

Characterization of Health Beneficial Components in Discarded Leaves of Three Escarole (Cichorium endivia L.) Cultivar and Study of Their Antioxidant and Anti-Inflammatory Activities


 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Extraction Procedure
2.3. NMR Analysis
2.3.1. Sample Preparation
2.3.2. NMR Spectroscopy and Processing
2.4. Mass Spectrometry Analyses
2.4.1. Qualitative Analyses
2.4.2. Quantitative Analyses
2.5. Antioxidant Assay
2.5.1. The DPPH (2,2′-Diphenyl-1-picrylhydrazyl Radical) Assay
2.5.2. ABTS Assay Radical Cation Decolorization Assay
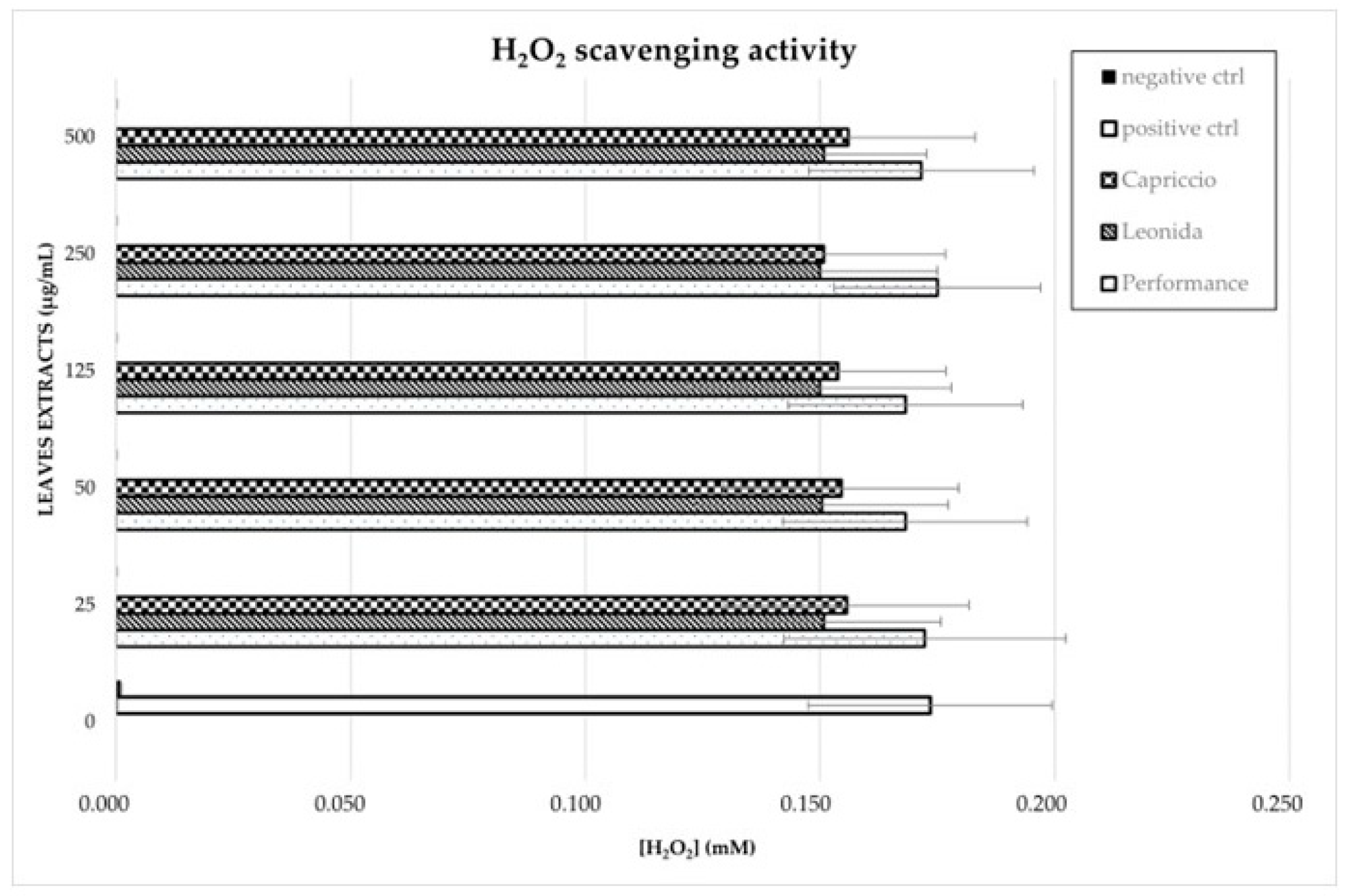
2.5.3. H2O2 Scavenging Activity Using HRP Assay
2.6. Cell Culture and Differentiation
2.7. Cell Viability Assay
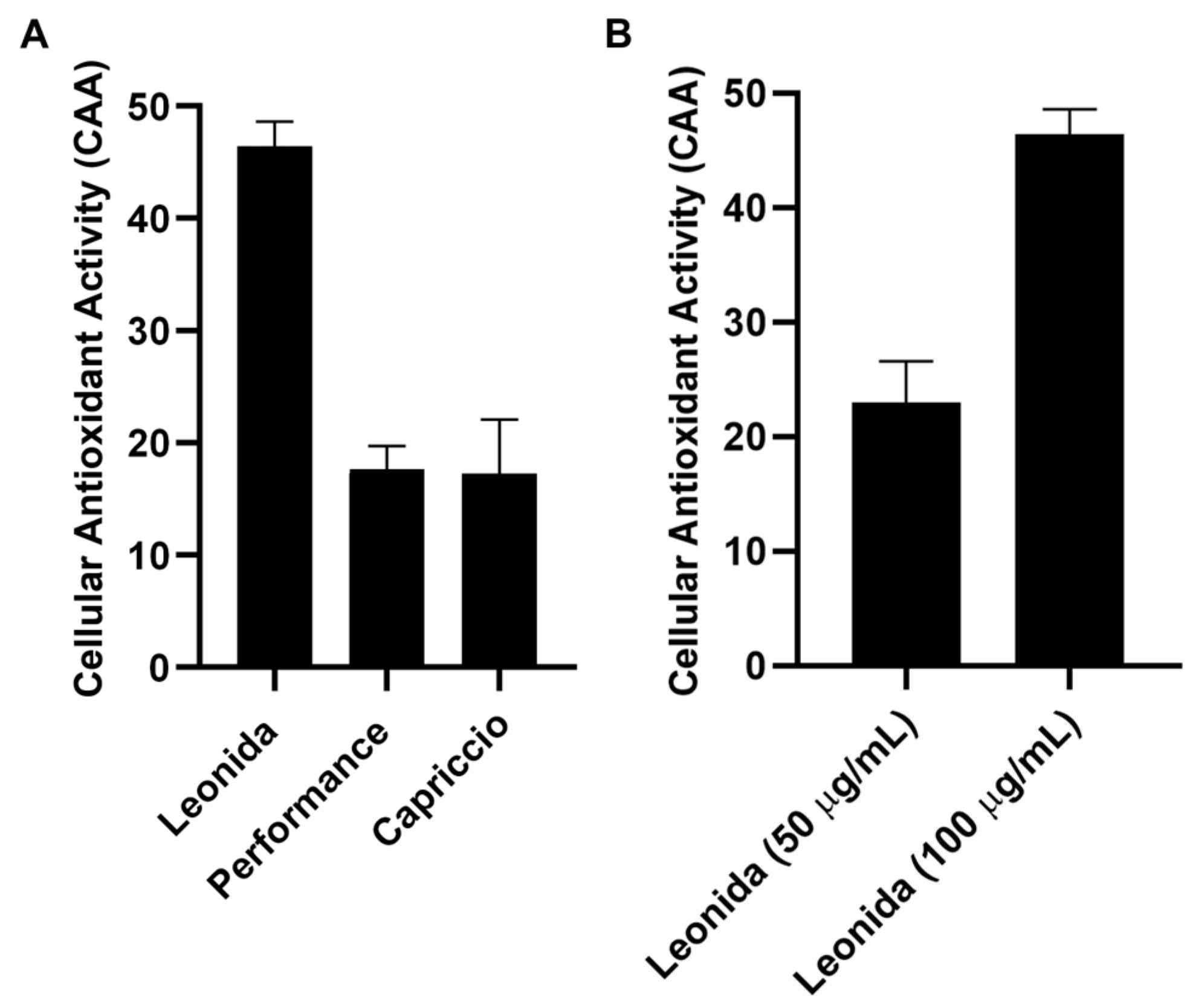
2.8. Cellular Antioxidant Activity (CAA) Evaluation on THP-1 Derived Macrophages M0
2.9. Cytokine Production and Enzyme-Linked Immunosorbent Assay (ELISA) on THP-1 Derived Macrophages M0
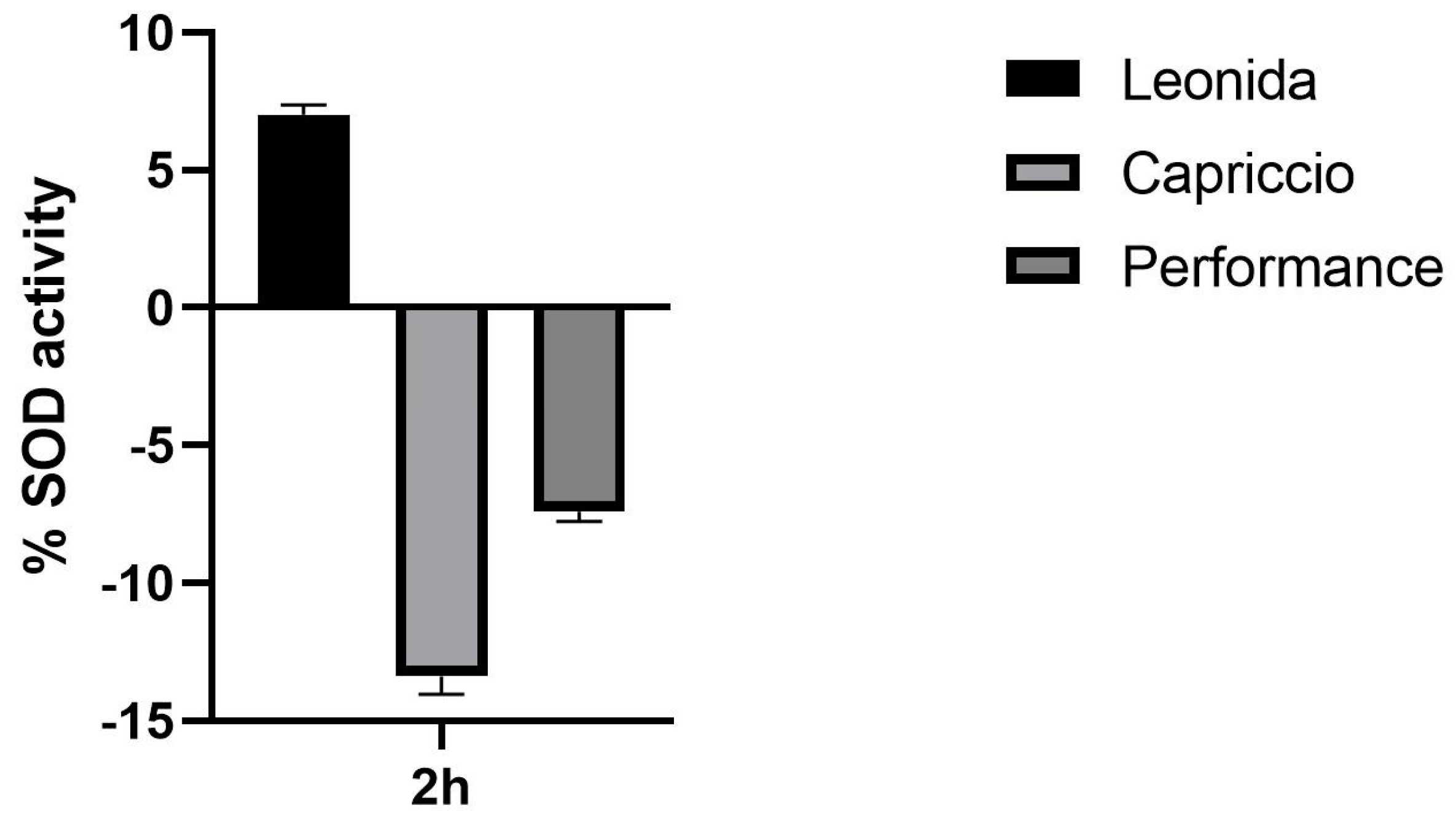
2.10. Superoxide Dismutase (SOD) Colorimetric Activity Kit
2.11. Western Blot Analysis
2.12. Catalase Activity Assay
3. Results
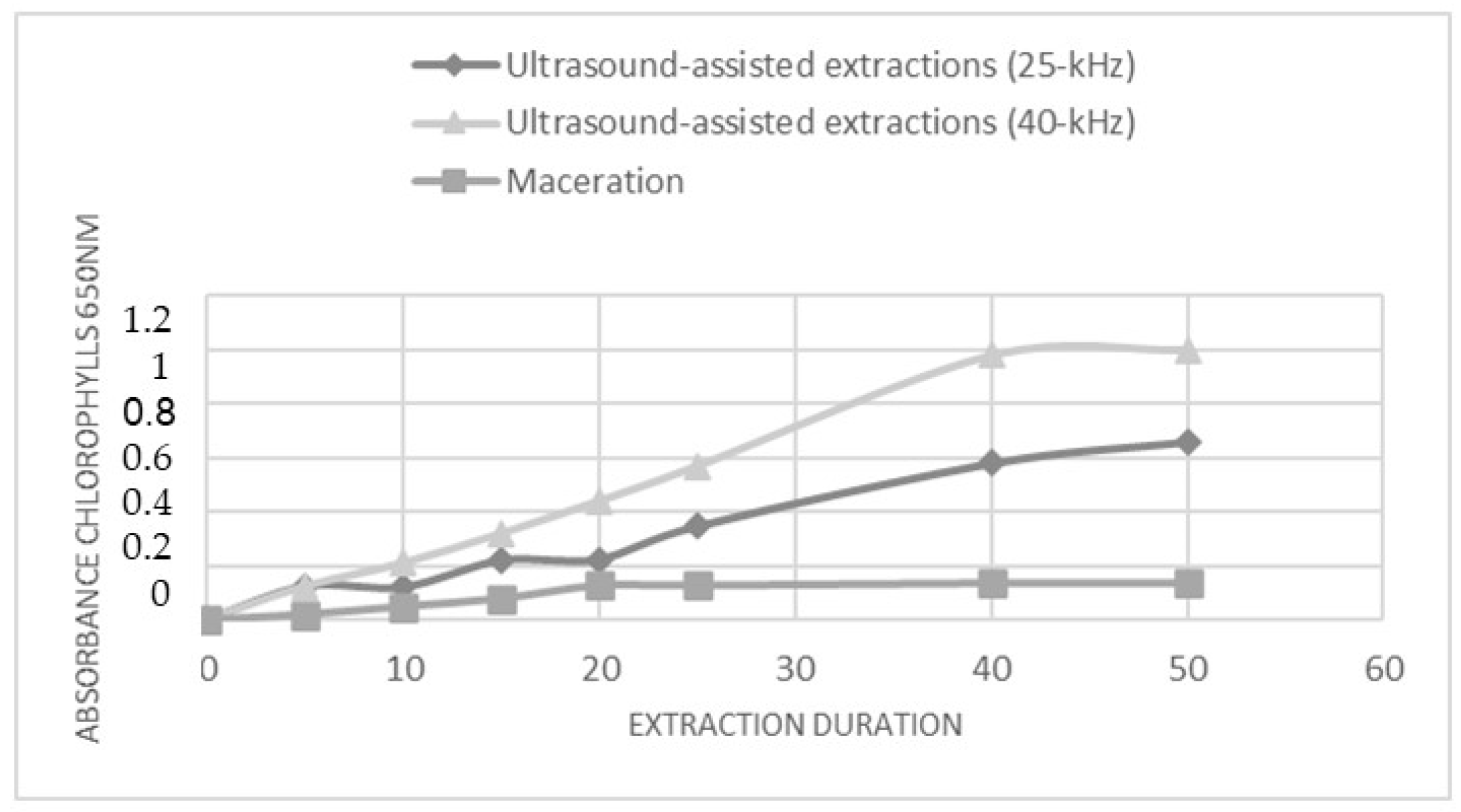
3.1. Optimized Ultrasound-Assisted Extraction Procedure
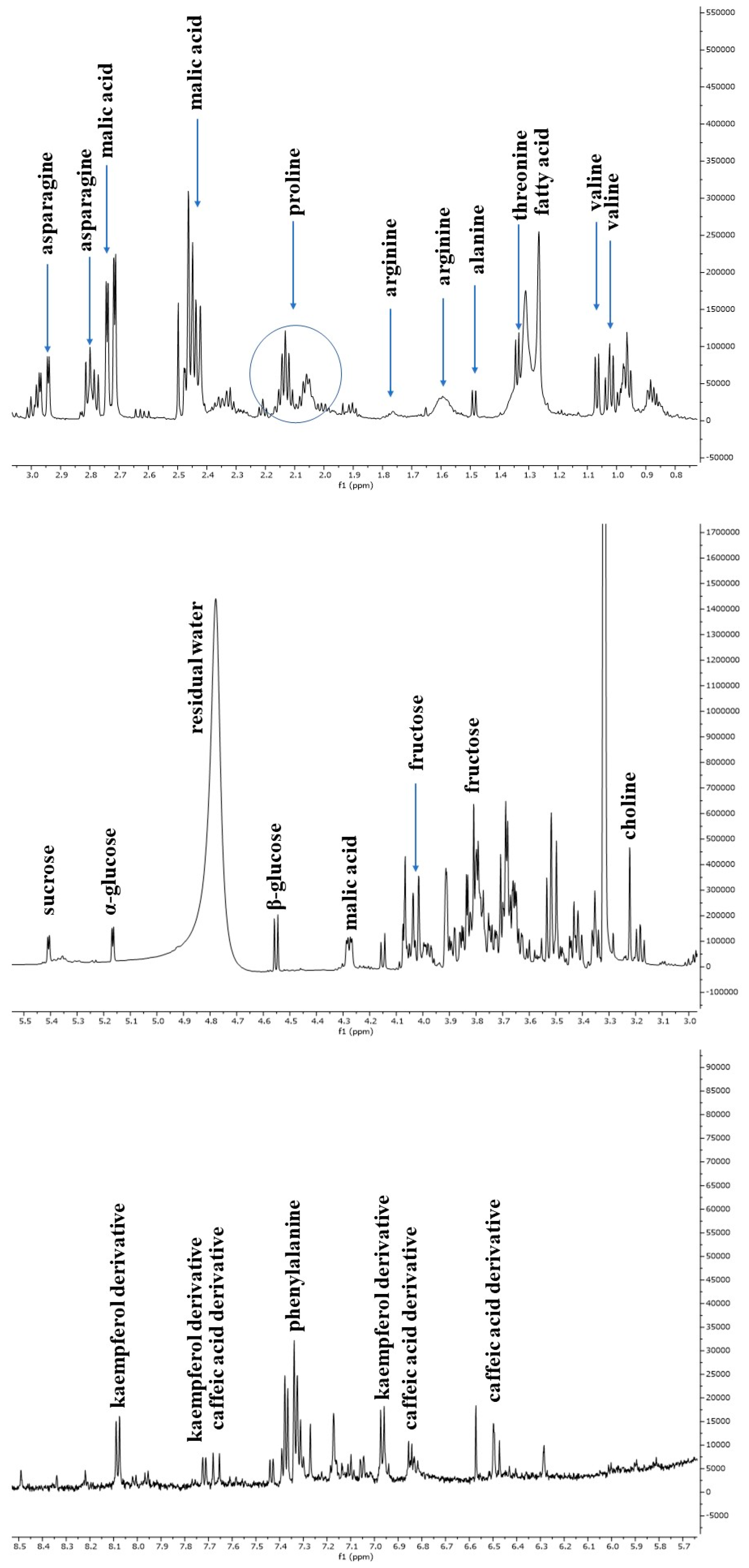
3.2. NMR-Based Metabolomic Profiling of Escarole
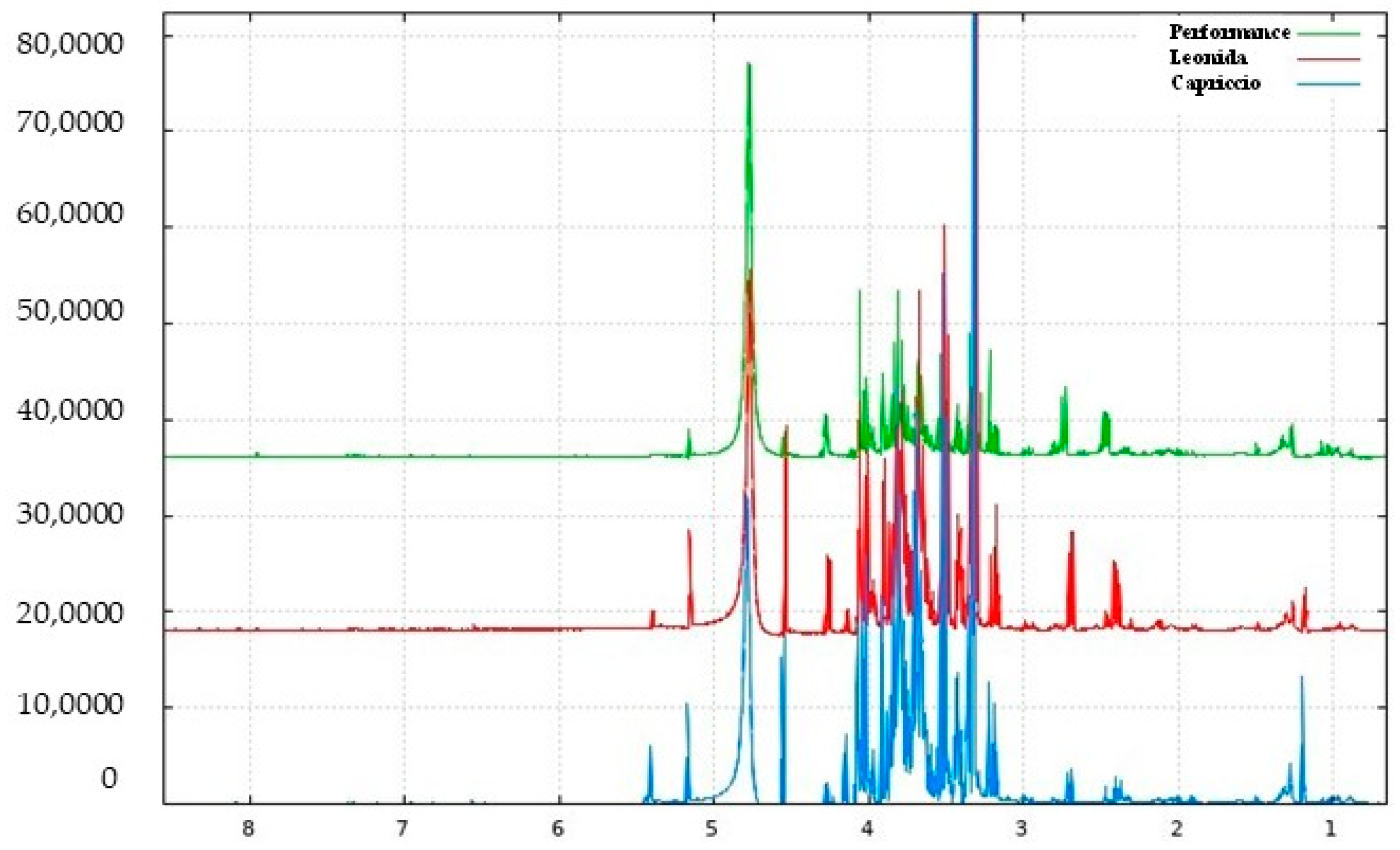
3.3. LC/MSMS Analysis
3.3.1. Qualitative Analyses
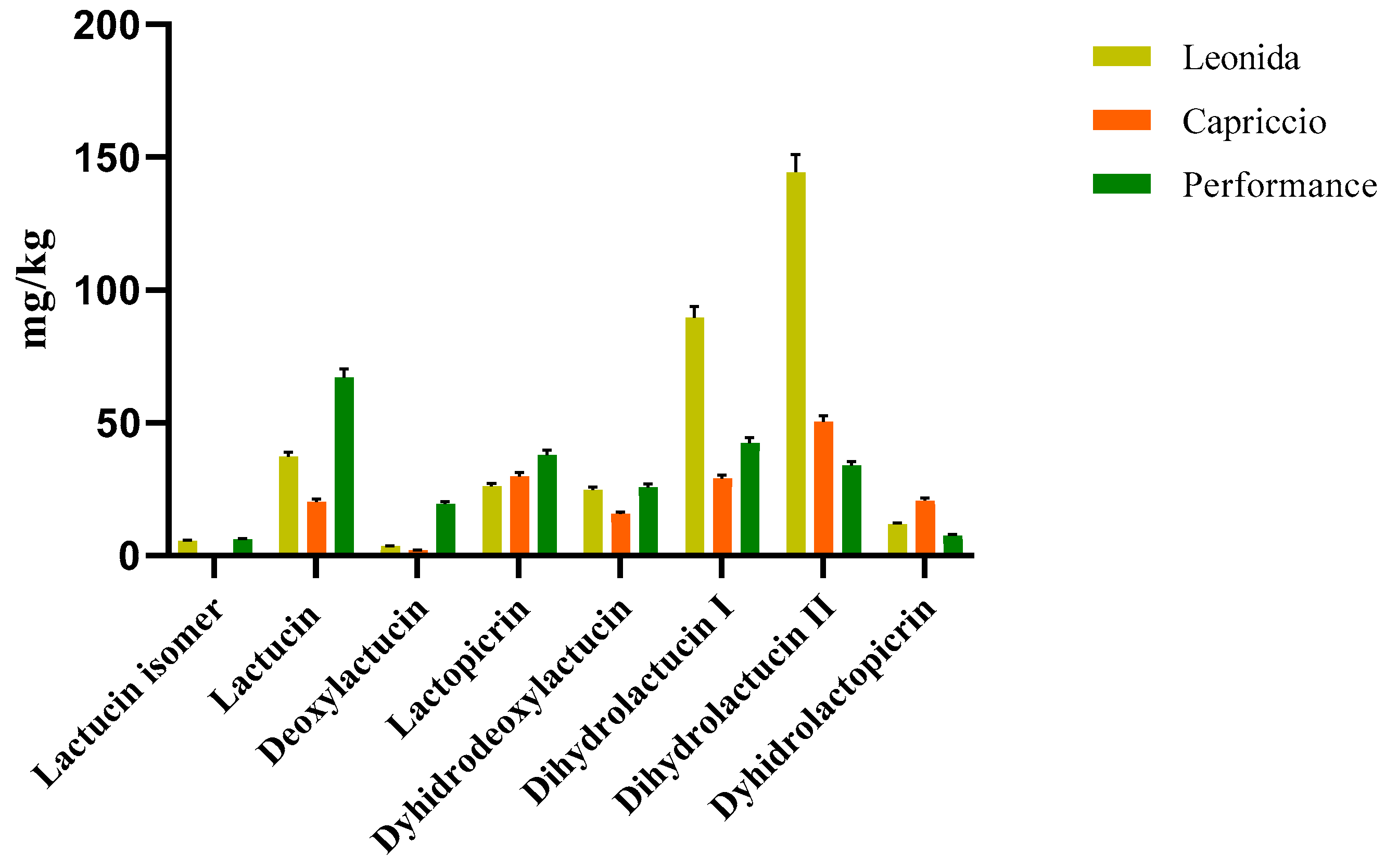
3.3.2. Quantitative Analyses
3.4. Antioxidant Activity
