


Physical Activity as a Modern Intervention in the Fight against Obesity-Related Inflammation in Type 2 Diabetes Mellitus and Gestational Diabetes

Abstract
:1. Introduction
2. Hyperglycaemia and Inflammation—Molecular Aspects of Inflammation in Diabetes
3. Fight against Chronic Inflammation
4. PA and Inflammation in Obesity, Metabolic Syndrome and T2DM
5. Exercise During Pregnancy and GDM
6. Athletes with T2DM
7. Feasibility and Usability of PA Treatments
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- International Diabetes Federation. IDF Diabetes Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021. [Google Scholar]
- American Diabetes Association. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2021. Diabetes Care 2021, 44, 15–33. [Google Scholar] [CrossRef] [PubMed]
- Ashcroft, F.M.; Rorsman, P. Diabetes mellitus and the beta cell: The last ten years. Cell 2012, 148, 1160–1171. [Google Scholar] [CrossRef] [Green Version]
- Mannering, S.I.; Di Carluccio, A.R.; Elso, C.M. Neoepitopes: A new take on beta cell autoimmunity in type 1 diabetes. Diabetologia 2019, 62, 351–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esser, N.; Legrand-Poels, S.; Piette, J.; Scheen, A.J.; Paquot, N. Inflammation as a link between obesity, metabolic syndrome and type 2 diabetes. Diabetes Res. Clin. Pract. 2014, 105, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolan, C.J.; Damm, P.; Prentki, M. Type 2 diabetes across generations: From pathophysiology to prevention and management. Lancet 2011, 378, 169–181. [Google Scholar] [CrossRef]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Chooi, Y.C.; Ding, C.; Magkos, F. The epidemiology of obesity. Metabolism 2019, 92, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Catalano, P.M.; Tyzbi, E.D.; Wolfe, R.R.; Roman, N.M.; Amini, S.B.; Sims, E.A. Longitudinal changes in basal hepatic glucose production and suppression during insulin infusion in normal pregnant women. Am. J. Obstet. Gynecol. 1992, 167, 913–919. [Google Scholar] [CrossRef]
- Friedman, J.E.; Ishizuka, T.; Shao, J.; Huston, L.; Highman, T.; Catalano, P. Impaired glucose transport and insulin receptor tyrosine phosphorylation in skeletal muscle from obese women with gestational diabetes. Diabetes 1999, 48, 1807–1814. [Google Scholar] [CrossRef]
- Kim, C. Gestational diabetes: Risks, management, and treatment options. Int. J. Womens Health 2010, 2, 339–351. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association. Standards of medical care in diabetes-2009. Diabetes Care 2009, 32, 13–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, S.; Bock, C.; Wetzel, M.; Maul, H.; Loerbroks, A. The prevalence of gestational diabetes in advanced economies. J Perinat. Med. 2012, 40, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Catalano, P.; Huston, L.; Amini, S.; Kalhan, S. Longitudinal changes in glucose metabolism iduring pregnancy in obese women with normal glucose tollerance and GDM. Am. J. Obstet. Gynecol. 1999, 180, 903–916. [Google Scholar] [CrossRef] [PubMed]
- Silverman, B.L.; Rizzo, T.A.; Cho, N.H.; Metzger, B.E. Long-term effects of the intrauterine environment. Diabetes Care 1998, 21, 142–149. [Google Scholar]
- Dabelea, D.; Hanson, R.L.; Lindsay, R.S.; Pettitt, D.J.; Imperatore, G.; Gabir, M.M.; Roumain, J.; Bennett, P.H.; Knowler, W.C. Intrauterine exposure to diabetes conveys risks for type 2 diabetes and obesity: A study of discordant sibships. Diabetes 2000, 49, 2208–2211. [Google Scholar] [CrossRef] [Green Version]
- Bellamy, L.; Casas, J.P.; Hingorani, A.D.; Williams, D. Type 2 diabetes mellitus after gestational diabetes: A systematic review and meta-analysis. Lancet 2009, 373, 1773–1779. [Google Scholar] [CrossRef]
- Barbour, L.A.; Mccurdy, C.E.; Hernandez, T.L.; Kirwan, J.P.; Catalano, P.M.; Friedman, J.E. Cellular mechanisms for insulin resistance in normal pregnancy and gestational diabetes. Diabetes Care 2007, 30, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Kühl, C. Etiology and pathogenesis of gestational diabetes. Diabetes Care 1998, 21, 19–26. [Google Scholar]
- Plows, J.F.; Stanley, J.L.; Baker, P.N.; Reynolds, C.M.; Vickers, M.H. The pathophysiology of gestational diabetes mellitus. Int. J. Mol. Sci. 2018, 19, 3342. [Google Scholar] [CrossRef] [Green Version]
- Bianco, A.T.; Smilen, S.W.; Davis, Y.; Lopez, S.; Lapinski, R.; Lockwood, C.J. Pregnancy outcome and weight gain recommendations for the morbidly obese woman. Obstet. Gynecol. 1998, 91, 97–102. [Google Scholar] [CrossRef]
- Oteng-Ntim, E.; Mononen, S.; Sawicki, O.; Seed, P.T.; Bick, D.; Poston, L. Interpregnancy Weight Change and Adverse Pregnancy Outcomes: A Systematic Review and Meta-Analysis. BMJ Open 2018, 8, e018778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teulings, N.; Masconi, K.L.; Ozanne, S.E.; Aiken, C.E.; Wood, A.M. Effect of Interpregnancy Weight Change on Perinatal Outcomes: Systematic Review and Meta-Analysis. BMC Pregnancy Childbirth 2019, 19, 3865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmermans, Y.E.G.; van de Kant, K.D.G.; Oosterman, E.O.; Spaanderman, M.E.A.; Villamor-Martinez, E.; Kleijnen, J.; Vreugdenhil, A.C.E. The Impact of Interpregnancy Weight Change on Perinatal Outcomes in Women and Their Children: A Systematic Review and Meta-Analysis. Obes. Rev. 2020, 21, e12974. [Google Scholar] [CrossRef] [Green Version]
- Lowe, L.P.; Metzger, B.E.; Dyer, A.R.; Lowe, J.; McCance, D.R.; Lappin, T.R.; Trimble, E.R.; Coustan, D.R.; Hadden, D.R.; Hod, M.; et al. Hyperglycemia and Adverse Pregnancy Outcome (HAPO) Study: Associations of maternal A1C and glucose with pregnancy outcomes. Diabetes Care 2012, 35, 574–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, S.E.; Cooper, M.E.; Del Prato, S. Pathophysiology and treatment of type 2 diabetes: Perspectives on the past, present, and future. Lancet 2014, 383, 1068–1083. [Google Scholar] [CrossRef] [Green Version]
- Pollack, R.M.; Donath, M.Y.; LeRoith, D.; Leibowitz, G. Anti-inflammatory Agents in the Treatment of Diabetes and Its Vascular Complications. Diabetes Care 2016, 39, 244–252. [Google Scholar] [CrossRef] [Green Version]
- Jarvandi, S.; Davidson, N.O.; Jeffe, D.B.; Schootman, M. Influence of lifestyle factors on inflammation in men and women with type 2 diabetes: Results from the National Health and Nutrition Examination Survey, 1999–2004. Ann. Behav. Med. 2012, 44, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Zhang, M.; Song, F.; Qin, J.; Wang, R.; Yao, P.; Ying, C.; Hu, F.B.; Liu, L. Association between C-reactive protein and pre-diabetic status in a Chinese Han clinical population. Diabetes Metab. Res. Rev. 2009, 25, 219–223. [Google Scholar] [CrossRef]
