
Differences in Airway Remodeling and Emphysematous Lesions between Rats Exposed to Smoke from New-Type and Conventional Tobacco Varieties

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tested Cigarettes
2.2. Chemicals and Reagents
2.3. Experimental Animals
2.4. Model Establishment and Sampling
2.5. Total and Differential Cell Counts
2.6. Histological Examination
2.7. TUNEL Analysis
2.8. Oxidative Stress Assay
2.9. ELISA Assay
2.10. Immunohistochemical Analysis
2.11. Statistical Analysis
3. Results
3.1. Observation of General Condition
3.2. New-Type Tobacco Caused Less Pathological Damage to Lung Tissues than “HHY”
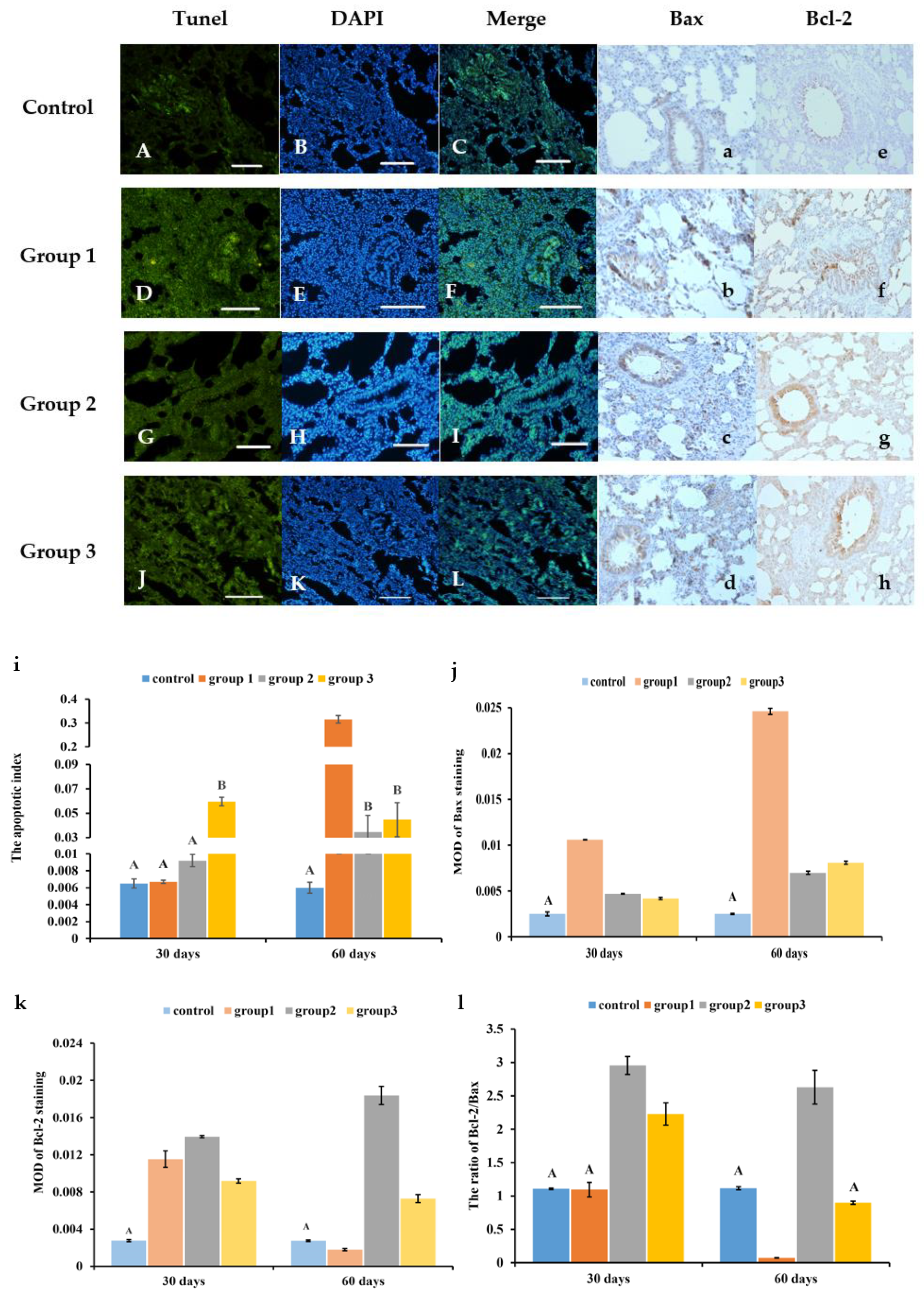
3.3. New-Type Tobacco Caused Less Apoptosis in Lung Tissues than “HHY”
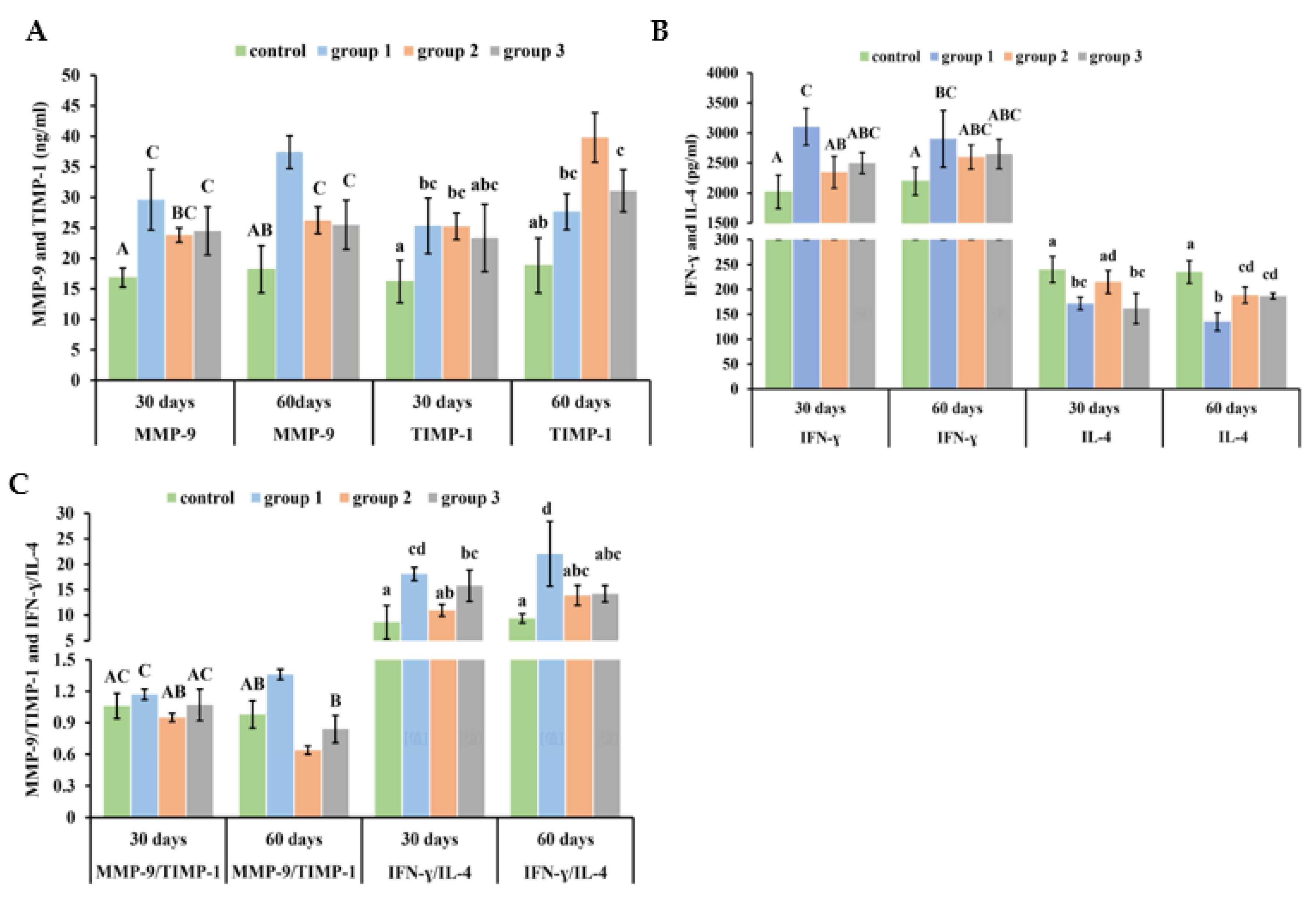
3.4. New-Type Tobacco Caused Less of an Increase in the Numbers of Total Leukocytes and Neutrophils in the BALF than “HHY”
3.5. New-Type Tobacco Caused Less Oxidative Stress in Lung Tissues than “HHY”
3.6. New-Type Tobacco Altered the Ratios of MMP-9/TIMP-1 and IFN-γ/IL-4 in Lung Tissue to a Lesser Degree than “HHY”
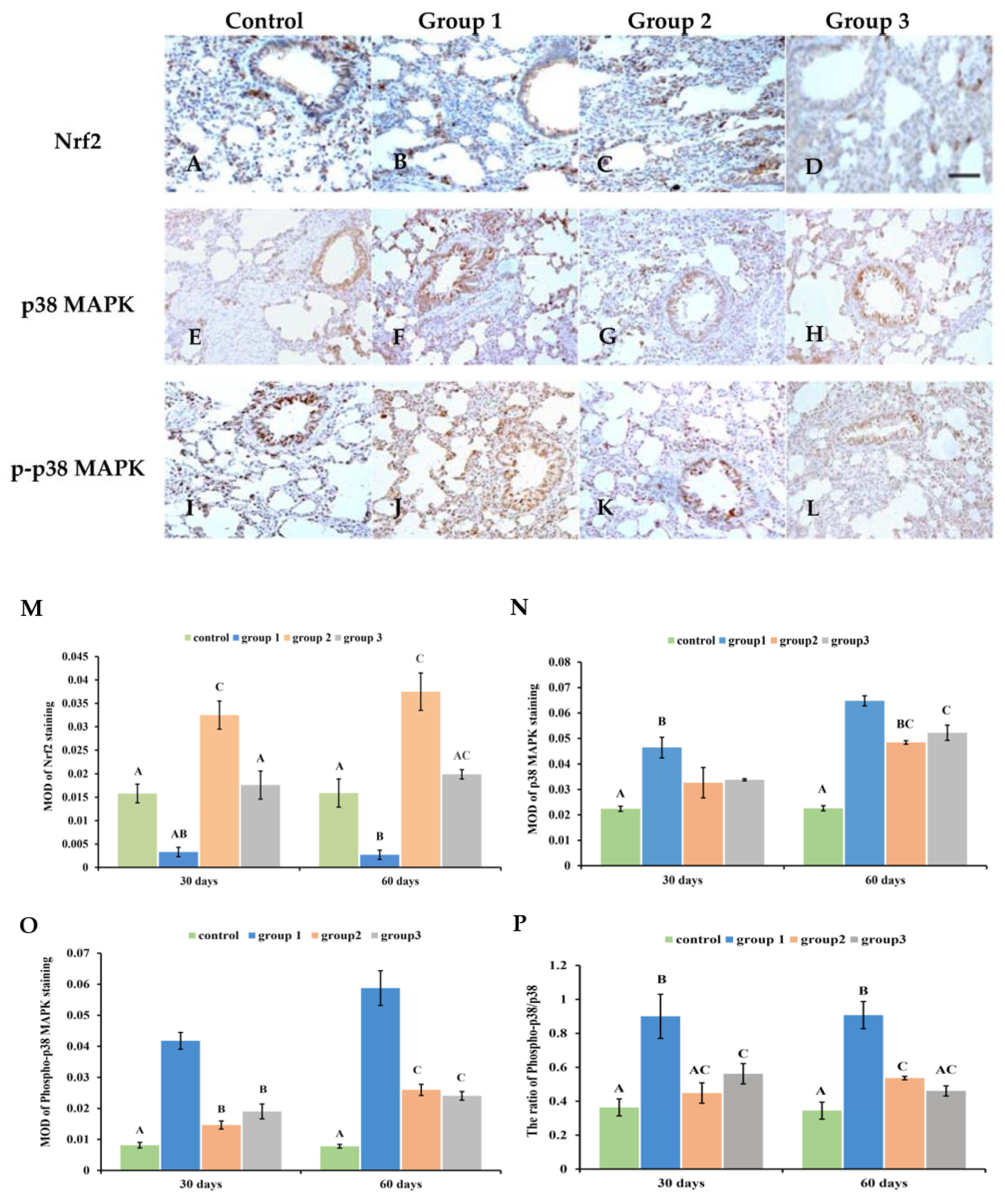
3.7. New-Type Tobacco Altered the Expression of Nrf2 and p38MAPK
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dai, X.C.; Gakidou, E.; Lopez, A.D. Evolution of the global smoking epidemic over the past half century: Strengthening the evidence base for policy action. Tob. Control 2022, 31, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, X.; Gao, X.; Lv, Y.; Zhou, L.; Shi, J.; Wei, W.; Huang, J.; Deng, L.; Wang, Z.; et al. Epidemiological evidence relating risk factors to chronic obstructive pulmonary disease in China: A systematic review and meta-analysis. PLoS ONE 2021, 16, e0261692. [Google Scholar] [CrossRef] [PubMed]
- Albano, G.D.; Gagliardo, R.P.; Montalbano, A.M.; Mirella, P. Overview of the mechanisms of oxidative stress: Impact in inflammation of the airway diseases. Antioxidants 2022, 11, 2237. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.F. Multifaceted mechanisms in COPD: Inflammation, immunity, and tissue repair and destruction. Eur. Respir. J. 2008, 31, 1334–1356. [Google Scholar] [CrossRef] [PubMed]