3.5. In Cell Antioxidant Activity
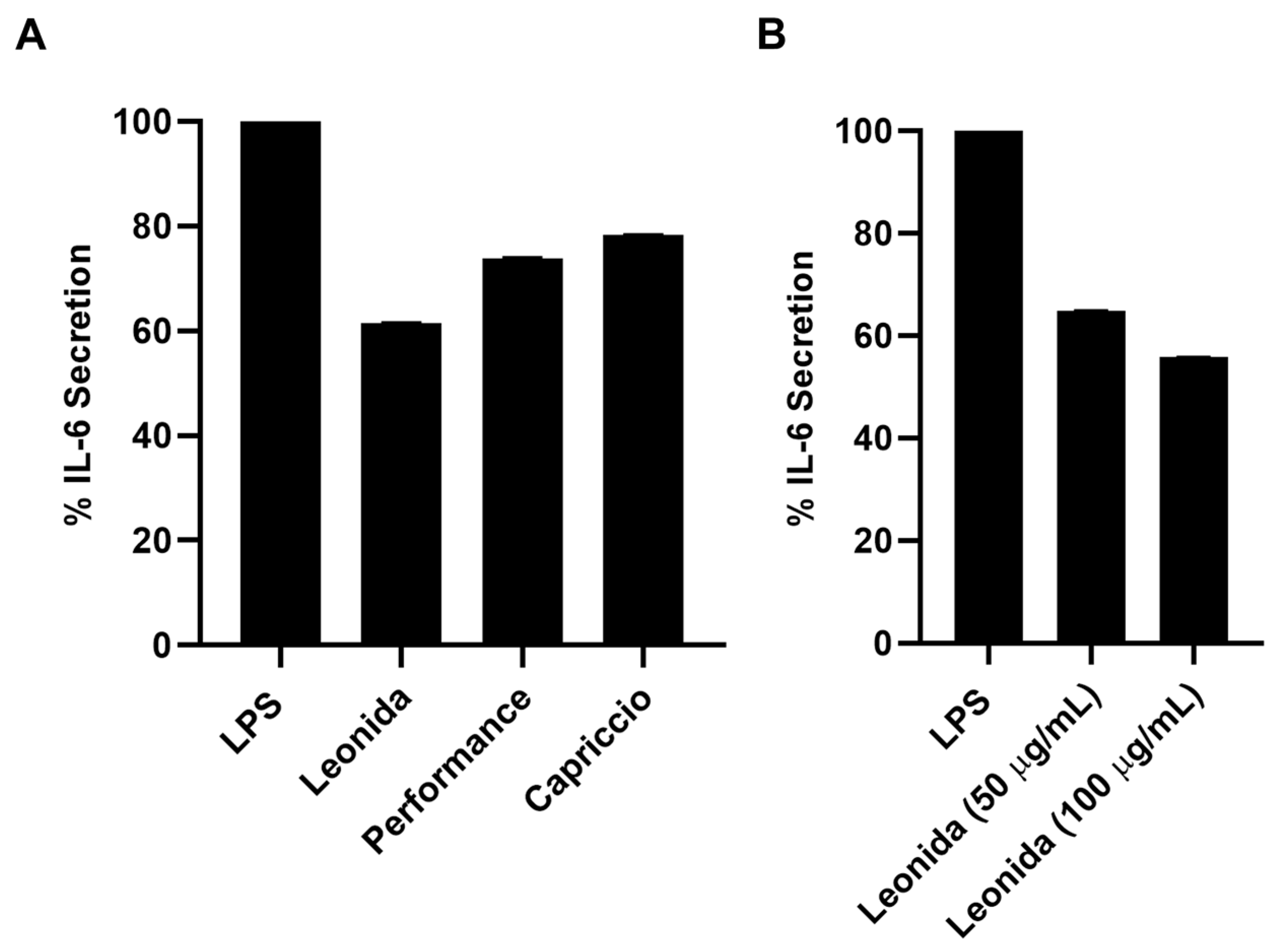
3.6. Anti-Inflammatory Activity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sareedenchai, V.; Zidorn, C. Flavonoids as chemosystematic markers in the tribe Cichorieae of the Asteraceae. Biochem. Syst. Ecol. 2015, 38, 935–957. [Google Scholar] [CrossRef]
- Hegazy, A.K.; Ezzat, S.M.; Qasem, I.B.; Ali-Shtayeh, M.S.; Basalah, M.O.; Ali, H.M.; Hatamleh, A.A. Diversity of active constituents in Cichorium endivia and Cynara cornigera extracts. Acta Biol. Hung. 2015, 66, 103–118. [Google Scholar] [CrossRef] [Green Version]
- Llorach, R.; Martinez-Sanchez, A.; Tomas-Barberan, F.A.; Gil, M.I.; Ferreres, F. Characterization of polyphenols and antioxidant properties of five lettuce varieties and escarole. Food Chem. 2008, 108, 1028–1038. [Google Scholar] [CrossRef]
- Warashina, T.; Miyase, T. Sesquiterpenes from the Roots of Cichorium endivia. Chem. Pharm. Bull. 2008, 56, 1445–1451. [Google Scholar] [CrossRef] [Green Version]
- Molan, A.L.; Duncan, A.J.; Barryand, T.N.; McNabb, W.C. Effect of condensed tannins and sesquiterpene lactones extracted from chicory on the motility of larvae of deer lungworm and gastrointestinal nematodes. Parasitol. Int. 2003, 52, 209–218. [Google Scholar] [CrossRef]