- Eguchi, K.; Nagai, R. Islet inflammation in type 2 diabetes and physiology. J. Clin. Investig. 2017, 127, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, S.H.; Pedersen, B.K. Targeting Inflammation Through a Physical Active Lifestyle and Pharmaceuticals for the Treatment of Type 2 Diabetes. Curr. Diab. Rep. 2015, 15, 82. [Google Scholar] [CrossRef]
- McElwain, C.J.; McCarthy, F.P.; McCarthy, C.M. Gestational Diabetes Mellitus and Maternal Immune Dysregulation: What We Know So Far. Int. J. Mol. Sci. 2021, 22, 4261. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Almoros, A.; Hang, T.; Peiro, C.; Soriano-Guillen, L.; Egido, J.; Tunon, J.; Lorenzo, O. Predictive and diagnostic biomarkers for gestational diabetes and its associated metabolic and cardiovascular diseases. Cardiovasc. Diabetol. 2019, 18, 140. [Google Scholar] [CrossRef] [PubMed]
- Sheu, A.; Chan, Y.; Ferguson, A.; Bakhtyari, M.B.; Hawke, W.; White, C.; Chan, Y.F.; Bertolino, P.J.; Woon, H.G.; Palendira, U.; et al. A proinflammatory CD4+ T cell phenotype in gestational diabetes mellitus. Diabetologia 2018, 61, 1633–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, O.T.; Czech, M.P.; Corvera, S. What causes the insulin resistance underlying obesity? Curr. Opin. Endocrinol. Diabetes Obes. 2012, 19, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Shoelson, S.E.; Lee, J.; Goldfine, A.B. Inflammation and insulin resistance. J. Clin. Investig. 2006, 116, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Uysal, K.T.; Wiesbrock, S.M.; Marino, M.W.; Hotamisligil, G.S. Protection from obesity-induced insulin resistance in mice lacking TNF-alpha function. Nature 1997, 389, 610–614. [Google Scholar] [CrossRef]
- Maedler, K.; Sergeev, P.; Ris, F.; Oberholzer, J.; Joller-Jemelka, H.I.; Spinas, G.A.; Kaiser, N.; Halban, P.A.; Donath, M.Y. Glucoseinduced beta cell production of IL-1beta contributes to glucotoxicity in human pancreatic islets. J. Clin. Investig. 2002, 110, 851–860. [Google Scholar] [CrossRef]
- Zhou, R.; Tardivel, A.; Thorens, B.; Choi, I.; Tschopp, J. Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nat. Immunol. 2010, 11, 136–140. [Google Scholar] [CrossRef]
- Böni-Schnetzler, M.; Boller, S.; Debray, S.; Bouzakri, K.; Meier, D.T.; Prazak, R.; Kerr-Conte, J.; Pattou, F.; Ehses, J.A.; Schuit, F.C.; et al. Free fatty acids induce a proinflammatory response in islets via the abundantly expressed interleukin-1 receptor I. Endocrinology 2009, 150, 5218–5229. [Google Scholar] [CrossRef] [Green Version]
- Ehses, J.A.; Perren, A.; Eppler, E.; Ribaux, P.; Pospisilik, J.A.; Maor-Cahn, R.; Gueripel, X.; Ellingsgaard, H.; Schneider, M.K.; Biollaz, G.; et al. Increased number of islet-associated macrophages in type 2 diabetes. Diabetes 2007, 56, 2356–2370. [Google Scholar] [CrossRef] [Green Version]
- Richardson, S.J.; Willcox, A.; Bone, A.J.; Foulis, A.K.; Morgan, N.G. Islet-associated macrophages in type 2 diabetes. Diabetologia 2009, 52, 1686–1688. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.T.; Favelyukis, S.; Nguyen, A.K.; Reichart, D.; Scott, P.A.; Jenn, A.; Liu-Bryan, R.; Glass, C.K.; Neels, J.G.; Olefsky, J.M. A subpopulation of macrophages infiltrates hypertrophic adipose tissue and is activated by free fatty acids via Toll-like receptors 2 and 4 and JNK-dependent pathways. J. Biol. Chem. 2007, 282, 35279–35292. [Google Scholar] [CrossRef] [Green Version]
- Vandanmagsar, B.; Youm, Y.H.; Ravussin, A.; Galgani, J.E.; Stadler, K.; Mynatt, R.L.; Ravussin, E.; Stephens, J.M.; Dixit, V.D. The NLRP3 inflammasome instigates obesityinduced inflammation and insulin resistance. Nat. Med. 2011, 17, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Yuan, M.; Frantz, D.F.; Melendez, P.A.; Hansen, L.; Lee, J.; Shoelson, S.E. Local and systemic insulin resistance resulting from hepatic activation of IKK-beta and NF-kappaB. Nat. Med. 2005, 11, 183–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor alpha: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Murray, D.L.; Choy, L.N.; Spiegelman, B.M. Tumor necrosis factor α inhibits signaling from the insulin receptor. Proc. Natl. Acad. Sci. USA 1994, 91, 4854–4858. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Arner, P.; Caro, J.F.; Atkinson, R.L.; Spiegelman, B.M. Increased adipose tissue expression of tumor necrosis factor-alpha in human obesity and insulin resistance. J. Clin. Investig. 1995, 95, 2409–2415. [Google Scholar] [CrossRef]
- Qiao, Y.C.; Chen, Y.L.; Pan, Y.H.; Tian, F.; Xu, Y.; Zhang, X.X.; Zhao, H.L. The change of serum tumor necrosis factor alpha in patients with type 1 diabetes mellitus: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0176157. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Gay, M.A.; De Matias, J.M.; Gonzalez-Juanatey, C.; Garcia-Porrua, C.; Sanchez-Andrade, A.; Martin, J.; Llorca, J. Anti-tumor necrosis factor-alpha blockade improves insulin resistance in patients with rheumatoid arthritis. Clin. Exp. Rheumatol. 2006, 24, 83–86. [Google Scholar]
- Tam, L.S.; Tomlinson, B.; Chu, T.T.; Li, T.K.; Li, E.K. Impact of TNF inhibition on insulin resistance and lipids levels in patients with rheumatoid arthritis. Clin. Rheumatol. 2007, 26, 1495–1498. [Google Scholar] [CrossRef]
- Huvers, F.C.; Popa, C.; Netea, M.G.; van den Hoogen, F.H.; Tack, C.J. Improved insulin sensitivity by anti-TNF-α antibody treatment in patients with rheumatic diseases. Ann. Rheum. Dis. 2007, 66, 558–559. [Google Scholar] [CrossRef] [PubMed]
- Seriolo, B.; Ferrone, C.; Cutolo, M. Longterm anti-tumor necrosis factor-α treatment in patients with refractory rheumatoid arthritis: Relationship between insulin resistance and disease activity. J. Rheumatol. 2008, 35, 355–357. [Google Scholar] [PubMed]
- Bissell, L.A.; Hensor, E.M.; Kozera, L.; Mackie, S.L.; Burska, A.N.; Nam, J.L.; Keen, H.; Villeneuve, E.; Donica, H.; Buch, M.H.; et al. Improvement in insulin resistance is greater when infliximab is added to methotrexate during intensive treatment of early rheumatoid arthritis-results from the IDEA study. Rheumatology 2016, 55, 2181–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Febbraio, M.A. Role of interleukins in obesity: Implications for metabolic disease. Trends Endocrinol. Metab. 2014, 25, 312–319. [Google Scholar] [CrossRef]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic Adipose Tissue Inflammation Linking Obesity to Insulin Resistance and Type 2 Diabetes. Front. Physiol. 2020, 29, 1607. [Google Scholar] [CrossRef]
- Prattichizzo, F.; De Nigris, V.; Spiga, R.; Mancuso, E.; La Sala, L.; Antonicelli, R.; Testa, R.; Procopio, A.D.; Olivieri, F.; Ceriello, A. Inflammageing and metaflammation: The yin and yang of type 2 diabetes. Ageing Res. Rev. 2018, 41, 1–17. [Google Scholar] [CrossRef]
- Kuryłowicz, A.; Koźniewski, K. Anti-Inflammatory Strategies Targeting Metaflammation in Type 2 Diabetes. Molecules 2020, 25, 2224. [Google Scholar] [CrossRef]