- Gan, Q.; Yang, J.; Yang, G.H.; Goniewicz, M.; Benowitz, N.L.; Glantz, S.A. Chinese “herbal” cigarettes are as carcinogenic and addictive as regular cigarettes. Cancer Epidem. Biomar. 2009, 18, 3497–3501. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, P.B.; Romeiro, N.C. Multi-target natural products as alternatives against oxidative stress in Chronic Obstructive Pulmonary Disease (COPD). Eur. J. Med. Chem. 2019, 163, 911–931. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.G.; Dai, Y.; Li, D.L.; Ma, K.Y. Ginkgo biloba leaf extract action in scavenging free radicals and reducing mutagenicity and toxicity of cigarette smoke in vivo. J. Environ. Sci. Health A Tox. Hazard. Subst. Environ. Eng. 2010, 45, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Carmines, E.L.; Lemus, R.; Gaworski, C.L. Toxicologic evaluation of licorice extract as a cigarette ingredient. Food Chem. Toxicol. 2005, 43, 1303–1322. [Google Scholar] [CrossRef] [PubMed]
- Thielen, A.; Klus, H.; Müller, L. Tobacco smoke: Unraveling a controversial subject. Exp. Toxicol. Pathol. 2008, 60, 141–156. [Google Scholar] [CrossRef]
- Vanscheeuwijck, P.M.; Teredesai, A.; Terpstra, P.M.; Verbeeck, J.; Kuhl, P.; Gerstenberg, B.; Gebel, S.; Carmines, E.L. Evaluation of the potential effects of ingredients added to cigarettes. Part 4: Subchronic inhalation toxicity. Food Chem. Toxicol. 2002, 40, 113–131. [Google Scholar] [CrossRef]
- Andreoli, C.; Gigante, D.; Nunziata, A. A review of in vitro methods to assess the biological activity of tobacco smoke with the aim of reducing the toxicity of smoke. Toxicol. Vitr. 2003, 17, 587–594. [Google Scholar] [CrossRef]
- Borgerding, M.; Klus, H. Analysis of complex mixtures-cigarette smoke. Exp. Toxicol. Pathol. 2005, 57, 43–73. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.S.; Nicholson, J.S. Aspects of the evolution of Nicotiana tabacum L. and the status of the United States Nicotiana Germplasm Collection. Genet. Resour. Crop Evol. 2007, 54, 727–740. [Google Scholar] [CrossRef]