- Muthusamy, V.S.; Anand, S.; Sangeetha, K.N.; Sujatha, S.; Arun, B.; Lakshami, B.S. Tannins present in Cichorium intybus enhance glucose uptake and inhibit adipogenesis in 3T3-L1 adipocytes through PTP1B inhibition. Chem. Biol. Interact. 2008, 174, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Innocenti, M.; Gallori, S.; Giaccherini, C.; Ieri, F.; Vincieri, F.F.; Mulinacci, N. Evaluation of phenolic content in the aerial parts different varieties of Cichorium intybus L. J. Agric. Food Chem. 2005, 53, 6497–6502. [Google Scholar] [CrossRef] [PubMed]
- Perović, J.; Šaponjac, V.T.; Kojić, J.; Krulj, J.; Moreno, D.A.; García-Viguera, C.; Bodroža-Solarov, M.; Ilić, N. Chicory (Cichorium intybus L.) as a food ingredient–Nutritional composition, bioactivity, safety, and health claims: A review. Food Chem. 2021, 336, 127676. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Contreras, M.M.; Arráez-Román, D.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Reversed-phase ultra-high- liquid chromatography coupled to electrospray ionization-quadrupole-time-of-flight mass spectrometry as a powerful tool for metabolic profiling of vegetables: Lactuca sativa as an example of its application. J. Chromatogr. A 2013, 1313, 212–227. [Google Scholar] [CrossRef]
- Atta, A.H.; Elkoly, T.A.; Mouneir, S.M.; Kamel, G.; Alwabel, N.A.; Zaher, S. Hepatoprotective effect of methanolic extracts of Zingiber officinale and Cichorium intybus. Indian J. Pharm. Sci. 2010, 72, 564–570. [Google Scholar] [CrossRef] [Green Version]
- Lucchin, M.; Varotto, S.; Barcaccia, G.; Parrini, P. Chicory and Endive. In Vegetables I: Asteraceae, Brassicaceae Chenopodi-Caceae; Prohens, J., Nuez, F., Eds.; Handbook of Plant Breeding; Springer: New York, NY, USA, 2008; Volume 1, pp. 3–48. [Google Scholar]