- Karin, M.; Cao, Y.; Greten, F.R.; Li, Z.W. NF-kappaB in cancer: From innocent bystander to major culprit. Nat. Rev. Cancer 2002, 2, 301–310. [Google Scholar] [CrossRef]
- Yuan, M.; Konstantopoulos, N.; Lee, J.; Hansen, L.; Li, Z.W.; Karin, M.; Shoelson, S.E. Reversal of obesity- and diet-induced insulin resistance with salicylates or targeted disruption of Ikkbeta. Science 2001, 293, 1673–1677. [Google Scholar] [CrossRef]
- Dinarello, C.A.; Simon, A.; van der Meer, J.W. Treating inflammation by blocking interleukin-1 in a broad spectrum of diseases. Nat. Rev. Drug Discov. 2012, 8, 633–652. [Google Scholar] [CrossRef] [Green Version]
- Schroder, K.; Zhou, R.; Tschopp, J. The NLRP3 inflammasome: A sensor for metabolic danger? Science 2010, 327, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Pedra, J.H.; Cassel, S.L.; Sutterwala, F.S. Sensing pathogens and danger signals by the inflammasome. Curr. Opin. Immunol. 2009, 21, 10–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assmann, T.S.; de Almeida Brondani, L.; Boucas, A.P.; Canani, L.H.; Crispim, D. Toll-like receptor 3 (TLR3) and the development of type 1 diabetes mellitus. Arq. Bras. Endocrinol. Metabol. 2015, 59, 4–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.; Kinio, A.; Saleh, M. Functions of NOD-like receptors in human diseases. Front. Immunol. 2013, 4, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, B.Z.; Xu, Z.Q.; Han, B.Z.; Su, D.F.; Liu, C. NLRP3 inflammasome and its inhibitors: A review. Front. Pharmacol. 2015, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Davis, B.K.; Wen, H.; Ting, J.P. The inflammasome NLRs in immunity, inflammation, and associated diseases. Annu. Rev. Immunol. 2011, 29, 707–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stienstra, R.; van Diepen, J.A.; Tack, C.J.; Zaki, M.H.; van de Veerdonk, F.L.; Perera, D.; Neale, G.A.; Hooiveld, G.J.; Hijmans, A.; Vroegrijk, I.; et al. Inflammasome is a central player in the induction of obesity and insulin resistance. Proc. Natl. Acad. Sci. USA 2011, 108, 15324–15329. [Google Scholar] [CrossRef]
- Wen, H.; Gris, D.; Lei, Y.; Jha, S.; Zhang, L.; Huang, M.T.; Brickey, W.J.; Ting, J.P. Fatty acid-induced NLRP3-ASC inflammasome activation interferes with insulin signaling. Nat. Immunol. 2011, 12, 408–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rheinheimer, J.; de Souza, B.M.; Cardoso, N.S.; Bauer, A.C.; Crispim, D. Current role of the NLRP3 inflammasome on obesity and insulin resistance: A systematic review. Metabolism 2017, 74, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, V.A. Defining and characterizing the progression of type 2 diabetes. Diabetes Care 2009, 32, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Tilg, H.; Moschen, A.R. Adipocytokines: Mediators linking adipose tissue, inflammation and immunity. Nat. Rev. Immunol. 2006, 6, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Yanai, H.; Yoshida, H. Beneficial Effects of Adiponectin on Glucose and Lipid Metabolism and Atherosclerotic Progression: Mechanisms and Perspectives. Int. J. Mol. Sci. 2019, 20, 1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, A.; Nguyen, K.D.; Goh, Y.P. Macrophage-mediated inflammation in metabolic disease. Nat. Rev. Immunol. 2011, 11, 738–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 12, 1796–1808. [Google Scholar] [CrossRef]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Daniele, G.; Mendoza, R.G.; Winnier, D.; Fiorentino, T.V.; Pengou, Z.; Cornell, J.; Andreozzi, F.; Jenkinson, C.; Cersosimo, E.; Federici, M.; et al. The inflammatory status score including IL-6, TNF-α, osteopontin, fractalkine, MCP-1 and adiponectin underlies whole-body insulin resistance and hyperglycemia in type 2 diabetes mellitus. Acta Diabetol. 2013, 51, 123–131. [Google Scholar] [CrossRef]
- Schöttker, B.; Herder, C.; Rothenbacher, D.; Roden, M.; Kolb, H.; Müller, H.; Brenner, H. Proinflammatory Cytokines, Adiponectin, and Increased Risk of Primary Cardiovascular Events in Diabetic Patients With or Without Renal Dysfunction. Diabetes Care 2013, 36, 1703–1711. [Google Scholar] [CrossRef] [Green Version]
- Strissel, K.J.; Stancheva, Z.; Miyoshi, H.; Perfield, J.W., 2nd; DeFuria, J.; Jick, Z.; Greenberg, A.S.; Obin, M.S. Adipocyte death, adipose tissue remodeling, and obesity complications. Diabetes 2007, 56, 2910–2918. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.M. Subcutaneous and visceral adipose tissue: Structural and functional differences. Obes. Rev. 2010, 11, 11–18. [Google Scholar] [CrossRef]
- Rocha, A.D.S.; Bernardi, J.R.; Matos, S.; Kretzer, D.C.; Schoffel, A.C.; Goldani, M.Z.; de Azevedo Magalhaes, J.A. Maternal visceral adipose tissue during the first half of pregnancy predicts gestational diabetes at the time of delivery—A cohort study. PLoS ONE 2020, 15, e0232155. [Google Scholar] [CrossRef]
- Catalano, P.M.; Nizielski, S.E.; Shao, J.; Preston, L.; Qiao, L.; Friedman, J.E. Downregulated IRS-1 and PPARgamma in obese women with gestational diabetes: Relationship to FFA during pregnancy. Am. J. Physiol. Endocrinol. Metab. 2002, 282, 522–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa-Silva, S.; Alencar, A.P.; Moreli, J.B.; Borbely, A.U.; Lima, L.D.S.; Scavone, C.; Damasceno, D.C.; Rudge, M.V.C.; Bevilacqua, E.; Calderon, I.M.P. Hyperglycemia induces inflammatory mediators in the human chorionic villous. Cytokine 2018, 111, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen-Ngo, C.; Jayabalan, N.; Haghvirdizadeh, P.; Salomon, C.; Lappas, M. Role of adipose tissue in regulating fetal growth in gestational diabetes mellitus. Placenta 2020, 102, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Khosrowbeygi, A.; Rezvanfar, M.R.; Ahmadvand, H. Tumor necrosis factor- α, adiponectin and their ratio in gestational diabetes mellitus. Caspian J. Intern. Med. 2018, 9, 71–79. [Google Scholar] [PubMed]
- Tsiotra, P.C.; Halvatsiotis, P.; Patsouras, K.; Maratou, E.; Salamalekis, G.; Raptis, S.A.; Dimitriadis, G.; Boutati, E. Circulating adipokines and mRNA expression in adipose tissue and the placenta in women with gestational diabetes mellitus. Peptides 2018, 101, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Bhattacharjee, R.; Sur, A.; Adhikary, P.; Chowdhury, S. A study of serum growth differentiation factor 15 in Indian women with and without gestational diabetes mellitus in the third trimester of pregnancy and its association with pro-inflammatory markers and glucose metabolism. Diabetol. Int. 2021, 12, 254–259. [Google Scholar] [CrossRef]
- Gomez Ribot, D.; Diaz, E.; Fazio, M.V.; Gómez, H.L.; Fornes, D.; Macchi, S.B.; Gresta, C.A.; Capobianco, E.; Jawerbaum, A. An extra virgin olive oil-enriched diet improves maternal, placental, and cord blood parameters in GDM pregnancies. Diabetes Metab. Res. Rev. 2020, 36, e3349. [Google Scholar] [CrossRef]
