


The Role of NOX2-Derived Reactive Oxygen Species in the Induction of Endothelin-Converting Enzyme-1 by Angiotensin II
 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Methods
2.1. Materials
2.2. Cell Culture
2.3. Immunoassays
2.4. Gene Expression Analysis
2.5. Promoter Analysis, Plasmid DNA Constructs
2.6. Cell Transfections
2.7. Electronic Shift Mobility Assay (EMSA)
2.8. Animal Studies
2.9. Statistical Analysis
3. Results
3.1. Role of ECE-1 in ET-1 Production by HMECs Stimulated with Ang II
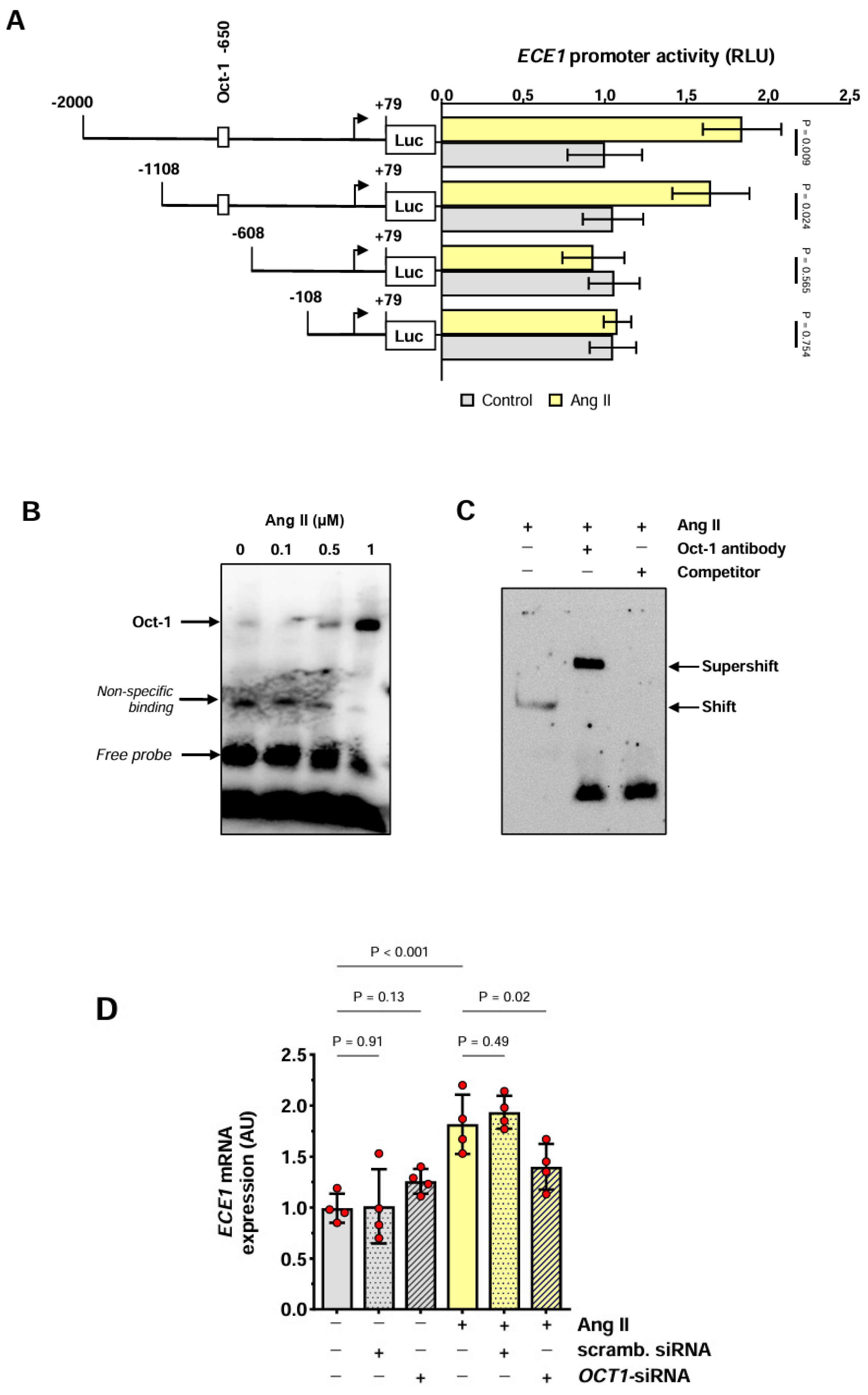
3.2. Induction of ECE1 Expression by Ang II in HMECs
3.3. Effect of Ang II on the ECE1 Promoter in HMECs
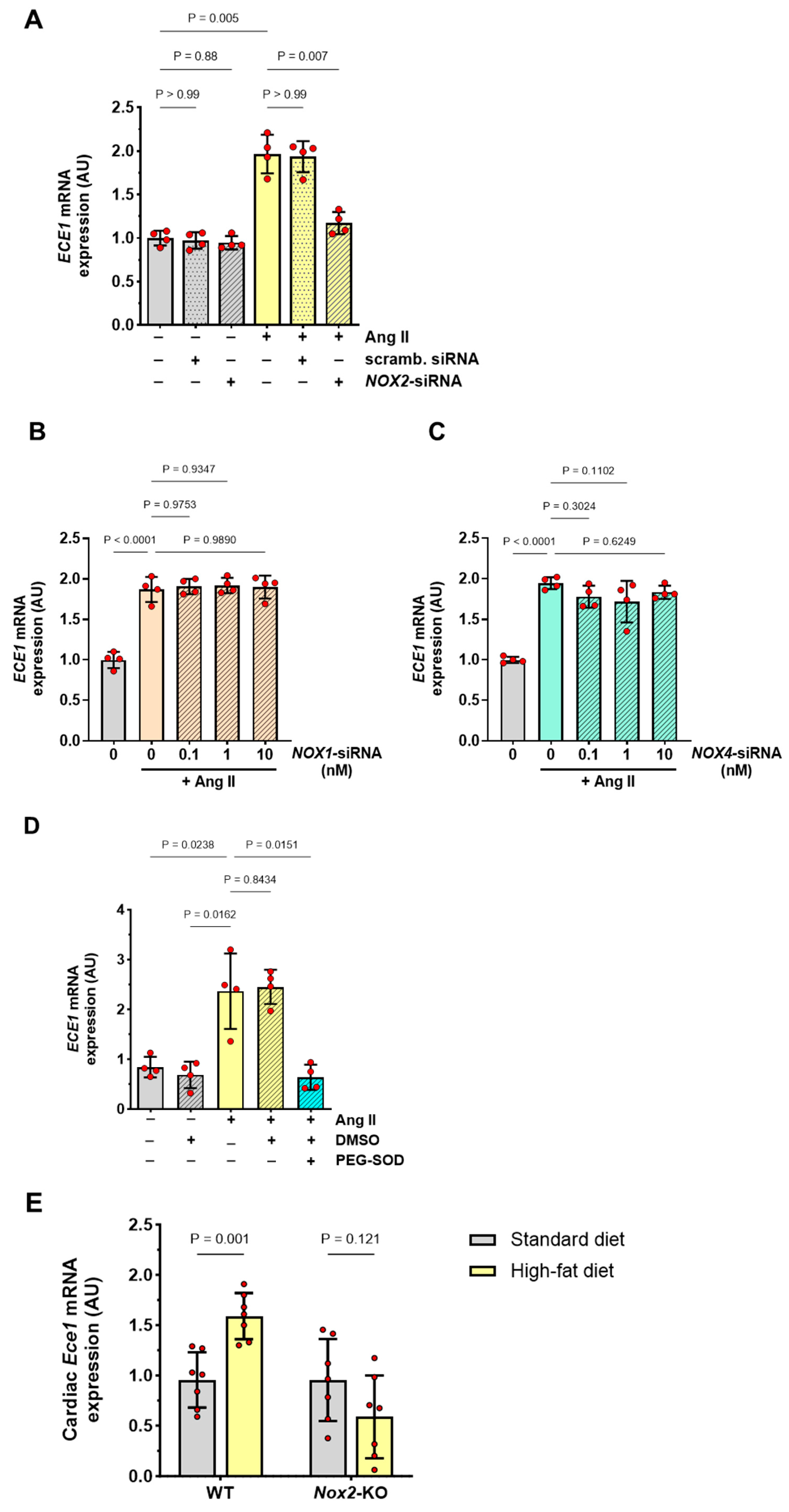
3.4. Role of NOX2-Derived Superoxide Anions in the Induction of ECE1 Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davenport, A.P.; Hyndman, K.A.; Dhaun, N.; Southan, C.; Kohan, D.E.; Pollock, J.S.; Pollock, D.M.; Webb, D.J.; Maguire, J.J. Endothelin. Pharmacol. Rev. 2016, 68, 357–418. [Google Scholar] [CrossRef] [PubMed]
- Stow, L.R.; Jacobs, M.E.; Wingo, C.S.; Cain, B.D. Endothelin-1 gene regulation. Faseb J. 2011, 25, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, A.; Valdenaire, O.; Nelböck, P.; Deuschle, U.; Dumas Milne Edwards, J.B.; Stumpf, J.G.; Löffler, B.M. Human endothelin-converting enzyme (ECE-1): Three isoforms with distinct subcellular localizations. Biochem. J. 1997, 328 Pt 3, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Valdenaire, O.; Lepailleur-Enouf, D.; Egidy, G.; Thouard, A.; Barret, A.; Vranckx, R.; Tougard, C.; Michel, J.B. A fourth isoform of endothelin-converting enzyme (ECE-1) is generated from an additional promoter molecular cloning and characterization. Eur. J. Biochem. 