- Wei, K.Q.; Yang, J.X.; Wei, Z.Z. Pollination following grafting introduces efficiently Ocimum basilicum L. genes into Nicotiana tabacum L. Span. J. Agric. Res. 2013, 11, 1068–1077. [Google Scholar] [CrossRef]
- Wei, K.Q.; Pang, S.X.; Yang, J.X.; Wei, Z.Z. Enhancement of cadmium tolerance and accumulation by introducing Perilla frutescens (L.) Britt var. frutescens genes in Nicotiana tabacum L. plants. Environ. Sci. Pollut. Res. 2015, 22, 5405–5416. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.Z.; Wei, K.Q. Distant Hybridization Breeding of Tobacco; China Agricultural Science and Technology Press: Beijing, China, 2008. [Google Scholar]
- Li, J.J. Metabolomics Study on Cigarette Smoke Exposure-Induced COPD Based on 1H-NMR. Master’s Thesis, Shanxi University, Taiyuan, China, 2019. [Google Scholar]
- Wei, Z.Z.; Wei, K.Q.; Yang, J.W. Distant Hybridization Breeding of Tobacco, 2nd ed.; China Agricultural Science and Technology Press: Beijing, China, 2019. [Google Scholar]
- Wei, K.Q.; Zhang, X.; Yang, J.W.; Chen, J.Y. Tobacco introduced Perilla frutescens and Ocimum basilicum genes attenuates neutrophilic inflammation in lung tissues of COPD rats. Ecotoxicol. Environ. Saf. 2024, 271, 115956. [Google Scholar] [CrossRef]
- Carmines, E.L. Evaluation of the potential effects of ingredients added to cigarettes. Part 1: Cigarette design, testing approach, and review of results. Food Chem. Toxicol. 2002, 40, 77–91. [Google Scholar] [CrossRef]
- Mattes, W.B.; Richter, P.; Mendrick, D.L. An omics strategy for discovering pulmonary biomarkers potentially relevant to the evaluation of tobacco products. Biomark. Med. 2012, 6, 849–860. [Google Scholar]
- Roemer, E.; Tewes, F.J.; Meisgen, T.J.; Veltel, D.J.; Carmines, E.L. Evaluation of the potential effects of ingredients added to cigarettes. Part 3: In vitro genotoxicity and cytotoxicity. Food Chem. Toxicol. 2002, 40, 105–111. [Google Scholar] [CrossRef]
- Ghorani, V.; Boskabady, M.H.; Khazdair, M.R.; Kianmeher, M. Experimental animal models for COPD: A methodological review. Tob. Induc. Dis. 2017, 15, 25. [Google Scholar] [CrossRef]
