
Selenium-Enriched E. coli Bacteria Mitigate the Age-Associated Degeneration of Cholinergic Neurons in C. elegans

 , , , ,
, , , ,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture and Treatment of E. coli OP50 Bacteria
2.2. Determination of the Se Content in Bacterial Growth Media and Pellets
2.3. Maintenance and Treatment of C. elegans
2.4. Determination of Body Length and Area of the Nematodes
2.5. Fecundity of the Nematodes
2.6. Determination of the Pharyngeal Pumping Rate of the Nematodes
2.7. Bacteria Choice Assay
2.8. Fluorescence Microscopy of the GFP-Tagged C. elegans Strains LOK158 and LX929
2.9. Statistical Analysis
3. Results and Discussion
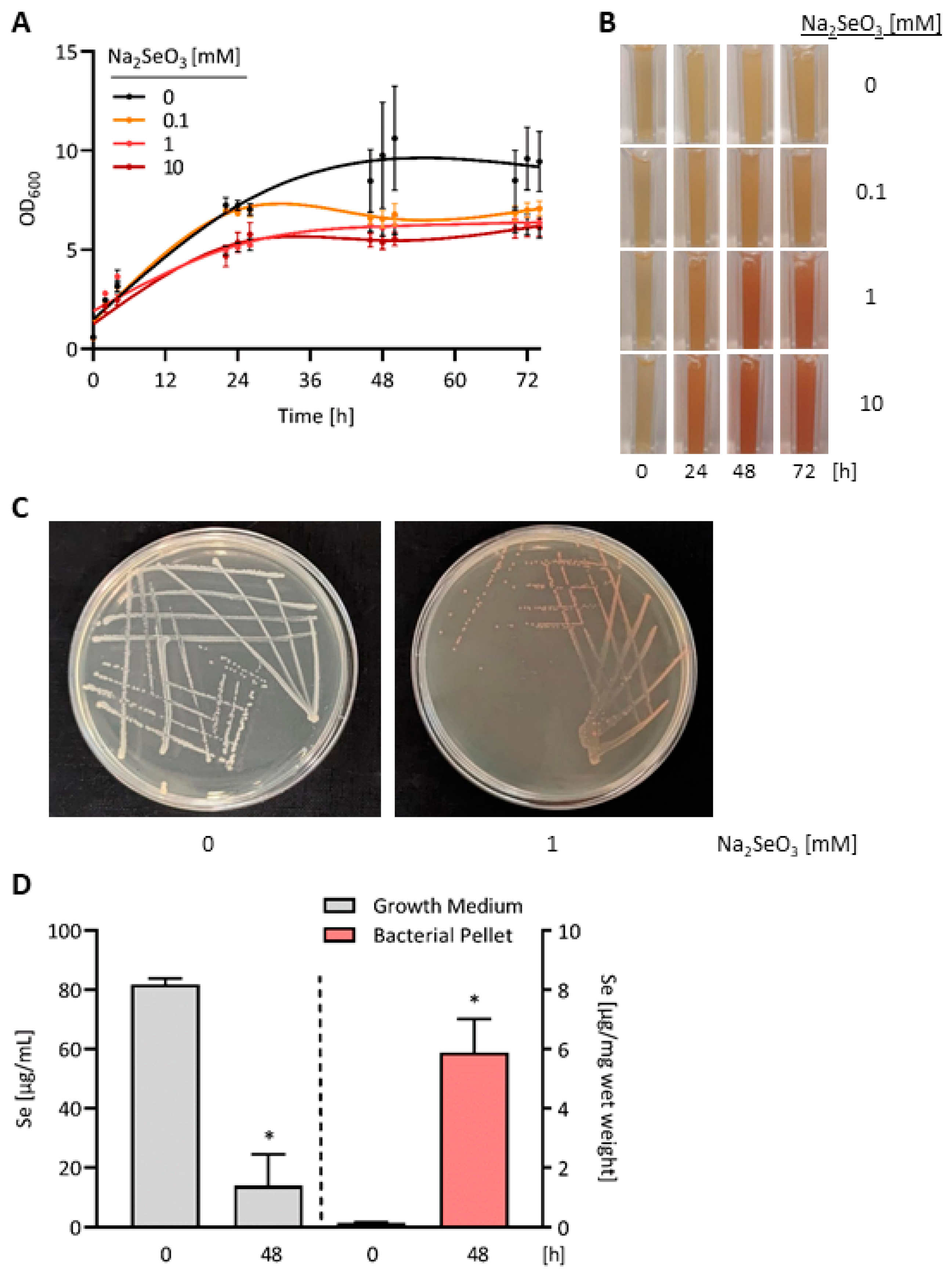
3.1. Accumulation of Red Elemental Se in E. coli OP50 Bacteria Exposed to High Doses of Selenite
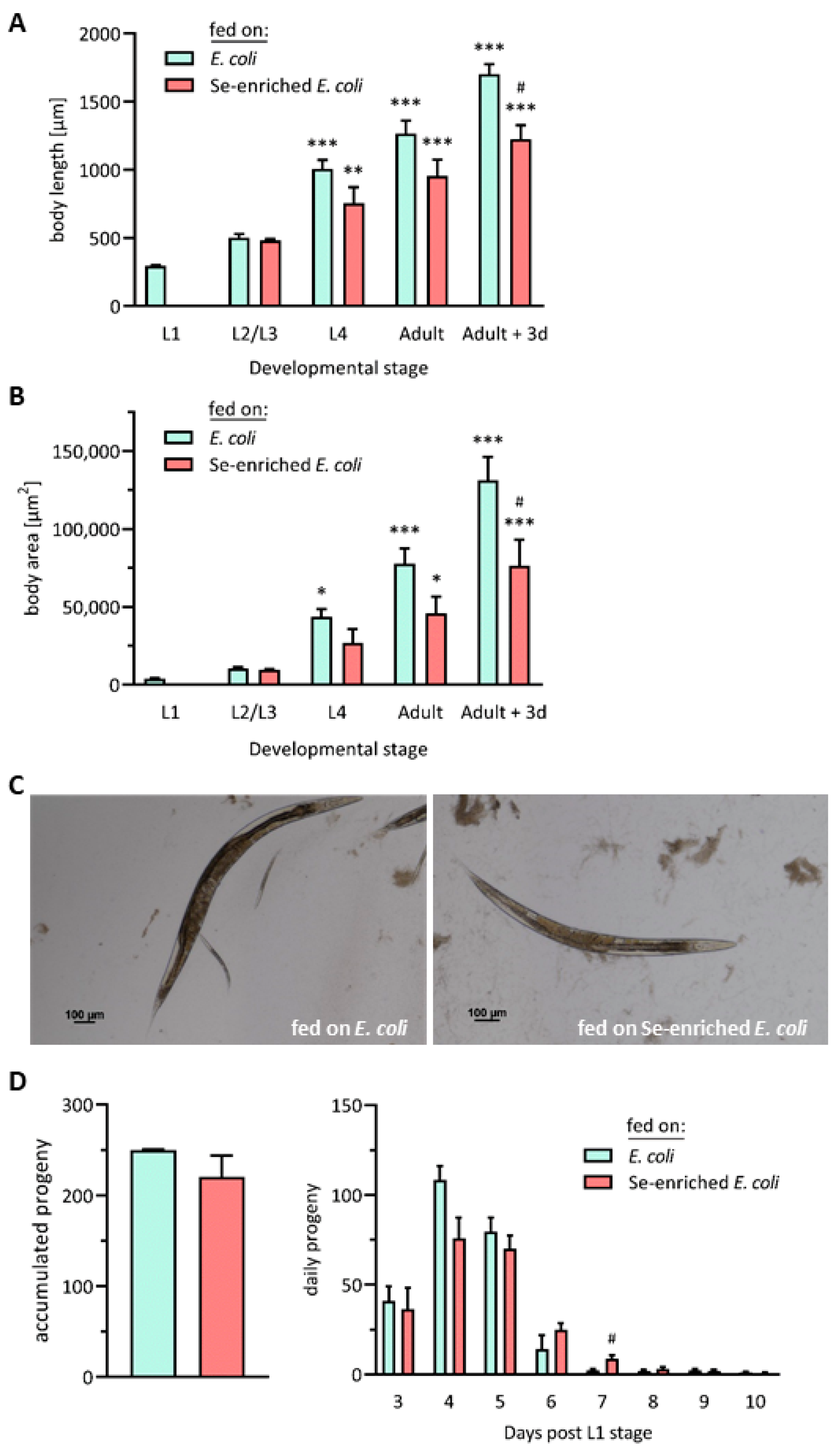
3.2. Influence of Se-Enriched E. coli OP50 Bacteria on Vital Parameters of C. elegans
3.3. Feeding Se-Enriched E. coli OP50 Bacteria Results in Elevated Levels of the Se-Binding Protein SEMO-1 in C. elegans
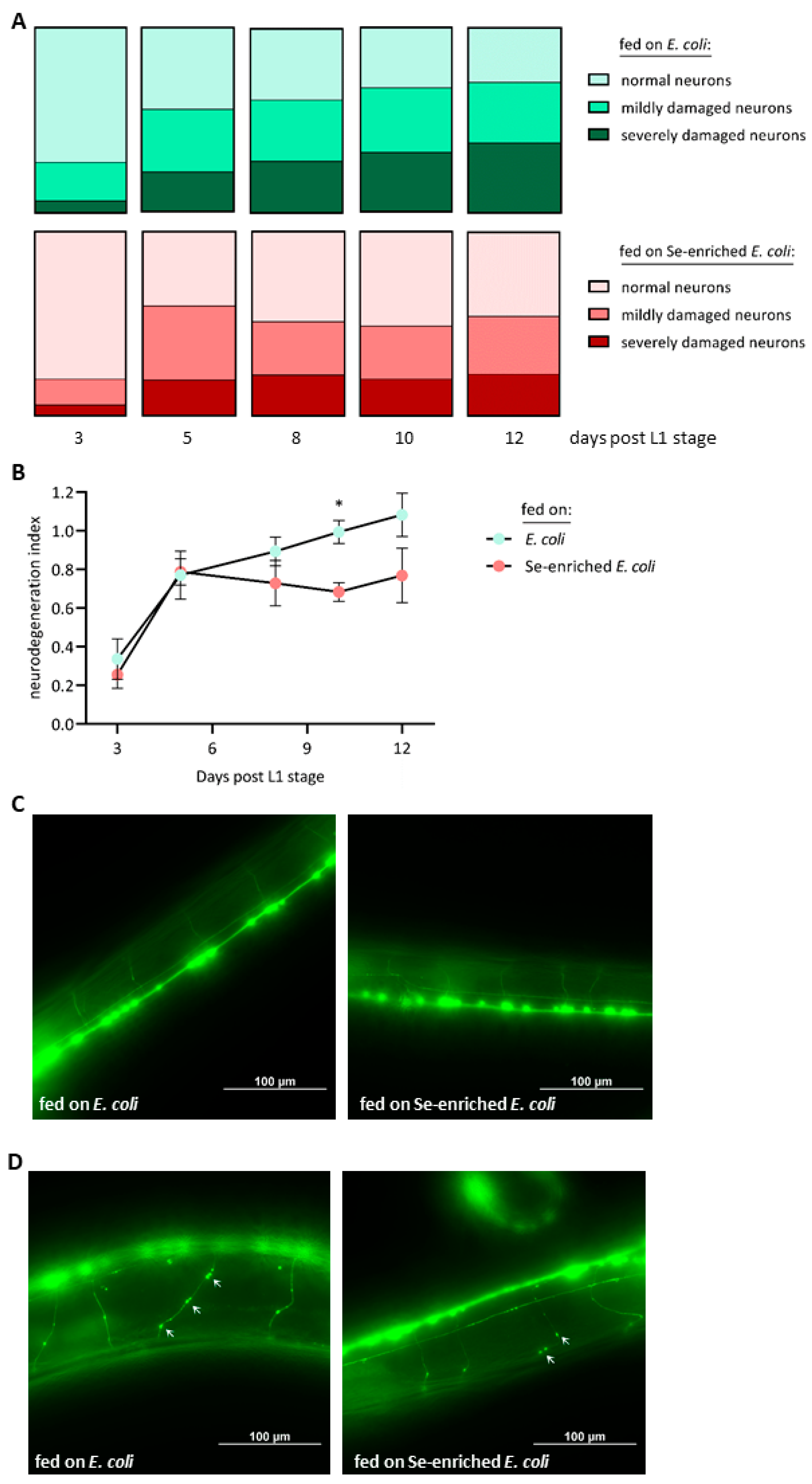
3.4. Se-Enriched E. coli OP50 Bacteria Mitigate the Progressive Damage of Cholinergic Neurons in Aging Nematodes
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steinbrenner, H.; Klotz, L.O. Selenium and zinc: “Antioxidants” for healthy aging? Z. Gerontol. Geriatr. 2020, 53, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, H.; Speckmann, B.; Klotz, L.O. Selenoproteins: Antioxidant selenoenzymes and beyond. Arch. Biochem. Biophys. 2016, 595, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.L.U.; Sena-Evangelista, K.C.M.; de Azevedo, E.P.; Pinheiro, F.I.; Cobucci, R.N.; Pedrosa, L.F.C. Selenium in human health and gut microflora: Bioavailability of selenocompounds and relationship with diseases. Front. Nutr. 2021, 8, 685317. [Google Scholar] [CrossRef] [PubMed]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA); Turck, D.; Bohn, T.; Castenmiller, J.; de Henauw, S.; Hirsch-Ernst, K.I.; Knutsen, H.K.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; et al. Scientific opinion on the tolerable upper intake level for selenium. EFSA J. 2023, 21, e07704. [Google Scholar] [PubMed]