1999, 264, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Orzechowski, H.D.; Richter, C.M.; Funke-Kaiser, H.; Kröger, B.; Schmidt, M.; Menzel, S.; Bohnemeier, H.; Paul, M. Evidence of alternative promoters directing isoform-specific expression of human endothelin-converting enzyme-1 mRNA in cultured endothelial cells. J. Mol. Med. 1997, 75, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Funke-Kaiser, H.; Bolbrinker, J.; Theis, S.; Lemmer, J.; Richter, C.M.; Paul, M.; Orzechowski, H.D. Characterization of the c-specific promoter of the gene encoding human endothelin-converting enzyme-1 (ECE-1). FEBS Lett. 2000, 466, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, H.; Yanagisawa, M.; Kapur, R.P.; Richardson, J.A.; Williams, S.C.; Clouthier, D.E.; de Wit, D.; Emoto, N.; Hammer, R.E. Dual genetic pathways of endothelin-mediated intercellular signaling revealed by targeted disruption of endothelin converting enzyme-1 gene. Development 1998, 125, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.D.; Stevenson, T.; Ahn, K. Hydrolysis of peptide hormones by endothelin-converting enzyme-1. A comparison with neprilysin. J. Biol. Chem. 1999, 274, 4053–4058. [Google Scholar] [CrossRef] [PubMed]
- Fecteau, M.H.; Honoré, J.C.; Plante, M.; Labonté, J.; Rae, G.A.; D’Orléans-Juste, P. Endothelin-1 (1-31) is an intermediate in the production of endothelin-1 after big endothelin-1 administration in vivo. Hypertension 2005, 46, 87–92. [Google Scholar] [CrossRef]
- D’Orléans-Juste, P.; Houde, M.; Rae, G.A.; Bkaily, G.; Carrier, E.; Simard, E. Endothelin-1 (1-31): From chymase-dependent synthesis to cardiovascular pathologies. Vasc. Pharmacol. 2008, 49, 51–62. [Google Scholar] [CrossRef]