- John, L.G.; Kohse, K.; Orasche, J.; Reda, A.; Schnelle-Kreis, J.; Zimmermann, R.; Schmid, O.; Eickelberg, O.; Yildirim, A.O. The composition of cigarette smoke determines inflammatory cell recruitment to the lung in COPD mouse models. Clin. Sci. 2014, 126, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.; Donovan, C.; Liu, G.; Gomez, H.M.; Chimankar, V.; Harrison, C.L.; Wiegman, C.H.; Adcock, I.M.; Knight, D.A.; Hirota, J.A.; et al. Animal models of COPD: What do they tell us? Respirology 2017, 22, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Che, H.Z.; Lin, J.T.; Ding, A.J.; Zhang, Q. Research directions and keywords under the secondary application codes of the atmospheric sciences discipline of the National Natural Science Foundation of China: D0514 Atmospheric environment and health meteorology. Chin. J. Atmos. Sci. 2023, 47, 220–229. [Google Scholar]
- Wang, Z.D.; Zhang, X.W.; Liu, Y.H. Atlas of Chinese Tobacco Core Collection; Scientific and Technical Documentation Press: Beijing, China, 2014. [Google Scholar]
- Zheng, H.G.; Liu, Y.N.; Huang, T.; Fang, Z.M.; Li, G.S.; He, S.H. Development and characterization of a rat model of chronic obstructive pulmonary disease (COPD) induced by sidestream cigarette smoke. Toxicol. Lett. 2009, 189, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.F.; Mohammadtursun, N.; Sun, J.; Lv, Y.B.; Jin, H.L.; Lin, J.P.; Kong, L.W.; Zhao, Z.X.; Zhang, H.Y.; Dong, J.C. Establishment and evaluation of a rat model of sidestream cigarette smoke-induced chronic obstructive pulmonary disease. Front. Physiol. 2018, 9, 58. [Google Scholar] [CrossRef]
- Jiao, P.F.; Wang, Y.R.; Sang, T.Q.; Jiao, J.; Li, Y.M. Molecular mechanism of betulin palliative therapy for chronic obstructive pulmonary disease (COPD) based on P2X7 receptor target of gated ion channel. Ann. Transl. Med. 2022, 10, 707. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Lv, Z.Q.; Jiang, S.P. The effects of triptolide on airway remodelling and transforming growth factor-β1/Smad signalling pathway in ovalbumin-sensitized mice. Immunology 2011, 132, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Sheng, H.Y.; Zhang, Y.J.; Shi, X.Q.; Hu, Y.H.; Pang, B.S.; Jin, J.W.; Ma, Y.M. Functional, ultrastructural, and transcriptomic changes in rat diaphragms with different durations of cigarette smoke exposure. Int. J. Chronic Obstr. 