- Hadrup, N.; Ravn-Haren, G. Toxicity of repeated oral intake of organic selenium, inorganic selenium, and selenium nanoparticles: A review. J. Trace Elem. Med. Biol. 2023, 79, 127235. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohe, R.; Flohe, L. Selenium and redox signaling. Arch. Biochem. Biophys. 2017, 617, 48–59. [Google Scholar] [CrossRef]
- Hoefig, C.S.; Renko, K.; Kohrle, J.; Birringer, M.; Schomburg, L. Comparison of different selenocompounds with respect to nutritional value vs. Toxicity using liver cells in culture. J. Nutr. Biochem. 2011, 22, 945–955. [Google Scholar] [CrossRef]
- Nie, X.; Yang, X.; He, J.; Liu, P.; Shi, H.; Wang, T.; Zhang, D. Bioconversion of inorganic selenium to less toxic selenium forms by microbes: A review. Front. Bioeng. Biotechnol. 2023, 11, 1167123. [Google Scholar] [CrossRef]
- Ullah, A.; Mu, J.; Wang, F.; Chan, M.W.H.; Yin, X.; Liao, Y.; Mirani, Z.A.; Sebt, E.H.S.; Aslam, S.; Naveed, M.; et al. Biogenic selenium nanoparticles and their anticancer effects pertaining to probiotic bacteria-a review. Antioxidants 2022, 11, 1916. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Qi, M.; Wu, Q.; Xiang, P.; Tang, D.; Li, Q. Recent research progress on the synthesis and biological effects of selenium nanoparticles. Front. Nutr. 2023, 10, 1183487. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Dou, X.; Song, X.; Chang, J.; Zeng, X.; Zhu, L.; Yi, H.; Xu, C. Replacing dietary sodium selenite with biogenic selenium nanoparticles improves the growth performance and gut health of early-weaned piglets. Anim. Nutr. 2023, 15, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Qiao, L.; Ma, L.; Guo, Y.; Dou, X.; Yan, S.; Zhang, B.; Roman, A. Biogenic selenium nanoparticles synthesized by lactobacillus casei atcc 393 alleviate intestinal epithelial barrier dysfunction caused by oxidative stress via nrf2 signaling-mediated mitochondrial pathway. Int. J. Nanomed. 2019, 14, 4491–4502. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Chen, Y.; Song, X.; Dou, X.; Xu, C. Selenium nanoparticles-enriched lactobacillus casei atcc 393 prevents cognitive dysfunction in mice through modulating microbiota-gut-brain axis. Int. J. Nanomed. 2022, 17, 4807–4827. [Google Scholar] [CrossRef]