- Daskalakis, G.; Marinopoulos, S.; Krielesi, V.; Papapanagiotou, A.; Papantoniou, N.; Mesogitis, S.; Antsaklis, A. Placental pathology in women with gestational diabetes. Acta Obstet. Gynecol. Scand. 2008, 87, 403–407. [Google Scholar] [CrossRef]
- Han, C.S.; Herrin, M.A.; Pitruzzello, M.C.; Mulla, M.J.; Werner, E.F.; Pettker, C.M.; Flannery, C.A.; Abrahams, V.M. Glucose and metformin modulate human first trimester trophoblast function: A model and potential therapy for diabetes-associated uteroplacental insufficiency. Am. J. Reprod. Immunol. 2015, 73, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Butryn, M.L.; Webb, V.; Wadden, T.A. Behavioral treatment of obesity. Psychiatr. Clin. N. Am. 2011, 34, 841–859. [Google Scholar] [CrossRef]
- Greenway, F.L. Physiological adaptations to weight loss and factors favouring weight regain. Int. J. Obes. 2015, 39, 1188–1196. [Google Scholar] [CrossRef] [Green Version]
- Magkos, F.; Fraterrigo, G.; Yoshino, J.; Luecking, C.; Kirbach, K.; Kelly, S.C.; de Las Fuentes, L.; He, S.; Okunade, A.L.; Patterson, B.W.; et al. Effects of moderate and subsequent progressive weight loss on metabolic function and at biology in humans with obesity. Cell Metab. 2016, 23, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlüssel, M.M.; Souza, E.B.; Reichenheim, M.E.; Kac, G. Physical activity during pregnancy and maternal-child health outcomes: A systematic literature review. Cad Saude Publica 2008, 24, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, J.C.; Butler, C.L.; Williams, M.A. No need for a pregnant pause: Physical activity may reduce the occurrence of gestational diabetes mellitus and preeclampsia. Exerc. Sport Sci. Rev. 2005, 33, 141–149. [Google Scholar] [CrossRef]
- ACOG Committee Obstetric Practice. ACOG Committee opinion. Exercise during pregnancy and the postpartum period. Obstet. Gynecol. 2002, 99, 171–173. [Google Scholar]
- American Diabetes Association. Gestational diabetes mellitus. Diabetes Care 2003, 26, 103–105. [Google Scholar] [CrossRef] [Green Version]
- Golbidi, S.; Laher, I. Potential mechanisms of exercise in gestational diabetes. J. Nutr. Metab. 2013, 2013, 285948. [Google Scholar] [CrossRef] [Green Version]
- Antunes, B.M.; Cayres, S.U.; Lira, F.S.; Fernandes, R.A. Arterial thickness and immunometabolism: The mediating role of chronic exercise. Curr. Cardiol. Rev. 2016, 12, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, G.I.; Febbraio, M.A. The immunomodulating role of exercise in metabolic disease. Trends Immunol. 2014, 35, 262–269. [Google Scholar] [CrossRef]
- Valacchi, G.; Virgili, F.; Cervellati, C.; Pecorelli, A. OxInflammation: From subclinical condition to pathological biomarker. Front. Physiol. 2018, 9, 858. [Google Scholar] [CrossRef] [Green Version]
- Koelwyn, G.J.; Wennerberg, E.; Demaria, S.; Jones, L.W. Exercise in regulation of inflammation-immune axis function in cancer initiation and progression. Oncology 2015, 29, 908–922. [Google Scholar] [PubMed]
- Tarnowski, M.; Kopytko, P.; Piotrowska, K. Epigenetic Regulation of Inflammatory Responses in the Context of Physical Activity. Genes 2021, 12, 1313. [Google Scholar] [CrossRef]
- de Lemos, E.T.; Oliveira, J.; Pinheiro, J.P.; Reis, F. Regular physical exercise as a strategy to improve antioxidant and anti-inflammatory status: Benefits in type 2 diabetes mellitus. Oxid. Med. Cell. Longev. 2012, 2012, 741545. [Google Scholar] [PubMed] [Green Version]
- Dias, M.R.J.; Correia, T.M.L.; Borges, J.O.; Santos, L.S.D.; Pereira, R.; Silva, R.A.A.D.; Soares, T.J.; Coqueiro, R.D.S.; Magalhães, A.C.M. Therapeutic or lifelong training effects on pancreatic morphological and functional parameters in an animal model of aging and obesity. Exp. Gerontol. 2023, 175, 112144. [Google Scholar] [CrossRef]
- de Lemos, E.T.; Reis, F.; Baptista, S.; Pinto, R.; Sepodes, B.; Vala, H.; Rocha-Pereira, P.; Correia da Silva, G.; Teixeira, N.; Silva, A.S.; et al. Exercise training decreases proinflammatory profile in Zucker diabetic (type 2) fatty rats. Nutrition 2009, 25, 330–339. [Google Scholar] [CrossRef]
- Carvalho, V.H.C.; Wang, Q.; Xu, X.; Liu, L.; Jiang, W.; Wang, X.; Wang, J.; Li, W.; Chen, J.; Li, T.; et al. Long-term exercise preserves pancreatic islet structure and β-cell mass through attenuation of islet inflammation and fibrosis. FASEB J. 2023, 37, e22822. [Google Scholar] [CrossRef] [PubMed]
- Tomita, T. Apoptosis in pancreatic β-islet cells in Type 2 diabetes. BJBMS 2016, 16, 162–179. [Google Scholar] [CrossRef] [Green Version]
- Christensen, C.S.; Christensen, D.P.; Lundh, M.; Dahllöf, M.S.; Haase, T.N.; Velasquez, J.M.; Laye, M.J.; Mandrup-Poulsen, T.; Solomon, T.P. Skeletal Muscle to Pancreatic beta-Cell Cross-talk: The Effect of Humoral Mediators Liberated by Muscle Contraction and Acute Exercise on beta-Cell Apoptosis. J. Clin. Endocrinol. Metab. 2015, 100, E1289-98. [Google Scholar] [CrossRef] [Green Version]
- Villaça, C.B.P.; de Paula, C.C.; de Oliveira, C.C.; Vilas-Boas, E.A.; Dos Santos-Silva, J.C.; de Oliveira, S.F.; Abdulkader, F.; Ferreira, S.M.; Ortis, F. Beneficial effects of physical exercise for beta-cell maintenance in a type 1 diabetes mellitus animal model. Exp. Physiol. 2021, 106, 1482–1497. [Google Scholar] [CrossRef]
- Zhang, T.; Tian, J.; Fan, J.; Liu, X.; Wang, R. Exercise training-attenuated insulin resistance and liver injury in elderly pre-diabetic patients correlates with NLRP3 inflammasome. Front. Immunol. 2023, 14, 1082050. [Google Scholar] [CrossRef]
- Lee, H.M.; Kim, J.J.; Kim, H.J.; Shong, M.; Ku, B.J.; Jo, E.K. Upregulated NLRP3 inflammasome activation in patients with type 2 diabetes. Diabetes 2013, 62, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Masters, S.L.; Dunne, A.; Subramanian, S.L.; Hull, R.L.; Tannahill, G.M.; Sharp, F.A.; Becker, C.; Franchi, L.; Yoshihara, E.; Chen, Z.; et al. Activation of the NLRP3 inflammasome by islet amyloid polypeptide provides a mechanism for enhanced IL-1β in type 2 diabetes. Nat. Immunol. 2010, 11, 897–904. [Google Scholar] [CrossRef]
- Sun, Y.; Ding, S. NLRP3 Inflammasome in Diabetic Cardiomyopathy and Exercise Intervention. Int. J. Mol. Sci. 2021, 22, 13228. [Google Scholar] [CrossRef] [PubMed]