2020, 15, 3135–3145. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Wang, P.P.; Wang, X.M.; He, Y.J.; Yao, W.Z.; Qi, Y.F.; Tang, C.S. Involvement of endogenous hydrogen sulfide in cigarette smoke-induced changes in airway responsiveness and inflammation of rat lung. Cytokine 2011, 53, 334–341. [Google Scholar] [CrossRef]
- Bhalla, D.K.; Hirata, F.; Rishi, A.K.; Gairola, C.G. Cigarette smoke, inflammation, and lung Injury: A mechanistic perspective. J. Toxicol. Environ. Health Part B 2009, 12, 45–64. [Google Scholar] [CrossRef]
- Liu, C.; DeGrandpre, Y.; Griffiths, A.; McAdam, K.; Voisine, R.; Cote, F.; Proctor, C. The use of a novel tobacco treatment process to reduce toxicant yields in cigarette smoke. Food Chem. Toxicol. 2011, 49, 1904–1917. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, D.; Ursini, C.L.; Fresegna, A.M.; Maiello, R.; Ciervo, A.; Ferrante, R.; Buresti, G.; Iavicoli, S. Cyto-genotoxic effects of smoke from commercial filter and non-filter cigarettes on human bronchial and pulmonary cells. Mutat. Res. Toxicol. Environ. Mutagen. 2013, 750, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rustemeier, K.; Stabbert, R.; Haussmann, H.J.; Roemer, E.; Carmines, E.L. Evaluation of the potential effects of ingredients added to cigarettes. Part 2: Chemical composition of mainstream smoke. Food Chem. Toxicol. 2002, 40, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, R.; Coggins, C.R.E.; Roemer, E. Toxicological assessment of cigarette ingredients. Regul. Toxicol. Pharm. 2011, 61, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Bombick, D.W.; Putnam, K.; Doolittle, D.J. Comparative cytotoxicity studies of smoke condensates from different types of cigarettes and tobaccos. Toxicol. Vitr. 1998, 12, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Richter, P.A.; Li, A.P.; Polzin, G.; Roy, S.K. Cytotoxicity of eight cigarette smoke condensates in three test systems: Comparisons between assays and condensates. Regul. Toxicol. Pharm. 