- Ji, H.F.; Shen, L. Probiotics as potential therapeutic options for alzheimer’s disease. Appl. Microbiol. Biotechnol. 2021, 105, 7721–7730. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, H.; Sies, H. Selenium homeostasis and antioxidant selenoproteins in brain: Implications for disorders in the central nervous system. Arch. Biochem. Biophys. 2013, 536, 152–157. [Google Scholar] [CrossRef]
- Tamtaji, O.R.; Heidari-Soureshjani, R.; Mirhosseini, N.; Kouchaki, E.; Bahmani, F.; Aghadavod, E.; Tajabadi-Ebrahimi, M.; Asemi, Z. Probiotic and selenium co-supplementation, and the effects on clinical, metabolic and genetic status in alzheimer’s disease: A randomized, double-blind, controlled trial. Clin. Nutr. 2019, 38, 2569–2575. [Google Scholar] [CrossRef]
- Lee, H.; Lee, S.V. Recent progress in regulation of aging by insulin/igf-1 signaling in caenorhabditis elegans. Mol. Cells 2022, 45, 763–770. [Google Scholar] [CrossRef]
- Shen, P.; Yue, Y.; Zheng, J.; Park, Y. Caenorhabditis elegans: A convenient in vivo model for assessing the impact of food bioactive compounds on obesity, aging, and alzheimer’s disease. Annu. Rev. Food Sci. Technol. 2018, 9, 1–22. [Google Scholar] [CrossRef]
- Roussos, A.; Kitopoulou, K.; Borbolis, F.; Palikaras, K. Caenorhabditis elegans as a model system to study human neurodegenerative disorders. Biomolecules 2023, 13, 478. [Google Scholar] [CrossRef] [PubMed]
- Estevez, A.O.; Mueller, C.L.; Morgan, K.L.; Szewczyk, N.J.; Teece, L.; Miranda-Vizuete, A.; Estevez, M. Selenium induces cholinergic motor neuron degeneration in caenorhabditis elegans. Neurotoxicology 2012, 33, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Hsu, F.L.; Liu, J.T.; Liao, V.H. The ameliorative and toxic effects of selenite on caenorhabditis elegans. Food Chem. Toxicol. 