- Kamhieh-Milz, J.; Chen, L.; Goettsch, C.; Pfefferkorn, A.M.; Hofmann, A.; Brunssen, C.; Müller, G.M.; Walther, T.; Ashraf, M.I.; Moll, G.; et al. Ang II Promotes ET-1 Production by Regulating NOX2 Activity Through Transcription Factor Oct-1. Arter. Thromb. Vasc. Biol. 2023, 43, 1429–1440. [Google Scholar] [CrossRef] [PubMed]
- Niemann, B.; Rohrbach, S.; Catar, R.A.; Muller, G.; Barton, M.; Morawietz, H. Native and oxidized low-density lipoproteins stimulate endothelin-converting enzyme-1 expression in human endothelial cells. Biochem. Biophys. Res. Commun. 2005, 334, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T.; Kurihara, H.; Takahashi, M.; Shimada, K.; Maemura, K.; Oda, H.; Ishikawa, T.; Uchiyama, T.; Tanzawa, K.; Yazaki, Y.J.C. Endothelin-converting enzyme expression in the rat vascular injury model and human coronary atherosclerosis. Circulation 1997, 95, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Chi, J.T.; Chang, H.Y.; Haraldsen, G.; Jahnsen, F.L.; Troyanskaya, O.G.; Chang, D.S.; Wang, Z.; Rockson, S.G.; van de Rijn, M.; Botstein, D.; et al. Endothelial cell diversity revealed by global expression profiling. Proc. Natl. Acad. Sci. USA 2003, 100, 10623–10628. [Google Scholar] [CrossRef] [PubMed]
- Cleuren, A.C.A.; van der Ent, M.A.; Jiang, H.; Hunker, K.L.; Yee, A.; Siemieniak, D.R.; Molema, G.; Aird, W.C.; Ganesh, S.K.; Ginsburg, D. The in vivo endothelial cell translatome is highly heterogeneous across vascular beds. Proc. Natl. Acad. Sci. USA 2019, 116, 23618–23624. [Google Scholar] [CrossRef] [PubMed]