2010, 58, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Horvath, G.; Acs, K. Essential oils in the treatment of respiratory tract diseases highlighting their role in bacterial infections and their anti-inflammatory action: A review. Flavour Frag. J. 2015, 3, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Combes, R.; Scott, K.; Dillon, D.; Meredith, C.; McAdam, K.; Proctor, C. The effect of a novel tobacco process on the in vitro cytotoxicity and genotoxicity of cigarette smoke particulate matter. Toxicol. Vitr. 2012, 26, 1022–1029. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J.; Sohn, H.O.; Han, J.H.; Park, C.H.; Lee, H.S.; Lee, D.W.; Hwang, K.J.; Hyun, H.C. Effect of cigarette filters on the chemical composition and in vitro biological activity of cigarette mainstream smoke. Food Chem. Toxicol. 2009, 47, 192–197. [Google Scholar] [CrossRef]
- Ram, A.; Balachandar, S.; Vijayananth, P.; Singh, V.P. Medicinal plants useful for treating chronic obstructive pulmonary disease (COPD): Current status and future perspectives. Fitoterapia 2011, 82, 141–151. [Google Scholar] [CrossRef]
- Tanner, L.; Single, A.B. Animal models reflecting chronic obstructive pulmonary disease and related respiratory disorders: Translating pre-clinical data into clinical relevance. J. Innate Immun. 2020, 12, 203–225. [Google Scholar] [CrossRef] [PubMed]
- Haustein, K.O. Tobacco Constituents and Additives. In Tobacco or Health? Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Sparks, N.R.L.; Walker, L.M.; Sera, S.R.; Madrid, J.V.; Hanna, M.; Dominguez, E.C.; zur Nieden, N.I. Sidestream smoke extracts from harm-reduction and conventional camel cigarettes inhibit osteogenic differentiation via oxidative stress and differential activation of intrinsic apoptotic pathways. Antioxidants 2022, 11, 2474. [Google Scholar] [CrossRef] [PubMed]
- Cosio, M.; Ghezzo, H.; Hogg, J.C.; Corbin, R.; Loveland, M.; Dosman, J.; Macklem, P.T. The relations between structural changes in small airways and pulmonary-function tests. N. Engl. J. Med. 1978, 298, 1277–1281. [Google Scholar] [CrossRef] [PubMed]