2011, 49, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Rohn, I.; Marschall, T.A.; Kroepfl, N.; Jensen, K.B.; Aschner, M.; Tuck, S.; Kuehnelt, D.; Schwerdtle, T.; Bornhorst, J. Selenium species-dependent toxicity, bioavailability and metabolic transformations in caenorhabditis elegans. Metallomics 2018, 10, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Rohn, I.; Raschke, S.; Aschner, M.; Tuck, S.; Kuehnelt, D.; Kipp, A.; Schwerdtle, T.; Bornhorst, J. Treatment of caenorhabditis elegans with small selenium species enhances antioxidant defense systems. Mol. Nutr. Food Res. 2019, 63, e1801304. [Google Scholar] [CrossRef] [PubMed]
- Taskov, K.; Chapple, C.; Kryukov, G.V.; Castellano, S.; Lobanov, A.V.; Korotkov, K.V.; Guigo, R.; Gladyshev, V.N. Nematode selenoproteome: The use of the selenocysteine insertion system to decode one codon in an animal genome? Nucleic Acids Res. 2005, 33, 2227–2238. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The genetics of caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Zecic, A.; Dhondt, I.; Braeckman, B.P. The nutritional requirements of caenorhabditis elegans. Genes. Nutr. 2019, 14, 15. [Google Scholar] [CrossRef] [PubMed]
- Kohnlein, K.; Urban, N.; Guerrero-Gomez, D.; Steinbrenner, H.; Urbanek, P.; Priebs, J.; Koch, P.; Kaether, C.; Miranda-Vizuete, A.; Klotz, L.O. A caenorhabditis elegans ortholog of human selenium-binding protein 1 is a pro-aging factor protecting against selenite toxicity. Redox Biol. 2020, 28, 101323. [Google Scholar] [CrossRef]
- Philipp, T.M.; Gong, W.; Kohnlein, K.; Ohse, V.A.; Muller, F.I.; Priebs, J.; Steinbrenner, H.; Klotz, L.O. Semo-1, a novel methanethiol oxidase in caenorhabditis elegans, is a pro-aging factor conferring selective stress resistance. Biofactors 2022, 48, 699–706. [Google Scholar] [CrossRef]
- Köhnlein, K.; Urban, N.; Steinbrenner, H.; Guerrero-Gómez, D.; Miranda-Vizuete, A.; Kaether, C.; Klotz, L.O. Selenite-induced expression of a caenorhabditis elegans pro-aging factor and ortholog of human selenium-binding protein 1. Curr. Neutraceuticals 2020, 1, 73–79. [Google Scholar] [CrossRef]
