeng, Q.; Chen, B.; Jiang, L. Effects of waterlogging and cadmium on ecophysiological responses and metal bio-accumulation in Bermuda grass (Cynodon dactylon). Environ. Earth Sci. 2017, 76, 719. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Răcuciu, M.; Tecucianu, A.; Oancea, S. Impact of magnetite nanoparticles coated with aspartic acid on the growth, antioxidant enzymes activity and chlorophyll content of maize. Antioxidants 2022, 11, 1193. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, X.D.; Li, T.L.; Cao, X. Photosynthesis and ultrastructure of photosynthetic apparatus in tomato leaves under elevated temperature. Photosynthetica 2014, 52, 430–436. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Main Factors | Interactions | |||||
|---|---|---|---|---|---|---|---|
| Cultivar | WL | Cd | Cultivar × WL | Cultivar × Cd | WL × Cd | Cultivar × WL × Cd | |
| Plant height | ** | ** | ** | ns | ns | * | ns |
| Stem diameter | ** | ** | ns | * | ns | ns | ns |
| Fresh weight of shoot | * | ** | ** | ns | ns | * | ns |
| Dry weight of shoot | * | ** | ** | ns | ns | ns | ns |
| Fresh weight of root | ns | ** | ns | * | ns | ** | ns |
| Dry weight of root | ns | ** | ns | ns | ns | ** | ns |
| MDA content | ** | * | ** | ** | ** | ns | ns |
| H2O2 content | ** | ** | ns | ns | ns | ns | ns |
| Proline content | ** | ns | * | ns | ns | ns | ns |
| O2·− production rate | * | ns | ns | ns | ns | ns | ** |
| SOD activity | ** | ns | ** | ** | ns | * | ns |
| CAT activity | ns | ns | ns | ns | ** | * | ** |
| POD activity | ** | ns | ns | * | ns | ns | ns |
| APX activity | ** | * | ns | ** | ** | ns | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, R.; Niu, L.; Yin, J.; Jiang, F.; Wang, Y.; Zhao, T.; Wu, Z.; Zhu, W. Differences in Physiological Responses of Two Tomato Genotypes to Combined Waterlogging and Cadmium Stresses. Antioxidants 2023, 12, 1205. https://doi.org/10.3390/antiox12061205
Zhou R, Niu L, Yin J, Jiang F, Wang Y, Zhao T, Wu Z, Zhu W. Differences in Physiological Responses of Two Tomato Genotypes to Combined Waterlogging and Cadmium Stresses. Antioxidants. 2023; 12(6):1205. https://doi.org/10.3390/antiox12061205
Chicago/Turabian StyleZhou, Rong, Lifei Niu, Jian Yin, Fangling Jiang, Yinlei Wang, Tongmin Zhao, Zhen Wu, and Weimin Zhu. 2023. "Differences in Physiological Responses of Two Tomato Genotypes to Combined Waterlogging and Cadmium Stresses" Antioxidants 12, no. 6: 1205. https://doi.org/10.3390/antiox12061205
APA StyleZhou, R., Niu, L., Yin, J., Jiang, F., Wang, Y., Zhao, T., Wu, Z., & Zhu, W. (2023). Differences in Physiological Responses of Two Tomato Genotypes to Combined Waterlogging and Cadmium Stresses. Antioxidants, 12(6), 1205. https://doi.org/10.3390/antiox12061205