- Aminian, A.R.; Mohebbati, R.; Boskabady, M.H. The effect of Ocimum basilicum L. and its main ingredients on respiratory disorders: An experimental, preclinical, and clinical review. Front. Pharmacol. 2022, 12, 805391. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.Q.; Li, X.Y.; Fang, N.; Li, P.; Zhang, Z.Q.; Lin, M.B.; Hou, Q. Perilla leaf extract (PLE) attenuates COPD airway inflammation via the TLR4/Syk/PKC/NF-κB pathway in vivo and in vitro. Front. Pharmacol. 2022, 12, 763624. [Google Scholar] [CrossRef] [PubMed]
- Zekonis, G.; Zekonis, J.; Sadzeviciene, R.; Simoniene, G.; Kevelaitis, E. Effect of Perilla frutescens aqueous extract on free radical production by human neutrophil leukocytes. Medicina 2008, 44, 699. [Google Scholar] [CrossRef]
- Barnes, P.J. Oxidative stress in chronic obstructive pulmonary disease. Antioxidants 2022, 11, 965. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I. Oxidative stress in pathogenesis of chronic obstructive pulmonary disease. Cell Biochem. Bioph. 2005, 43, 167–188. [Google Scholar] [CrossRef] [PubMed]
- Abdella, A.M.; Attia, G.A.; Eed, M.A.; Eldib, A.S.; Haleem, S.S. Evaluation of matrix metalloproteinase-9 and tissue inhibitor metalloproteinase-1 levels in bronchoalveolar lavage of apparently healthy smokers. Egypt. J. Chest Dis. Tuberc. 2015, 64, 371–378. [Google Scholar] [CrossRef]
- MacNee, W. Pathogenesis of chronic obstructive pulmonary disease. Clin. Chest Med. 2007, 28, 479–513. [Google Scholar] [CrossRef]
- Rangasamy, T.; Misra, V.; Zhen, L.J.; Tankersley, C.G.; Tuder, R.M.; Biswal, S. Cigarette smoke-induced emphysema in A/J mice is associated with pulmonary oxidative stress, apoptosis of lung cells, and global alterations in gene expression. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, 888–900. [Google Scholar] [CrossRef] [PubMed]