- Philipp, T.M.; Gernoth, L.; Will, A.; Schwarz, M.; Ohse, V.A.; Kipp, A.P.; Steinbrenner, H.; Klotz, L.O. Selenium-binding protein 1 (selenbp1) is a copper-dependent thiol oxidase. Redox Biol. 2023, 65, 102807. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Lossow, K.; Schirl, K.; Hackler, J.; Renko, K.; Kopp, J.F.; Schwerdtle, T.; Schomburg, L.; Kipp, A.P. Copper interferes with selenoprotein synthesis and activity. Redox Biol. 2020, 37, 101746. [Google Scholar] [CrossRef] [PubMed]
- Abada, E.A.; Sung, H.; Dwivedi, M.; Park, B.J.; Lee, S.K.; Ahnn, J.C. Elegans behavior of preference choice on bacterial food. Mol. Cells 2009, 28, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Margie, O.; Palmer, C.; Chin-Sang, I.C. Elegans chemotaxis assay. J. Vis. Exp. 2013, 27, e50069. [Google Scholar]
- Turner, R.J.; Weiner, J.H.; Taylor, D.E. Selenium metabolism in escherichia coli. Biometals 1998, 11, 223–227. [Google Scholar] [CrossRef]
- Tetteh, A.Y.; Sun, K.H.; Hung, C.Y.; Kittur, F.S.; Ibeanu, G.C.; Williams, D.; Xie, J. Transcriptional response of selenopolypeptide genes and selenocysteine biosynthesis machinery genes in escherichia coli during selenite reduction. Int. J. Microbiol. 2014, 2014, 394835. [Google Scholar] [CrossRef] [PubMed]
- Ahluwalia, G.S.; Saxena, Y.R.; Williams, H.H. Quantitative studies on selenite metabolism in escherichia coli. Arch. Biochem. Biophys. 1968, 124, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Martin, G.; Sanz-Landaluze, J.; Leon-Gonzalez, M.E.; Madrid, Y. In vivo quantification of volatile organoselenium compounds released by bacteria exposed to selenium with hs-spme-gc-ms. Effect of selenite and selenium nanoparticles. Talanta 2021, 224, 121907. [Google Scholar] [CrossRef]
- Gerrard, T.L.; Telford, J.N.; Williams, H.H. Detection of selenium deposits in escherichia coli by electron microscopy. J. Bacteriol. 1974, 119, 1057–1060. [Google Scholar] [CrossRef]
- Boehler, C.J.; Raines, A.M.; Sunde, R.A. Deletion of thioredoxin reductase and effects of selenite and selenate toxicity in caenorhabditis elegans. PLoS ONE 2013, 8, e71525. [Google Scholar] [CrossRef] [PubMed]
- Garza-Garcia, J.J.O.; Hernandez-Diaz, J.A.; Zamudio-Ojeda, A.; Leon-Morales, J.M.; Guerrero-Guzman, A.; Sanchez-Chipres, D.R.; Lopez-Velazquez, J.C.; Garcia-Morales, S. The role of selenium nanoparticles in agriculture and food technology. Biol. Trace Elem. Res. 2022, 200, 2528–2548. [Google Scholar] [CrossRef] [PubMed]