- Testone, G.; Mele, G.; Di Giacomo, E.; Tenore, G.C.; Gonnella, M.; Nicolodi, C.; Frugis, G.; Iannelli, M.A.; Arnesi, G.; Schiappa, A.; et al. Transcriptome driven characterization of curly- and smooth-leafed endives reveals molecular differences in the sesquiterpenoid pathway. Hortic. Res. 2019, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Wesolowska, A.; Nikiforuk, A.; Michalska, K.; Kisiel, W.; Chojnacka-Wojcik, E. Analgesic and sedative activities of lactucin and some lactucin-like guaianolides in mice. J. Ethnopharmacol. 2006, 107, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.S.; Anastácio, J.D.; Nunes dos Santos, C. Sesquiterpene lactones: Promising natural compounds to fight inflammation. Pharmaceutics 2021, 13, 991. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Thompson, S.; Turchini, G.M. Organic aquaculture productivity, environmental sustainability, and food security: Insights from organic agriculture. Food Secur. 2020, 12, 1253–1267. [Google Scholar] [CrossRef]
- Vertakova, Y.V.; Plotnikov, V.A. The integrated approach to sustainable development: The case of energy efficiency and solid waste management. Int. J. Energy Econ. Policy 2019, 9, 194–201. [Google Scholar] [CrossRef]