- Sadowska, A.M.; van Overveld, F.J.; Gorecka, D.; Zdral, A.; Filewska, M.; Demkow, U.A.; Luyten, C.; Saenen, E.; Zielinski, J.; De Backer, W.A. The interrelationship between markers of inflammation and oxidative stress in chronic obstructive pulmonary disease: Modulation by inhaled steroids and antioxidant. Resp. Med. 2005, 99, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Demedts, I.K.; Demoor, T.; Bracke, K.R.; Joos, G.F.; Brusselle, G.G. Role of apoptosis in the pathogenesis of COPD and pulmonary emphysema. Resp. Res. 2006, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, P.; Rahman, I. Oxidative stress in asthma and COPD: Antioxidants as a therapeutic strategy. Pharmacol. Therapeut. 2006, 111, 476–494. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Fu, C.; Liu, J.; Sai, X.; Qin, C.; Di, T.; Yang, Y.; Wu, Y.; Bian, T. Hypermethylation of the Nrf2 promoter induces ferroptosis by inhibiting the Nrf2-GPX4 axis in COPD. Int. J. Chronic. Obstr. 2021, 16, 3347–3362. [Google Scholar] [CrossRef]
- Kubo, H.; Asai, K.; Kojima, K.; Sugitani, A.; Kyomoto, Y.; Okamoto, A.; Yamada, K.; Ijiri, N.; Watanabe, T.; Hirata, K.; et al. Astaxanthin suppresses cigarette smoke-induced emphysema through Nrf2 activation in mice. Mar. Drugs 2019, 17, 673. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, K.; Li, Y.; Du, B.; Wu, J. Differences in Airway Remodeling and Emphysematous Lesions between Rats Exposed to Smoke from New-Type and Conventional Tobacco Varieties. Antioxidants 2024, 13, 511. https://doi.org/10.3390/antiox13050511
Wei K, Li Y, Du B, Wu J. Differences in Airway Remodeling and Emphysematous Lesions between Rats Exposed to Smoke from New-Type and Conventional Tobacco Varieties. Antioxidants. 2024; 13(5):511. https://doi.org/10.3390/antiox13050511
Chicago/Turabian StyleWei, Keqiang, Yuanyuan Li, Bin Du, and Juan Wu. 2024. "Differences in Airway Remodeling and Emphysematous Lesions between Rats Exposed to Smoke from New-Type and Conventional Tobacco Varieties" Antioxidants 13, no. 5: 511. https://doi.org/10.3390/antiox13050511
APA StyleWei, K., Li, Y., Du, B., & Wu, J. (2024). Differences in Airway Remodeling and Emphysematous Lesions between Rats Exposed to Smoke from New-Type and Conventional Tobacco Varieties. Antioxidants, 13(5), 511. https://doi.org/10.3390/antiox13050511