- Al Ghanem, A.; Nasim, M.J.; Alnahas, F.; Ney, Y.; Weiss, A.V.; Koch, M.; Schneider, M.; Jacob, C. Incredible edible selenium nanoparticles produced by food-grade microorganisms. Curr. Neutraceuticals 2021, 2, 135–144. [Google Scholar] [CrossRef]
- Estevam, E.C.; Griffin, S.; Nasim, M.J.; Denezhkin, P.; Schneider, R.; Lilischkis, R.; Dominguez-Alvarez, E.; Witek, K.; Latacz, G.; Keck, C.; et al. Natural selenium particles from staphylococcus carnosus: Hazards or particles with particular promise? J. Hazard. Mater. 2017, 324, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, L.S.; Walhout, A.J. Worms, bacteria, and micronutrients: An elegant model of our diet. Trends Genet. 2014, 30, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Boehler, C.J.; Raines, A.M.; Sunde, R.A. Toxic-selenium and low-selenium transcriptomes in caenorhabditis elegans: Toxic selenium up-regulates oxidoreductase and down-regulates cuticle-associated genes. PLoS ONE 2014, 9, e101408. [Google Scholar] [CrossRef] [PubMed]
- Chai, V.Z.; Farajzadeh, T.; Meng, Y.; Lo, S.B.; Asaed, T.A.; Taylor, C.J.; Glater, E.E. Chemical basis of microbiome preference in the nematode c. Elegans. Sci. Rep. 2024, 14, 1350. [Google Scholar] [CrossRef] [PubMed]
- Worthy, S.E.; Haynes, L.; Chambers, M.; Bethune, D.; Kan, E.; Chung, K.; Ota, R.; Taylor, C.J.; Glater, E.E. Identification of attractive odorants released by preferred bacterial food found in the natural habitats of c. Elegans. PLoS ONE 2018, 13, e0201158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lu, H.; Bargmann, C.I. Pathogenic bacteria induce aversive olfactory learning in caenorhabditis elegans. Nature 2005, 438, 179–184. [Google Scholar] [CrossRef]
- Stenvall, J.; Fierro-Gonzalez, J.C.; Swoboda, P.; Saamarthy, K.; Cheng, Q.; Cacho-Valadez, B.; Arner, E.S.; Persson, O.P.; Miranda-Vizuete, A.; Tuck, S. Selenoprotein trxr-1 and gsr-1 are essential for removal of old cuticle during molting in caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2011, 108, 1064–1069. [Google Scholar] [CrossRef]