- Nariya, P.B.; Bhalodia, N.R.; Shukla, V.J.; Acharya, R.; Nariya, M.B. In vitro evaluation of antioxidant activity of Cordia dichotoma (Forst f.) bark. Ayu 2013, 34, 124–128. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assay. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Cellular Antioxidant Activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, C.; Wang, B.; Yagoub, A.E.G.A.; Ma, H.; Zhang, X.; Wu, M. Study of ultrasonic cavitation during extraction of the peanut oil at varying frequencies. Ultrason. Sonochem. 2017, 37, 106–113. [Google Scholar] [CrossRef]
- Sobolev, A.P.; Brosio, E.; Gianferri, R.; Segre, A.L. Metabolic profile of lettuce leaves by high-field NMR spectra. Magn. Reson. Chem. 2005, 43, 625–638. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The Human Metabolome Database. Nucleic Acids Res. 2007, 35, D521–D526. [Google Scholar] [CrossRef] [PubMed]
- Markley, J.L.; Anderson, M.E.; Cui, Q.; Eghbalnia, H.R.; Lewis, I.A.; Hegeman, A.D.; Li, J.; Schulte, C.F.; Sussman, M.R.; Westler, W.M.; et al. New bioinformatics resources for metabolomics. Pac. Symp. Biocomput. 2007, 12, 157–168. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Graziani, G.; Ferracane, R.; Sambo, P.; Santagata, S.; Nicoletto, C.; Fogliano, V. Profiling chicory sesquiterpene lactones by high resolution mass spectrometry. Food Res. Int. 2015, 67, 193–198. [Google Scholar] [CrossRef]
- Abbas, Z.K.; Saggu, S.; Sakeran, M.I.; Zidan, N.; Rehman, H.; Ansari, A.A. Phytochemical, antioxidant and mineral composition of hydroalcoholic extract of chicory (Cichorium intybus L.) leaves. Saudi J. Biol. Sci. 2015, 22, 322–326. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Shahidi, F. Methods for the assessment of antioxidant activity in foods. In Handbook of Antioxidants for Food Preservation, 1st ed.; Shadidi, F., Ed.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 287–333. [Google Scholar]
- Jiménez, I.; Sánchez-Moreno, C.; Saura-Calixto, F. Evaluation of free radical scavenging of dietary carotenoids by the stable radical 2,2-diphenyl-1-picrylhydrazyl. J. Sci. Food Agric. 2000, 80, 1686–1690. [Google Scholar] [CrossRef]
- Sokovic, M.; Ciric, A.; Glamoclija, J.; Skaltsa, H. Biological Activities of Sesquiterpene Lactones Isolated from the Genus centaurea L. (Asteraceae). Curr. Pharm. Des. 2017, 23, 2767–2786. [Google Scholar] [CrossRef] [PubMed]
- Kreuger, M.R.; Grootjans, S.; Biavatti, M.W.; Vandenabeele, P.; D’Herde, K. Sesquiterpene lactones as drugs with multiple targets in cancer treatment: Focus on parthenolide. Anti-Cancer Drugs 2012, 23, 883–896. [Google Scholar] [CrossRef] [PubMed]
- Quintana, J.; Estévez, F. Recent Advances on Cytotoxic Sesquiterpene Lactones. Curr. Pharm. Des. 2018, 24, 4355–4361. [Google Scholar] [CrossRef]
- Paço, A.; Brás, T.; Santos, J.O.; Sampaio, P.; Gomes, A.C.; Duarte, M.F. Anti-inflammatory and immunoregulatory action of sesquiterpene lactones. Molecules 2022, 27, 1142. [Google Scholar] [CrossRef]
- Crops: Areas and Production—Overall Data—Provinces. Available online: http://dati.istat.it/Index.aspx?QueryId=37850# (accessed on 13 May 2023).