- Khakroo Khakroo Abkenar, I.; Rahmani-Nia, F.; Lombardi, G.I.; Rahmani-Nia, F.; Lombardi, G. The Effects of Acute and Chronic Aerobic Activity on the Signaling Pathway of the Inflammasome NLRP3 Complex in Young Men. Medicina 2019, 55, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejías-Peña, Y.; Estébanez, B.; Rodriguez-Miguelez, P.; Fernandez-Gonzalo, R.; Almar, M.; de Paz, J.A.; González-Gallego, J.; Cuevas, M.J. Impact of resistance training on the autophagy-inflammation-apoptosis crosstalk in elderly subjects. Aging 2017, 9, 408–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyngbaek, M.P.P.; Legaard, G.E.; Bennetsen, S.L.; Feineis, C.S.; Rasmussen, V.; Moegelberg, N.; Brinkløv, C.F.; Nielsen, A.B.; Kofoed, K.S.; Lauridsen, C.A.; et al. The effects of different doses of exercise on pancreatic β-cell function in patients with newly diagnosed type 2 diabetes: Study protocol for and rationale behind the “DOSE-EX” multi-arm parallel-group randomised clinical trial. Trials 2021, 22, 244. [Google Scholar] [CrossRef]
- Curran, M.; Drayson, M.T.; Andrews, R.C.; Zoppi, C.; Barlow, J.P.; Solomon, T.P.J.; Narendran, P. The benefits of physical exercise for the health of the pancreatic β-cell: A review of the evidence. Exp. Physiol. 2020, 105, 579–589. [Google Scholar] [CrossRef]
- Ryan, A.S.; Nicklas, B.J. Reductions in plasma cytokine levels with weight loss improve insulin sensitivity in overweight and obese postmenopausal women. Diabetes Care 2004, 27, 1699–1705. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.D.; Patrizi, R.M.; Cheek, D.J.; Wooten, J.S.; Barbee, J.J.; Mitchell, J.B. Resistance training reduces subclinical inflammation in obese, postmenopausal women. Med. Sci. Sports Exerc. 2012, 44, 2099–2110. [Google Scholar] [CrossRef] [Green Version]
- Bruun, J.M.; Helge, J.W.; Richelsen, B.; Stallknecht, B. Diet and exercise reduce low-grade inflammation and macrophage infiltration in adipose tissue but not in skeletal muscle in severely obese subjects. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E961–E967. [Google Scholar] [CrossRef]
- Frøsig, C.; Richter, E.A. Improved insulin sensitivity after exercise: Focus on insulin signaling. Obesity 2009, 3, 15–20. [Google Scholar] [CrossRef]
- Maarbjerg, S.J.; Sylow, L.; Richter, E.A. Current understanding of increased insulin sensitivity after exercise–emerging candidates. Acta Physiol. 2011, 202, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Zanuso, S.; Sacchetti, M.; Sundberg, C.J.; Orlando, G.; Benvenuti, P.; Balducci, S. Exercise in type 2 diabetes: Genetic, metabolic and neuromuscular adaptations. A review of the evidence. Br. J. Sports Med. 2017, 51, 1533–1538. [Google Scholar] [CrossRef] [PubMed]
- Gholami, F.; Nikookheslat, S.; Salekzamani, Y.; Boule, N.; Jafari, A. Effect of aerobic training on nerve conduction in men with type 2 diabetes and peripheral neuropathy: A randomized controlled trial. Neurophysiol. Clin. 2018, 48, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Contrepois, K.; Wu, S.; Moneghetti, K.J.; Hornburg, D.; Ahadi, S.; Tsai, M.S.; Metwally, A.A.; Wei, E.; Lee-McMullen, B.; Quijada, J.V.; et al. Molecular Choreography of Acute Exercise. Cell 2020, 181, 1112–1130. [Google Scholar] [CrossRef]
- Savikj, M.; Stocks, B.; Sato, S.; Caidahl, K.; Krook, A.; Deshmukh, A.S.; Zierath, J.R.; Wallberg-Henriksson, H. Exercise timing influences multi-tissue metabolome and skeletal muscle proteome profiles in type 2 diabetic patients—A randomized crossover trial. Metabolism 2022, 135, 155268. [Google Scholar] [CrossRef]
- Savikj, M.; Gabriel, B.M.; Alm, P.S.; Smith, J.; Caidahl, K.; Björnholm, M.; Fritz, T.; Krook, A.; Zierath, J.R.; Wallberg-Henriksson, H. Afternoon exercise is more efficacious than morning exercise at improving blood glucose levels in individuals with type 2 diabetes: A randomised crossover trial. Diabetologia 2019, 62, 233–237. [Google Scholar] [CrossRef] [Green Version]
- Sherwani, S.I.; Khan, H.A.; Ekhzaimy, A.; Masood, A.; Sakharkar, M.K. Significance of HbA1c Test in Diagnosis and Prognosis of Diabetic Patients. Biomark. Insights 2016, 11, 95–104. [Google Scholar] [CrossRef]
- Yasunari, K.; Maeda, K.; Nakamura, M.; Yoshikawa, J. Oxidative stress in leukocytes is a possible link between blood pressure, blood glucose, and C-reacting protein. Hypertension 2002, 39, 777–780. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ye, W.; Chen, Q.; Zhang, Y.; Kuo, C.H.; Korivi, M. Resistance Exercise Intensity is Correlated with Attenuation of HbA1c and Insulin in Patients with Type 2 Diabetes: A Systematic Review and Meta-Analysis. Int. J. Environ. Res. Public Health 2019, 16, 140. [Google Scholar] [CrossRef] [Green Version]
- Elsisia, H.F.; Aneisb, Y.M.; Mounirc, K.M. Impact of high-intensity interval training on HbA1c in patients with type 2 diabetes mellitus. Bull. Fac. Phys. Ther. 2015, 20, 168–175. [Google Scholar] [CrossRef]
- Winding, K.M.; Munch, G.W.; Iepsen, U.W.; Van Hall, G.; Pedersen, B.K.; Mortensen, S.P. The effect on glycaemic control of low-volume high-intensity interval training versus endurance training in individuals with type 2 diabetes. Diabetes Obes. Metab. 2018, 20, 1131–1139. [Google Scholar] [CrossRef] [Green Version]
- Nesto, R.W. Correlation between cardiovascular disease and diabetes mellitus: Current concepts. Am. J. Med. 2004, 116, 11S–22S. [Google Scholar] [CrossRef] [PubMed]
- Perks, J.; Zaccardi, F.; Rayt, H.; Sayers, R.; Brady, E.M.; Davies, M.J.; Rowlands, A.V.; Edwardson, C.L.; Hall, A.; Yates, T.; et al. Device-measured physical activity behaviours, and physical function, in people with type 2 diabetes mellitus and peripheral artery disease: A cross-sectional study. Exp. Gerontol. 2023, 15, 112207. [Google Scholar] [CrossRef] [PubMed]
- Jorge, M.L.; de Oliveira, V.N.; Resende, N.M.; Paraiso, L.F.; Calixto, A.; Diniz, A.L.; Resende, E.S.; Ropelle, E.R.; Carvalheira, J.B.; Espindola, F.S.; et al. The effects of aerobic, resistance, and combined exercise on metabolic control, inflammatory markers, adipocytokines, and muscle insulin signaling in patients with type 2 diabetes mellitus. Metabolism 2011, 60, 1244–1252. [Google Scholar] [CrossRef]
- Magalhães, J.P.; Melo, X.; Correia, I.R.; Ribeiro, R.T.; Raposo, J.; Dores, H.; Bicho, M.; Sardinha, L.B. Effects of combined training with different intensities on vascular health in patients with type 2 diabetes: A 1-year randomized controlled trial. Cardiovasc. Diabetol. 2019, 18, 34. [Google Scholar] [CrossRef] [Green Version]
- Garneau, L.; Terada, T.; Mistura, M.; Mulvihill, E.E.; Reed, J.L.; Aguer, C. Exercise training reduces circulating cytokines in male patients with coronary artery disease and type 2 diabetes: A pilot study. Physiol. Rep. 2023, 11, 15634. [Google Scholar] [CrossRef]
- Persons, J.E.; Pierce, G.L.; Fiedorowicz, J.G. Anxiety, C-reactive protein, and obesity in NHANES 1999–2004. Heliyon 2019, 5, e02267. [Google Scholar] [CrossRef] [Green Version]