- Goettsch, W.; Schubert, A.; Morawietz, H. Expression of human endothelin-converting enzyme isoforms: Role of angiotensin II. Can. J. Physiol. Pharmacol. 2008, 86, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Judkins, C.P.; Diep, H.; Broughton, B.R.; Mast, A.E.; Hooker, E.U.; Miller, A.A.; Selemidis, S.; Dusting, G.J.; Sobey, C.G.; Drummond, G.R. Direct evidence of a role for Nox2 in superoxide production, reduced nitric oxide bioavailability, and early atherosclerotic plaque formation in ApoE−/− mice. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H24–H32. [Google Scholar] [CrossRef] [PubMed]
- Dikalov, S.I.; Nazarewicz, R.R.; Bikineyeva, A.; Hilenski, L.; Lassègue, B.; Griendling, K.K.; Harrison, D.G.; Dikalova, A.E. Nox2-induced production of mitochondrial superoxide in angiotensin II-mediated endothelial oxidative stress and hypertension. Antioxid. Redox Signal 2014, 20, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.; Bendall, J.K.; Crabtree, M.J.; Tatham, A.L.; Carter, E.E.; Hale, A.B.; Channon, K.M. Endothelial-specific Nox2 overexpression increases vascular superoxide and macrophage recruitment in ApoE⁻/⁻ mice. Cardiovasc. Res. 2012, 94, 20–29. [Google Scholar] [CrossRef]
- Pance, A. Oct-1, to go or not to go? That is the PolII question. Biochim. Biophys. Acta 2016, 1859, 820–824. [Google Scholar] [CrossRef]
- Hermonat, P.L.; Zhu, H.; Cao, M.; Mehta, J.L. LOX-1 transcription. Cardiovasc. Drugs Ther. 2011, 25, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, Y.; Liu, H.; Hermonat, P.L.; Mehta, J.L. Lectin-like oxidized low-density lipoprotein receptor-1 (LOX-1) transcriptional regulation by Oct-1 in human endothelial cells: Implications for atherosclerosis. Biochem. J. 2006, 393, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Goettsch, C.; Goettsch, W.; Muller, G.; Seebach, J.; Schnittler, H.J.; Morawietz, H. Nox4 overexpression activates reactive oxygen species and p38 MAPK in human endothelial cells. Biochem. Biophys. Res. Commun. 2009, 380, 355–360. [Google Scholar] [CrossRef] [PubMed]