- Arancon, R.A.D.; Lin, C.S.K.; Chan, K.M.; Kwan, T.H.; Luque, R. Advances on waste valorization: New horizons for a more sustainable society. Energy Sci. Eng. 2013, 1, 53–71. [Google Scholar] [CrossRef]
- Stefaniu, A.; Pirvu, L.C. In Silico Study Approach on a Series of 50 Polyphenolic Compounds in Plants; A Comparison on the Bioavailability and Bioactivity Data. Molecules 2022, 27, 1413. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, J.; Wu, Y.; Li, Y.; Guo, F. Guaiane-type sesquiterpenes from Curcumawenyujin. Phytochemistry 2022, 198, 113164. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
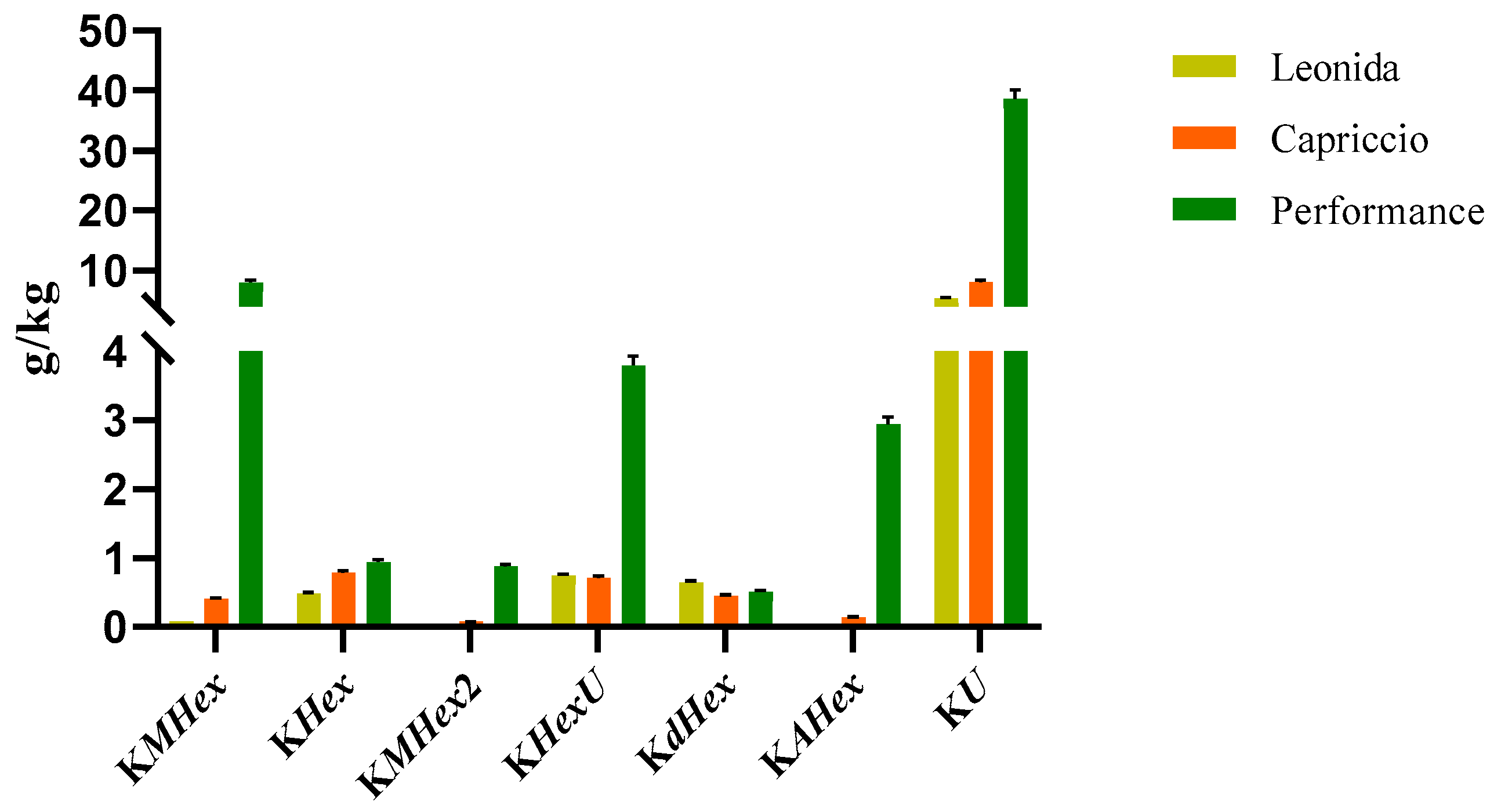{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Chemical Shift (ppm) Multiplicity (J in Hz) | MSI Statusa | Identification Confirmation |
|---|---|---|---|
| Valine | 1.07 d (J = 7.2); 1.02 d (J = 7.2) | 2 | 1H-13C HSQC |
| Threonine | 1.34 d (J = 6.5) | 2 | 1H-13C HSQC |
| Alanine | 1.49 d (J = 7.30) | 2 | 1H-13C HSQC |
| Malic Acid | 2.46 dd, (J = 15.6, 8.3); 2.74 dd (J = 16.0, 3.5) | 2 | 1H-13C HSQC |
| Arginine | 1.59 m; 1.77 m | 2 | 1H-13C HSQC |
| Asparagine | 2.95 dd (J = 14.7, 3.9); 2.78 dd (J = 16.8, 9.0) | 2 | 1H-13C HSQC |
| Choline | 3.2 s | 2 | 1H-13C HSQC |
| α-Glucose | 5.16 d (J = 4.40) | 1 | 1H-13C HSQC, Spike |
| β-Glucose | 4.54 d (J = 7.9) | 1 | 1H-13C HSQC Spike |
| Sucrose | 5.40 d (J = 3.8) | 1 | 1H-13C HSQC, Spike |
| Fructose | 4.02 dd (J = 12.3, 1.3) | 2 | 1H-13C HSQC |
| α-Galactose | 5.2 d (J = 3.4) | 2 | 1H-13C HSQC |
| Phenylalanine | 7.37 m | 2 | 1H-13C HSQC |
| Fatty acid | 1.27 s | 3 | 1H-13C HSQC |
| Caffeic acid derivative | 7.70 d (J = 16.0); 6.48 d (J = 16.0) | 3 | 1H-13C HSQC |
| Kaempferol derivative | 8.08 d (J = 8.7); 6.97 d (J = 8.7) | 3 | 1H-13C HSQC |
| Pek | Rt (Min) | [M+H]+ | [M-H]− | Fragment Ions | Compound Identity | MSI Status a |
|---|---|---|---|---|---|---|
| 1 | 11.66 | 515.1406 | 353 [M-162]− 191 [M-162-162]− | Caffeoylquinic acid-O-hexoside | 2 | |
| 2 | 12.68 | 277.1070 | 241 [M-36]+ 213 [M-64]+ 185 [M-92]+ | Lactucin isomer | 2 | |
| 3 | 13.62 | 341.0867 | 179 [M-162]+ | Aesculin | 1 | |
| 4 | 15.69 | 355.1023 | 163 [M-192]+ | Chlorogenic acid | 2 | |
| 5 | 16.04 | 279.1227 | 243 [M-36]+ 215 [M-64]+ 187 [M-92]+ | Dihydrolactucin | 2 | |
| 6 | 17.17 | 611.1609 | 287 [M-324]+ | Kaempferol-O-dihexoside | 2 | |
| 7 | 17.26 | 625.1401 | 287 [M-338]+ | Kaempferol-O-hexoside-uronide | 2 | |
| 8 | 17.56 | 389.2177 | 371 [M-18]+ | Dihydroroseoside | 2 | |
| 9 | 17.65 | 279.1227 | 243 [M-36]+ 215 [M-64]+ 187 [M-92]+ | Dihydrolactucin isomer | 2 | |
| 10 | 17.77 | 387.2015 | 369 [M-18]+ | Roseoside | 1 | |
| 11 | 18.35 | 697.1608 | 287 [M-410]+ | Kaempferol-O-malonyldihexoside | 2 | |
| 12 | 18.68 | 277.1070 | 241 [M-36]+ 213 [M-64]+ 185 [M-92]+ | Lactucin | 1 | |
| 13 | 19.19 | 367.1023 | 191 [M-176]− 173 [M-176-18]− | Feruloylquinic acid | 2 | |
| 14 | 19.46 | 639.1555 | 477 [M-162]− 315 [M-162-162]− | Isorhamnetina-O-dihexoside | 2 | |
| 15 | 19.69 | 337.0935 | 191 [M-196]− | Coumaroylquinic acid | 2 | |
| 16 | 21.86 | 479.0820 | 303 [M-176]+ | Quercetin-O-uronide | 2 | |
| 17 | 22.12 | 223.1328 | 205 [M-18]+ 187 [M-36]+ 105 [M-118]+ | Dihydro-vomifoliol | 2 | |
| 18 | 22.64 | 473.0733 | 293 [M-180]− | Chicoric acid | 2 | |
| 19 | 22.68 | 197.1172 | 179 [M-26]+ 161 [M-36]+ 135 [M-62]+ | Loliolide | 1 | |
| 20 | 23.02 | 263.1277 | 245 [M-18]+ 217 [M-46]+ 189 [M-74]+ | Dihydro-deoxylactucin | 2 | |