- Schliebs, R.; Arendt, T. The cholinergic system in aging and neuronal degeneration. Behav. Brain Res. 2011, 221, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Herndon, L.A.; Schmeissner, P.J.; Dudaronek, J.M.; Brown, P.A.; Listner, K.M.; Sakano, Y.; Paupard, M.C.; Hall, D.H.; Driscoll, M. Stochastic and genetic factors influence tissue-specific decline in ageing c. Elegans. Nature 2002, 419, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.L.; Peng, C.Y.; Chen, C.H.; McIntire, S. Genetic analysis of age-dependent defects of the caenorhabditis elegans touch receptor neurons. Proc. Natl. Acad. Sci. USA 2011, 108, 9274–9279. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.M.; Waddell, B.M.; Wu, C.W. Neuron-specific toxicity of chronic acrylamide exposure in c. Elegans. Neurotoxicol. Teratol. 2020, 77, 106848. [Google Scholar] [CrossRef] [PubMed]
- Parrales-Macias, V.; Michel, P.P.; Tourville, A.; Raisman-Vozari, R.; Haik, S.; Hunot, S.; Bizat, N.; Lannuzel, A. The pesticide chlordecone promotes parkinsonism-like neurodegeneration with tau lesions in midbrain cultures and c. Elegans worms. Cells 2023, 12, 1336. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Park, S.J.; Lange, M.; Tseyang, T.; Doshi, M.B.; Kim, T.Y.; Song, Y.; Kim, D.I.; Greer, P.L.; Olzmann, J.A.; et al. Selenium reduction of ubiquinone via sqor suppresses ferroptosis. Nat. Metab. 2024, 6, 343–358. [Google Scholar] [CrossRef]
- Chambers, I.G.; Ratan, R.R. Selenium abandons selenoproteins to inhibit ferroptosis rapidly. Nat. Metab. 2024, 6, 200–202. [Google Scholar] [CrossRef] [PubMed]
- Mayorga-Weber, G.; Rivera, F.J.; Castro, M.A. Neuron-glia (mis)interactions in brain energy metabolism during aging. J. Neurosci. Res. 2022, 100, 835–854. [Google Scholar] [CrossRef]
- Berr, C.; Arnaud, J.; Akbaraly, T.N. Selenium and cognitive impairment: A brief-review based on results from the eva study. Biofactors 2012, 38, 139–144. [Google Scholar] [CrossRef]