- López-Ongil, S.; Senchak, V.; Saura, M.; Zaragoza, C.; Ames, M.; Ballermann, B.; Rodríguez-Puyol, M.; Rodríguez-Puyol, D.; Lowenstein, C.J. Superoxide regulation of endothelin-converting enzyme. J. Biol. Chem. 2000, 275, 26423–26427. [Google Scholar] [CrossRef]
- Muller, D.N.; Mullally, A.; Dechend, R.; Park, J.K.; Fiebeler, A.; Pilz, B.; Löffler, B.M.; Blum-Kaelin, D.; Masur, S.; Dehmlow, H.; et al. Endothelin-converting enzyme inhibition ameliorates angiotensin II-induced cardiac damage. Hypertension 2002, 40, 840–846. [Google Scholar] [CrossRef]





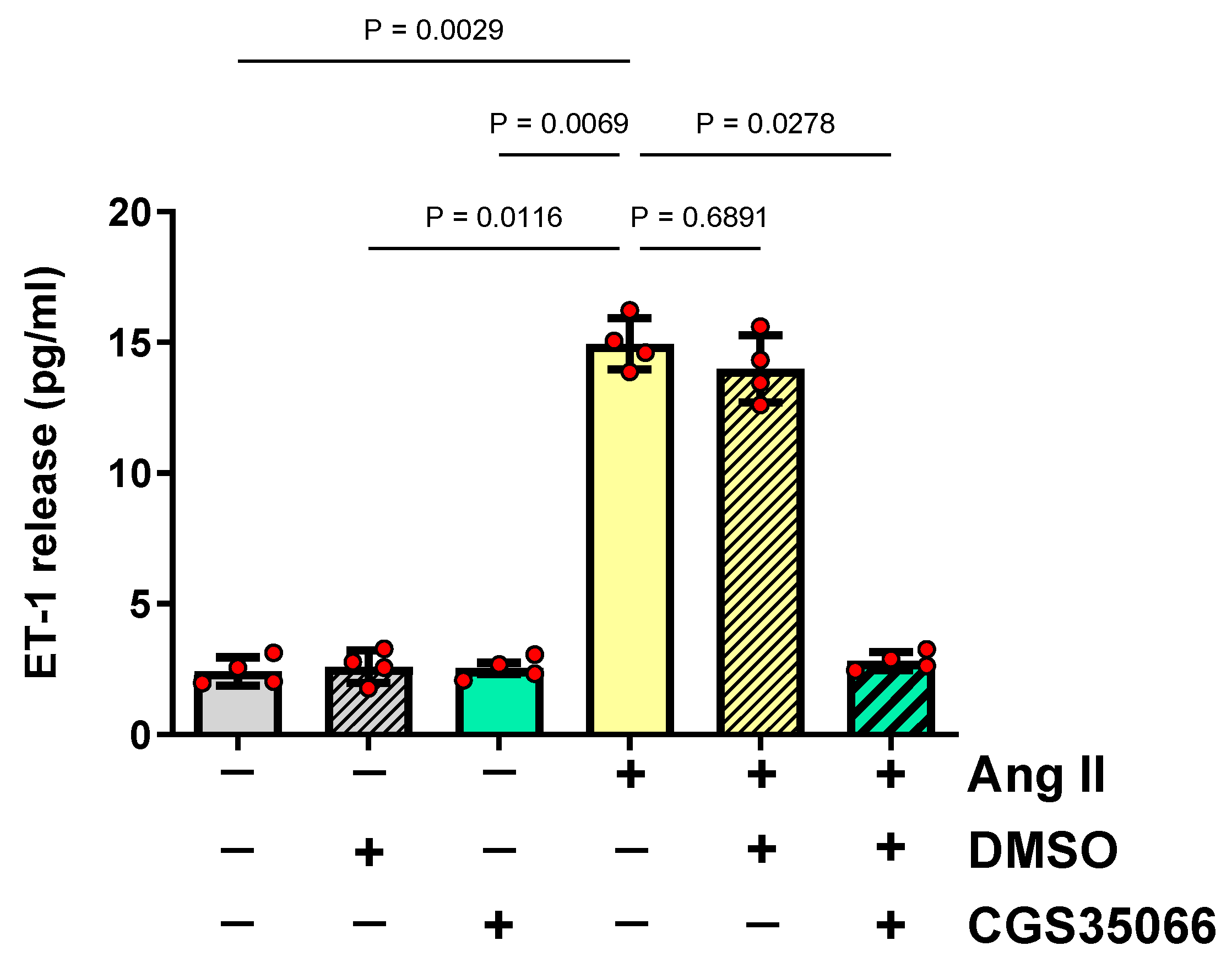{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene-ID | Sense Primer 5′→3′ | Anti-Sense Primer 5′→3′ |
|---|---|---|---|
| Mouse ECE-1 | NM_199307.1 | gCAAAACAAgCTCCTTCCTg | TggCTgATCTCCgAgTCTCT |
| Mouse GAPDH | NM_008084.2 | CATCACCATCTTCCAGGAGC | TGACCTTGCCCACAGCCTTG |
| Human ECE-1 | NM_001397.2 | CAAgCTCCTTCCTTgACCAg | gCCCAggTTgTTTTCTgTgT |
| Human GAPDH | AF261085.1 | CATCACCATCTTCCAggAgCg | TgACCTTgCCCACAgCCTTg |
| Human B2M | NM_004048.2 | GTGCTCGCGCTACTCTCTCT | CGGCAGGCATACTCATCTTT |
| pLuc 2000 | NC_000001.11 | TGGCCTAACTGGCCGGTACCACCTGGGCAAGGGTTGCAGTC | TCTTGATATCCTCGAGTGCCACCCGCGGCACCGCTGC |
| pLuc 1508 | NC_000001.11 | TGGCCTAACTGGCCGGTACCTACAACAGGGACACCACATTT | Identical sequence as pLuc 2000 above |
| pLuc 1008 | NC_000001.11 | TGGCCTAACTGGCCGGTACCACAGACACACGGCAACAAACC | Identical sequence as pLuc 2000 above |
| pLuc 608 | NC_000001.11 | TGGCCTAACTGGCCGGTACCCCTCCCACCGTTTCTGTCTCC | Identical sequence as pLuc 2000 above |