| 21 | 23.14 | 517.1346 | 163 [M-354]+ | Dicaffeoylquinic acid | 2 | |
| 22 | 23.21 | 449.1078 | 287 [M-162]+ | Kaempferol-O-hexoside | 2 | |
| 23 | 23.33 | 463.0869 | 287 [M-176]+ | Kaempferol-O-uronide | 2 | |
| 24 | 23.95 | 225.1488 | 207 [M-18]+ | Vomifoliol | 2 | |
| 25 | 24.76 | 535.1069 | 287 [M-248]+ | Kaempferol-O malonylhexoside | 2 | |
| 26 | 25.08 | 489.1049 | 285 [M-204]− | Kaempferol-O-acetylhexoside | 2 | |
| 27 | 30.12 | 261.1121 | 243 [M-18]+ 215 [M-46]+ 187 [M-74]+ | Deoxyilactucin | 2 | |
| 28 | 30.13 | 413.1598 | 261 [M-152]− 215 [M-198]− | Dihydrolactucopicrin | 2 | |
| 29 | 30.19 | 411.1442 | 259 [M-152]− 213 [M-198]− 185 [M-226]− | Lactucopicrin | 2 | |
| 30 | 31.14 | 327.2181 | 309 [M-18]− 291 [M-18]− 229 [M-98]− 211 [M-98-18]− | Trihydroxy-octadecadienoic acid | 2 | |
| 31 | 32.87 | 329.2339 | 311 [M-18]− 293 [M-18]− | Trihydroxy-octadecadienoic acid | 2 | |
| 32 | 46 | 293.2124 | 275 [M-18]− 231 [M-18-44]− | Hydroxy-octadecatrienoic acid | 2 | |
| 33 | 46.21 | 293.2124 | 275 [M-18]− 231 [M-18-44]− | Hydroxy-octadecatrienoic acid isomer | 2 | |
| 34 | 48.67 | 277.2173 | 259 [M-18]− | Octadecatrienoic acid | 2 | |
| 35 | 49.85 | 279.2328 | 261 [M-18]− | Octadecadienoic acid | 2 |
| Samples | DPPH TEAC mM | ABTS TEAC mM |
|---|---|---|
| C. endivia var latifolium. ‘Performance’ | 2.51 ± 0.25 | 3.49 ± 0.28 |
| C. endivia var latifolium ‘Leonida’ | 0.60 ± 0.04 | 0.78 ± 0.07 |
| C. endivia var crispum. ‘Capriccio’ | 0.95 ± 0.05 | 1.14 ± 0.22 |
| Samples | Abs 420 nm | H2O2 Concentration (mM) |
|---|---|---|
| Negative control (without catalase) | 2.200 | 0.625 ± 0.1 |
| Positive control (30 U catalase) | 0.002 | 0.001 |
| C. endivia var latifolium. ‘Performance’ | 0.003 | 0.001 |
| C. endivia var latifolium ‘Leonida‘ | 0.002 | 0.001 |
| C. endivia var crispum. ‘Capriccio’ | 0.002 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donadio, G.; Bellone, M.L.; Mensitieri, F.; Parisi, V.; Santoro, V.; Vitiello, M.; Dal Piaz, F.; De Tommasi, N. Characterization of Health Beneficial Components in Discarded Leaves of Three Escarole (Cichorium endivia L.) Cultivar and Study of Their Antioxidant and Anti-Inflammatory Activities. Antioxidants 2023, 12, 1402. https://doi.org/10.3390/antiox12071402
Donadio G, Bellone ML, Mensitieri F, Parisi V, Santoro V, Vitiello M, Dal Piaz F, De Tommasi N. Characterization of Health Beneficial Components in Discarded Leaves of Three Escarole (Cichorium endivia L.) Cultivar and Study of Their Antioxidant and Anti-Inflammatory Activities. Antioxidants. 2023; 12(7):1402. https://doi.org/10.3390/antiox12071402
Chicago/Turabian StyleDonadio, Giuliana, Maria Laura Bellone, Francesca Mensitieri, Valentina Parisi, Valentina Santoro, Maria Vitiello, Fabrizio Dal Piaz, and Nunziatina De Tommasi. 2023. "Characterization of Health Beneficial Components in Discarded Leaves of Three Escarole (Cichorium endivia L.) Cultivar and Study of Their Antioxidant and Anti-Inflammatory Activities" Antioxidants 12, no. 7: 1402. https://doi.org/10.3390/antiox12071402
APA StyleDonadio, G., Bellone, M. L., Mensitieri, F., Parisi, V., Santoro, V., Vitiello, M., Dal Piaz, F., & De Tommasi, N. (2023). Characterization of Health Beneficial Components in Discarded Leaves of Three Escarole (Cichorium endivia L.) Cultivar and Study of Their Antioxidant and Anti-Inflammatory Activities. Antioxidants, 12(7), 1402. https://doi.org/10.3390/antiox12071402