- Trøseid, M.; Lappegård, K.T.; Claudi, T.; Damås, J.K.; Mørkrid, L.; Brendberg, R.; Mollnes, T.E. Exercise reduces plasma levels of the chemokines MCP-1 and IL-8 in subjects with the metabolic syndrome. Eur. Heart J. 2004, 25, 349–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomeleri, C.M.; Souza, M.F.; Burini, R.C.; Cavaglieri, C.R.; Ribeiro, A.S.; Antunes, M.; Nunes, J.P.; Venturini, D.; Barbosa, D.S.; Sardinha, L.B.; et al. Resistance training reduces metabolic syndrome and inflammatory markers in older women: A randomized controlled trial. J. Diabetes 2018, 10, 328–337. [Google Scholar] [CrossRef]
- Fuentes, G.C.; Castañer, O.; Warnberg, J.; Subirana, I.; Buil-Cosiales, P.; Salas-Salvadó, J.; Corella, D.; Serra-Majem, L.; Romaguera, D.; Estruch, R.; et al. Prospective association of physical activity and inflammatory biomarkers in older adults from the PREDIMED-Plus study with overweight or obesity and metabolic syndrome. Clin. Nutr. 2020, 39, 3092–3098. [Google Scholar] [CrossRef]
- Del Rosso, S.; Baraquet, M.L.; Barale, A.; Defagó, M.D.; Tortosa, F.; Perovic, N.R.; Aoki, M.P. Long-term effects of different exercise training modes on cytokines and adipokines in individuals with overweight/obesity and cardiometabolic diseases: A systematic review, meta-analysis, and meta-regression of randomized controlled trials. Obes. Rev. 2023, 24, e13564. [Google Scholar] [CrossRef] [PubMed]
- Papagianni, G.; Panayiotou, C.; Vardas, M.; Balaskas, N.; Antonopoulos, C.; Tachmatzidis, D.; Didangelos, T.; Lambadiari, V.; Kadoglou, N.P.E. The anti-inflammatory effects of aerobic exercise training in patients with type 2 diabetes: A systematic review and meta-analysis. Cytokine 2023, 164, 156157. [Google Scholar] [CrossRef]
- Dekker, M.J.; Lee, S.; Hudson, R.; Kilpatrick, K.; Graham, T.E.; Ross, R.; Robinson, L.E. An exercise intervention without weight loss decreases circulating interleukin-6 in lean and obese men with and without type 2 diabetes mellitus. Metabolism 2007, 56, 332–338. [Google Scholar] [CrossRef]
- Dadrass, A.; Mohamadzadeh Salamat, K.; Hamidi, K.; Azizbeigi, K. Anti-inflammatory effects of vitamin D and resistance training in men with type 2 diabetes mellitus and vitamin D deficiency: A randomized, double-blinded, placebo-controlled clinical trial. J. Diabetes Metab. Disord. 2019, 18, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Arslan, M.; Ipekci, S.H.; Kebapcilar, L.; Dogan Dede, N.; Kurban, S.; Erbay, E.; Gonen, M.S. Effect of Aerobic Exercise Training on MDA and TNF- alpha Levels in Patients with Type 2 Diabetes Mellitus. Int. Sch. Res. Notices 2014, 2014, 820387. [Google Scholar]
- Hoene, M.; Zhao, X.; Machann, J.; Birkenfeld, A.L.; Heni, M.; Peter, A.; Niess, A.; Moller, A.; Lehmann, R.; Xu, G.; et al. Exercise-Induced N-Lactoylphenylalanine Predicts Adipose Tissue Loss during Endurance Training in Overweight and Obese Humans. Metabolites 2023, 13, 15. [Google Scholar] [CrossRef] [PubMed]
- Chow, L.S.; Gerszten, R.E.; Tylor, J.M.; Pedersen, B.K.; van Praag, H.; Trappe, S.; Febbraio, M.A.; Galis, Z.S.; Gao, Y.; Haus, J.M.; et al. Exerkines in health, resilience and disease. Nat. Rev. Endocrinol. 2022, 18, 273–289. [Google Scholar] [CrossRef]
- Li, V.L.; He, Y.; Contrepois, K.; Liu, H.; Kim, J.T.; Wiggenhorn, A.L.; Tanzo, J.T.; Tung, A.S.; Lyu, X.; Zushin, P.H.; et al. An exercise-inducible metabolite that suppresses feeding and obesity. Nature 2022, 606, 785–790. [Google Scholar] [CrossRef]
- Neuparth, M.J.; Proença, J.B.; Santos-Silva, A.; Coimbra, S. The positive effect of moderate walking exercise on chemerin levels in Portuguese patients with type 2 diabetes mellitus. J. Investig. Med. 2014, 62, 350–353. [Google Scholar] [CrossRef]
- Kadoglou, N.P.; Fotiadis, G.; Kapelouzou, A.; Kostakis, A.; Liapis, C.D.; Vrabas, I.S. The differential anti-inflammatory effects of exercise modalities and their association with early carotid atherosclerosis progression in patients with type 2 diabetes. Diabet. Med. 2013, 30, e41–e50. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Kader, M.S.; Al-Jiffri, O.; Ashmawy, E.M. Impact of weight loss on markers of systemic inflammation in obese Saudi children with asthma. Afr. Health Sci. 2013, 13, 682–688. [Google Scholar] [PubMed] [Green Version]
- Borges, L.; Passos, M.E.P.; Silva, M.B.B.; Santos, V.C.; Momesso, C.M.; Pithon-Curi, T.C.; Gorjão, R.; Gray, S.R.; Lima, K.C.A.; de Freitas, P.B.; et al. Dance Training Improves Cytokine Secretion and Viability of Neutrophils in Diabetic Patients. Mediators Inflamm. 2019, 20, 2924818. [Google Scholar] [CrossRef] [Green Version]
- Asle Mohammadi Zadeh, M.; Kargarfard, M.; Marandi, S.M.; Habibi, A. Diets along with interval training regimes improves inflammatory & anti-inflammatory condition in obesity with type 2 diabetes subjects. J. Diabetes Metab. Disord. 2018, 17, 253–267. [Google Scholar]
- Hooshmand Moghadam, B.; Rashidlamir, A.; Attarzadeh Hosseini, S.R.; Gaeini, A.A.; Kaviani, M. The effects of saffron (Crocus sativus L.) in conjunction with concurrent training on body composition, glycaemic status, and inflammatory markers in obese men with type 2 diabetes mellitus: A randomized double-blind clinical trial. Br. J. Clin. Pharmacol. 2022, 88, 3256–3271. [Google Scholar] [CrossRef]
- Ried-Larsen, M.; Johansen, M.Y.; MacDonald, C.S.; Hansen, K.B.; Christensen, R.; Wedell-Neergaard, A.S.; Pilmark, N.S.; Langberg, H.; Vaag, A.A.; Pedersen, B.K.; et al. Type 2 diabetes remission 1 year after an intensive lifestyle intervention: A secondary analysis of a randomized clinical trial. Diabetes Obes. Metab. 2019, 10, 2257–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregg, E.W.; Chen, H.; Wagenknecht, L.E.; Clark, J.M.; Delahanty, L.M.; Bantle, J.; Pownall, H.J.; Johnson, K.C.; Safford, M.M.; Kitabchi, A.E.; et al. Association of an intensive lifestyle intervention with remission of type 2 diabetes. Look AHEAD Research Group. JAMA 2012, 308, 2489–2496. [Google Scholar] [CrossRef] [Green Version]
- Badon, S.E.; Enquobahrie, D.A.; Wartko, P.D.; Miller, R.S.; Qiu, C.; Gelaye, B.; Sorensen, T.K.; Williams, M.A. Healthy Lifestyle During Early Pregnancy and Risk of Gestational Diabetes Mellitus. Am. J. Epidemiol. 2017, 186, 326–333. [Google Scholar] [CrossRef] [Green Version]
- Dipla, K.; Zafeiridis, A.; Mintziori, G.; Boutou, A.K.; Goulis, D.G.; Hackney, A.C. Exercise as a Therapeutic Intervention in Gestational Diabetes Mellitus. Endocrines 2021, 2, 65–78. [Google Scholar] [CrossRef]
- Rand, B.G.; Johnson, T.M.; Ehrlich, S.F.; Wideman, L.; Pivarnik, J.M.; Richardson, M.R.; Stone, M.L.; Churilla, J.R. Diabetes risk status and physical activity in pregnancy: U.S. BRFSS 2011, 2013, 2015, 2017. BMC Pregnancy Childbirth 2020, 20, 743. [Google Scholar] [CrossRef]