| pLuc 108 | NC_000001.11 | TGGCCTAACTGGCCGGTACCAGGCAGCCGAGCCGTCCGAGC | Identical sequence as pLuc 2000 above |
| EMSA | NC_000001.11 | CAAATCCCAAATATAGTCAGGACT | AGTCCTGACTATATTTGGGATTTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adu-Gyamfi, M.; Goettsch, C.; Kamhieh-Milz, J.; Chen, L.; Pfefferkorn, A.M.; Hofmann, A.; Brunssen, C.; Müller, G.; Walther, T.; Ashraf, M.I.; et al. The Role of NOX2-Derived Reactive Oxygen Species in the Induction of Endothelin-Converting Enzyme-1 by Angiotensin II. Antioxidants 2024, 13, 500. https://doi.org/10.3390/antiox13040500
Adu-Gyamfi M, Goettsch C, Kamhieh-Milz J, Chen L, Pfefferkorn AM, Hofmann A, Brunssen C, Müller G, Walther T, Ashraf MI, et al. The Role of NOX2-Derived Reactive Oxygen Species in the Induction of Endothelin-Converting Enzyme-1 by Angiotensin II. Antioxidants. 2024; 13(4):500. https://doi.org/10.3390/antiox13040500
Chicago/Turabian StyleAdu-Gyamfi, Michael, Claudia Goettsch, Julian Kamhieh-Milz, Lei Chen, Anna Maria Pfefferkorn, Anja Hofmann, Coy Brunssen, Gregor Müller, Thomas Walther, Muhammad Imtiaz Ashraf, and et al. 2024. "The Role of NOX2-Derived Reactive Oxygen Species in the Induction of Endothelin-Converting Enzyme-1 by Angiotensin II" Antioxidants 13, no. 4: 500. https://doi.org/10.3390/antiox13040500
APA StyleAdu-Gyamfi, M., Goettsch, C., Kamhieh-Milz, J., Chen, L., Pfefferkorn, A. M., Hofmann, A., Brunssen, C., Müller, G., Walther, T., Ashraf, M. I., Morawietz, H., Witowski, J., & Catar, R. (2024). The Role of NOX2-Derived Reactive Oxygen Species in the Induction of Endothelin-Converting Enzyme-1 by Angiotensin II. Antioxidants, 13(4), 500. https://doi.org/10.3390/antiox13040500