- Kryscio, R.J.; Abner, E.L.; Caban-Holt, A.; Lovell, M.; Goodman, P.; Darke, A.K.; Yee, M.; Crowley, J.; Schmitt, F.A. Association of antioxidant supplement use and dementia in the prevention of alzheimer’s disease by vitamin e and selenium trial (preadvise). JAMA Neurol. 2017, 74, 567–573. [Google Scholar] [CrossRef]
- Ewald, C.Y.; Li, C. Caenorhabditis elegans as a model organism to study app function. Exp. Brain Res. 2012, 217, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Schmidlin, C.J.; Dodson, M.B.; Madhavan, L.; Zhang, D.D. Redox regulation by nrf2 in aging and disease. Free Radic. Biol. Med. 2019, 134, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Cheng, Y.; Li, X.; Wang, F.; Lu, Z.; Xiao, X.; Wang, Y. Biogenic nanoselenium particles effectively attenuate oxidative stress-induced intestinal epithelial barrier injury by activating the nrf2 antioxidant pathway. ACS Appl. Mater. Interfaces 2017, 9, 14724–14740. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zytner, P.; Kutschbach, A.; Gong, W.; Ohse, V.A.; Taudte, L.; Kipp, A.P.; Klotz, L.-O.; Priebs, J.; Steinbrenner, H. Selenium-Enriched E. coli Bacteria Mitigate the Age-Associated Degeneration of Cholinergic Neurons in C. elegans. Antioxidants 2024, 13, 492. https://doi.org/10.3390/antiox13040492
Zytner P, Kutschbach A, Gong W, Ohse VA, Taudte L, Kipp AP, Klotz L-O, Priebs J, Steinbrenner H. Selenium-Enriched E. coli Bacteria Mitigate the Age-Associated Degeneration of Cholinergic Neurons in C. elegans. Antioxidants. 2024; 13(4):492. https://doi.org/10.3390/antiox13040492
Chicago/Turabian StyleZytner, Palina, Anne Kutschbach, Weiye Gong, Verena Alexia Ohse, Laura Taudte, Anna Patricia Kipp, Lars-Oliver Klotz, Josephine Priebs, and Holger Steinbrenner. 2024. "Selenium-Enriched E. coli Bacteria Mitigate the Age-Associated Degeneration of Cholinergic Neurons in C. elegans" Antioxidants 13, no. 4: 492. https://doi.org/10.3390/antiox13040492
APA StyleZytner, P., Kutschbach, A., Gong, W., Ohse, V. A., Taudte, L., Kipp, A. P., Klotz, L. -O., Priebs, J., & Steinbrenner, H. (2024). Selenium-Enriched E. coli Bacteria Mitigate the Age-Associated Degeneration of Cholinergic Neurons in C. elegans. Antioxidants, 13(4), 492. https://doi.org/10.3390/antiox13040492