- de Oliveira Santini, C.; Imakawa, T.dS.; Duarte, G.; Quintana, S.M.; Moise’s, E.C.D. Do the body mass index and the diagnosis of gestational diabetes mellitus influence the level of physical activity during pregnancy and postpartum? PLoS ONE 2019, 14, e0220947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renault, K.; Carlsen, E.; Hædersdal, S.; Nilas, L.; Secher, N.J.; Eugen-Olsen, J.; Cortes, D.; Olsen, S.F.; Halldorsson, T.I.; Nørgaard, K. Impact of lifestyle intervention for obese women during pregnancy on maternal metabolic and inflammatory markers. Int. J. Obes. 2017, 41, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Loprinzi, P.D.; Fitzgerald, E.M.; Woekel, E.; Cardinal, B.J. Association of physical activity and sedentary behavior with biological markers among U.S. pregnant women. J. Womens Health 2013, 22, 953–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Zhu, W.; Wei, Y.; Wang, C.; Zhu, W.; Wei, Y.; Feng, H.; Su, R.; Yang, H. Exercise intervention during pregnancy can be used to manage weight gain and improve pregnancy outcomes in women with gestational diabetes mellitus. BMC Pregnancy Childbirth 2015, 15, 255. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Cupul-Uicab, L.A.; Rogan, W.J.; Eggesbo, M.; Travlos, G.; Wilson, R.; Longnecker, M.P. Recreational Exercise Before and During Pregnancy in Relation to Plasma C-Reactive Protein Concentrations in Pregnant Women. J. Phys. Act. Health 2015, 12, 770–775. [Google Scholar] [CrossRef] [Green Version]
- Padmapriya, N.; Bernard, J.Y.; Liang, S.; Loy, S.L.; Cai, S.; Zhe, I.S.; Kwek, K.; Godfrey, K.M.; Gluckman, P.D.; Saw, S.M.; et al. Associations of physical activity and sedentary behavior during pregnancy with gestational diabetes mellitus among Asian women in Singapore. BMC Pregnancy Childbirth 2017, 17, 364. [Google Scholar] [CrossRef] [Green Version]
- Simmons, D.; Jelsma, J.G.; Galjaard, S.; Devlieger, R.; van Assche, A.; Jans, G.; Corcoy, R.; Adelantado, J.M.; Dunne, F.; Desoye, G.; et al. Results From a European Multicenter Randomized Trial of Physical Activity and/or Healthy Eating to Reduce the Risk of Gestational Diabetes Mellitus: The DALI Lifestyle Pilot. Diabetes Care 2015, 38, 1650–1656. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, S.F.; Sternfeld, B.; Krefman, A.E.; Hedderson, M.M.; Brown, S.D.; Mevi, A.; Chasan-Taber, L.; Quesenberry, C.P., Jr.; Ferrara, A. Moderate and Vigorous Intensity Exercise during Pregnancy and Gestational Weight Gain in Women with Gestational Diabetes. Matern. Child Health J. 2016, 20, 1247–1257. [Google Scholar] [CrossRef] [Green Version]
- Nobles, C.; Marcus, B.H.; Stanek, E.J., 3rd; Braun, B.; Whitcomb, B.W.; Solomon, C.G.; Manson, J.E.; Markenson, G.; Chasan-Taber, L. Effect of an exercise intervention on gestational diabetes mellitus: A randomized controlled trial. Obstet. Gynecol. 2015, 125, 1195–1204. [Google Scholar] [CrossRef] [Green Version]
- Gou, B.H.; Guan, H.M.; Bi, Y.X.; Ding, B.J. Gestational diabetes: Weight gain during pregnancy and its relationship to pregnancy outcomes. Chin. Med. J. 2019, 132, 154–160. [Google Scholar] [CrossRef]
- Embaby, H.; Elsayed, E.; Fawzy, M. Insulin Sensitivity and Plasma Glucose Response to Aerobic Exercise in Pregnant Women at Risk for Gestational Diabetes Mellitus. Ethiop. J. Health Sci. 2016, 26, 409–414. [Google Scholar] [CrossRef]
- Da Silva, S.G.; Hallal, P.C.; Domingues, M.R.; Bertoldi, A.D.; Silveira, M.F.D.; Bassani, D.; da Silva, I.C.M.; da Silva, B.G.C.; Coll, C.V.N.; Evenson, K. A randomized controlled trial of exercise during pregnancy on maternal and neonatal outcomes: Results from the PAMELA study. Int. J. Behav. Nutr. Phys. Act 2017, 14, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barakat, R.; Pelaez, M.; Lopez, C.; Lucia, A.; Ruiz, J.R. Exercise during pregnancy and gestational diabetes-related adverse effects: A randomised controlled trial. Br. J. Sports Med. 2013, 47, 630–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barakat, R.; Franco, E.; Perales, M.; López, C.; Mottola, M.F. Exercise during pregnancy is associated with a shorter duration of labor. A randomized clinical trial. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 224, 33–40. [Google Scholar] [CrossRef]
- Bossick, A.S.; Peters, R.M.; Burmeister, C.; Kakumanu, N.; Shill, J.E.; Cassidy-Bushrow, A.E. Antenatal inflammation and gestational diabetes mellitus risk among pregnant African-American women. J. Reprod. Immunol. 2016, 115, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Alamolhoda, S.H.; Yazdkhasti, M.; Namdari, M.; Zakariayi, S.J.; Mirabi, P. Association between C-reactive protein and gestational diabetes: A prospective study. Obstet. Gynaecol. 2020, 40, 349–353. [Google Scholar] [CrossRef]
- Hawkins, M.; Pekow, P.; Chasan-Taber, L. Physical activity, sedentary behavior, and C-reactive protein in pregnancy. Med. Sci. Sports Exerc. 2014, 46, 284–292. [Google Scholar] [CrossRef]
- Tinius, R.A.; Cahill, A.G.; Strand, E.A.; Cade, W.T. Maternal inflammation during late pregnancy is lower in physically active compared with inactive obese women. Appl. Physiol. Nutr. Metab. 2016, 41, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Manzano, P.; Acosta, F.M.; Femia, P.; Coll-Risco, I.; Segura-Jiménez, V.; Díaz-Castro, J.; Ochoa-Herrera, J.J.; Van Poppel, M.N.M.; Aparicio, V.A. Association of sedentary time and physical activity levels with immunometabolic markers in early pregnancy: The GESTAFIT project. Scand. J. Med. Sci. Sports 2020, 30, 148–158. [Google Scholar] [CrossRef]
- Acosta-Manzano, P.; Coll-Risco, I.; Van Poppel, M.N.M.; Segura-Jiménez, V.; Femia, P.; Romero-Gallardo, L.; Borges-Cosic, M.; Díaz-Castro, J.; Moreno-Fernández, J.; Ochoa-Herrera, J.J. Influence of a Concurrent Exercise Training Intervention during Pregnancy on Maternal and Arterial and Venous Cord Serum Cytokines: The GESTAFIT Project. J. Clin. Med. 2019, 8, 1862. [Google Scholar] [CrossRef] [Green Version]
- Aparicio, V.A.; Ocón, O.; Diaz-Castro, J.; Acosta-Manzano, P.; Coll-Risco, I.; Borges-Cósic, M.; Romero-Gallardo, L.; Moreno-Fernández, J.; Ochoa-Herrera, J.J. Influence of a Concurrent Exercise Training Program During Pregnancy on Colostrum and Mature Human Milk Inflammatory Markers: Findings From the GESTAFIT Project. J. Hum. Lact. 2018, 34, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Trojian, T.; Colberg, S.; Harris, G.; Oh, R.; Dixit, S.; Dixit, S.; Gibson, M.; Corcoran, M.; Ramey, L.; Berg, P.V. American Medical Society for Sports Medicine Position Statement on the Care of the Athlete and Athletic Person With Diabetes. Clin. J. Sport Med. 2022, 32, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Parisi, E.J.; Baggish, A.L. Exercise and Cardiovascular Risk among Masters Athletes with Type 2 Diabetes. Curr. Diabetes Rep. 2019, 19, 127. [Google Scholar] [CrossRef] [PubMed]
- Climstein, M.; Walsh, J.; Adams, K.; Sevene, T.; Heazlewood, T.; DeBeliso, M. Prevalence of hyperglycemia in masters athletes. PeerJ 2022, 10, e13389. [Google Scholar] [CrossRef]
- Laine, M.K.; Eriksson, J.G.; Kujala, U.M.; Wasenius, N.S.; Kaprio, J.; Bäckmand, H.M.; Peltonen, M.; Mertsalmi, T.H.; Sarna, S. A former career as a male elite athlete--does it protect against type 2 diabetes in later life? Diabetologia 2014, 57, 270–274. [Google Scholar] [CrossRef]
- Sacre, J.W.; Jellis, C.L.; Jenkins, C.; Haluska, B.A.; Baumert, M.; Coombes, J.S.; Marwick, T.H. A six-month exercise intervention in subclinical diabetic heart disease: Effects on exercise capacity, autonomic and myocardial function. Metabolism 2014, 63, 1104–1114. [Google Scholar] [CrossRef] [Green Version]
- La Gerche, A.; Burns, A.T.; Mooney, D.J.; Inder, W.J.; Taylor, A.J.; Bogaert, J.; Macisaac, A.I.; Heidbüchel, H.; Prior, D.L. Exercise-induced right ventricular dysfunction and structural remodelling in endurance athletes. Eur. Heart J. 2012, 33, 998–1006. [Google Scholar] [CrossRef] [Green Version]
- Calvo, N.; Brugada, J.; Sitges, M.; Mont, L. Atrial fibrillation and atrial flutter in athletes. Br. J. Sports Med. 2012, 46, 37–43. [Google Scholar] [CrossRef]
- Wilder, R.P.; Cicchetti, M. Common injuries in athletes with obesity and diabetes. Clin. Sports Med. 2009, 28, 441–453. [Google Scholar] [CrossRef]
- Macknight, J.M.; Mistry, D.J.; Pastors, J.G.; Holmes, V.; Rynders, C.A. The daily management of athletes with diabetes. Clin. Sports Med. 2009, 28, 479–495. [Google Scholar] [CrossRef]
- Knowler, W.C.; Barrett-Connor, E.; Fowler, S.E.; Hamman, R.F.; Lachin, J.M.; Walker, E.A.; Nathan, D.M. Diabetes Prevention Program Research Group. Reduction in the incidence of type 2 diabetes with lifestyle intervention or metformin. N. Engl. J. Med. 2002, 346, 393–403. [Google Scholar]
- Bannell, D.J.; France-Ratcliffe, M.; Buckley, B.J.R.; Crozier, A.; Davies, A.P.; Hesketh, K.L.; Jones, H.; Cocks, M.; Sprung, V.S.; MOTIVATE Team. Adherence to unsupervised exercise in sedentary individuals: A randomised feasibility trial of two mobile health interventions. Digit. Health 2023, 9, 20552076231183552. [Google Scholar] [CrossRef] [PubMed]
- Niemeijer, A.; Lund, H.; Stafne, S.N.; Ipsen, T.; Goldschmidt, C.L.; Jørgensen, C.T.; Juhl, C.B. Adverse events of exercise therapy in randomised controlled trials: A systematic review and meta-analysis. Br. J. Sports Med. 2020, 54, 1073–1080. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, S.R.; Pedersen, M.E.; Skou, S.T.; Ried-Larsen, M. Online Physical Exercise and Group Sessions to Increase and Maintain Physical Activity in Individuals with Type 2 Diabetes: A Single-Arm Feasibility Study. Int. J. Environ. Res. Public Health 2023, 20, 2893. [Google Scholar] [CrossRef] [PubMed]
- Delrieu, L.; Pialoux, V.; Pérol, O.; Morelle, M.; Martin, A.; Friedenreich, C.; Febvey-Combes, O.; Pérol, D.; Belladame, E.; Clémençon, M.; et al. Feasibility and Health Benefits of an Individualized Physical Activity Intervention in Women With Metastatic Breast Cancer: Intervention Study. JMIR mHealth uHealth 2020, 8, e12306. [Google Scholar] [CrossRef]
- Daly, R.M.; Gianoudis, J.; Hall, T.; Mundell, N.L.; Maddison, R. Feasibility, Usability, and Enjoyment of a Home-Based Exercise Program Delivered via an Exercise App for Musculoskeletal Health in Community-Dwelling Older Adults: Short-term Prospective Pilot Study. JMIR mHealth uHealth 2021, 9, e21094. [Google Scholar] [CrossRef] [PubMed]
- Schoeppe, S.; Salmon, J.; Williams, S.L.; Power, D.; Alley, S.; Rebar, A.L.; Hayman, M.; Duncan, M.J.; Vandelanotte, C. Effects of an Activity Tracker and App Intervention to Increase Physical Activity in Whole Families-The Step It Up Family Feasibility Study. Int. J. Environ. Res. Public Health 2020, 17, 7655. [Google Scholar] [CrossRef]


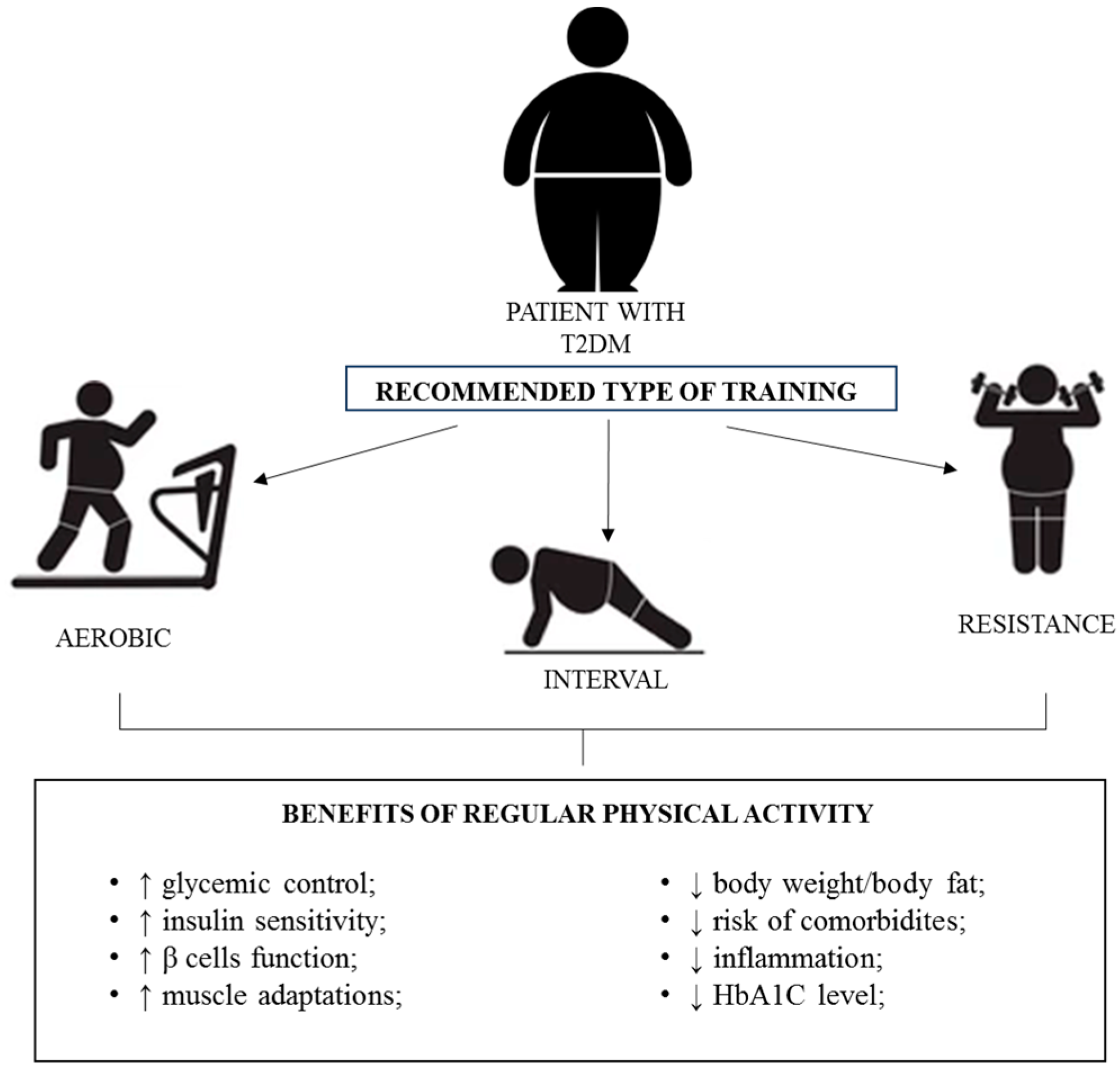{kind=link}
| Number of Participants | Age and Gender | Type of PA | Duration of PA | Results (Inflammatory Status) | Reference |
|---|---|---|---|---|---|
| 83 | M, F | Walking (self-reported) | 12 mo | ↓CRP | [151] |
| 100 | M, F, 56–70 y.a. | AT, RT, AT + RT | 4/week, 6 mo, 60 min/session | ↓hs-CRP (AT, AT + RT) | [152] |
| 48 | M, F, 53.9 ± 9.9 y.a. | AT, RT, AT + RT | 3/week, 3 mo, 60 min/session | ↓hs-CRP, ↑TNF = α | [136] |
| 50 | M, F, 40–55 y.a. | AT: mild (55–65% HRmax) Moderate (65–75% HRmax) | 3/week, 3 mo, 40 min/session | ↓TNF = α, IL-2, IL-4, IL-6 | [153] |
| 24 (ob/L)ob + T2DM | M, mean 47 y.a. | AT (about 60% VO2max) | 5/week, 3 mo, 60 min/session | ↓IL-6, ≈CRP | [145] |
| 10T2DM 12 c | F, 60–73 y.a. | AT (Dance classes) | 2/week, 4 mo, 60 min/session | ↓CRP, ↓TNF-α, (T2DM Control) ≈ IL-1β, ↑IL-1ra (C) ≈ IL-1ra (T2DM) | [154] |
| 23 ob 12 c | F, 60–70 y.a. | RT (moderate-high intensity) | 3/week, 3 mo, (10 ex. 8–10 rep. max.) | ↓CRP, ↓TNF-α, ↑WBC (shortly after training) | [120] |
| 42 T2DM 20 c | M, F, mean 52,6 y.a. | AT RT | 3/week,3 mo, 40–60min/session; 3/week, 3 mo, resistance band | ↓TLR4, IL-18, ↑IL-33 (but lower than c) | [131] |
| 31 ex 33 c | M, F, mean 53.5 y.a. | AT | 3/week, at least 12 sessions/2 mo | ↓MDA,≈ TNF-α | [147] |
| 33 T2DM 9 c | M 47 ± 5 y.a. | HIIT + diet | 20 min/session, 3/week, 12 weeks | ↓ IL-6, TNF-α | [155] |
| 48 T2DM | M 40–65 y.a. | RT + vit.D suppl. | 3/week, 12 weeks, 50 min/session | ↓ IL-6, TNF-α, ≈ CRP | [146] |
| 52 T2DM | M 39 ± 5 y.a. | AT+RT + saffron suppl. | 3/week, 12 weeks, 1–1.5 h/session | ↓IL-6, TNF-α, hsCRP, IL-1β, ↑IL-10 | [156] |
| T2DM | M 40–74 y.a. | MICT vs. HIIT | 12-week, twice weekly | ↓FGF21, IL-6, IL-8, IL-10 and IL-18. | [138] |
| 1239 T2DM | M, F | AT | meta-analysis | ↓CRP, resistin, TNF-α, IL-6 | [144] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piotrowska, K.; Zgutka, K.; Tkacz, M.; Tarnowski, M. Physical Activity as a Modern Intervention in the Fight against Obesity-Related Inflammation in Type 2 Diabetes Mellitus and Gestational Diabetes. Antioxidants 2023, 12, 1488. https://doi.org/10.3390/antiox12081488
Piotrowska K, Zgutka K, Tkacz M, Tarnowski M. Physical Activity as a Modern Intervention in the Fight against Obesity-Related Inflammation in Type 2 Diabetes Mellitus and Gestational Diabetes. Antioxidants. 2023; 12(8):1488. https://doi.org/10.3390/antiox12081488
Chicago/Turabian StylePiotrowska, Katarzyna, Katarzyna Zgutka, Marta Tkacz, and Maciej Tarnowski. 2023. "Physical Activity as a Modern Intervention in the Fight against Obesity-Related Inflammation in Type 2 Diabetes Mellitus and Gestational Diabetes" Antioxidants 12, no. 8: 1488. https://doi.org/10.3390/antiox12081488
APA StylePiotrowska, K., Zgutka, K., Tkacz, M., & Tarnowski, M. (2023). Physical Activity as a Modern Intervention in the Fight against Obesity-Related Inflammation in Type 2 Diabetes Mellitus and Gestational Diabetes. Antioxidants, 12(8), 1488. https://doi.org/10.3390/antiox12081488