
Dietary Astaxanthin Can Promote the Growth and Motivate Lipid Metabolism by Improving Antioxidant Properties for Swimming Crab, Portunus trituberculatus
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Diets Preparation
2.2. Sample Collection
2.3. Fatty Acids Analysis in Hepatopancreas
2.4. Antioxidant and Non-Specific Immune-Related Enzyme Activities
2.5. Lipid Metabolism-Related Enzyme Activities and Crude Lipid Content of Hepatopancreatic and Muscle
2.6. Tissue Section and Observation
2.7. Quantitative RT-PCR
2.8. Statistical Analysis
3. Results
3.1. Color Parameters
3.2. Growth Performance and Survival
3.3. Composition of Fatty Acids in the Hepatopancreas
3.4. Enzymes Activities of Non-Specific Immunity and Antioxidant in Hemolymph
3.5. The Activities of Enzymes Related to Lipid Metabolism in Hepatopancreas and Oil Red Staining
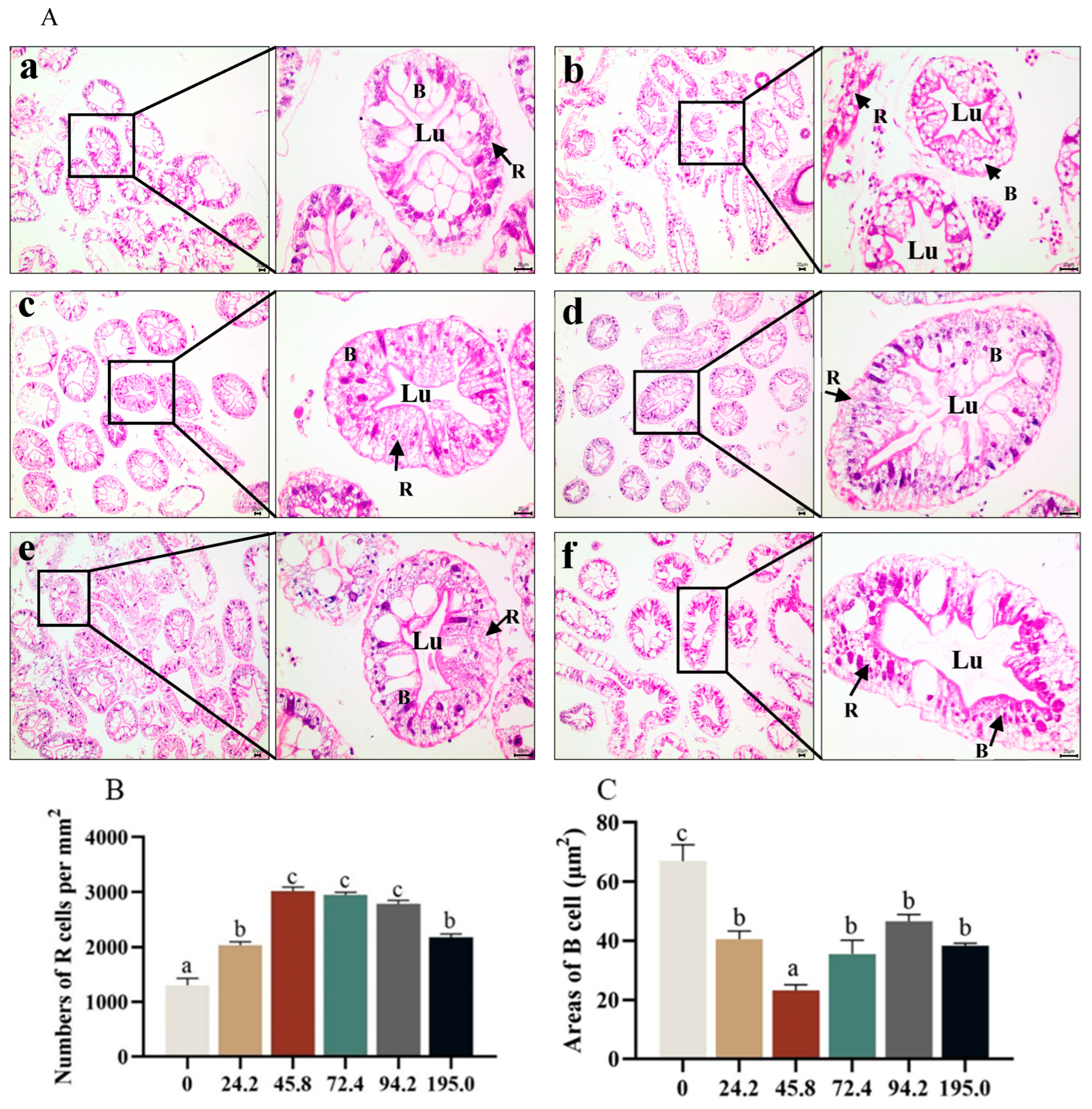
3.6. Histological Structure in Hepatopancreas
3.7. The Expression of Genes Related to Immune-Related in Intestinal
3.8. Expression of Genes Related to Antioxidant and Fatty Acid Metabolism in Hepatopancreas
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, W.; Liu, J. Astaxanthin isomers: Selective distribution and isomerization in aquatic animals. Aquaculture 2020, 520, 734915. [Google Scholar] [CrossRef]
- Li, B.; Lee, J.Y.; Luo, Y. Health benefits of astaxanthin and its encapsulation for improving bioavailability: A review. J. Agric. Food Res. 2023, 14, 100685. [Google Scholar] [CrossRef]
- Najoan, G.C.; Prasetyaningsih, A.; Prakasita, V.C.; Wisaksono, A.A.; Rahardjo, D. Anti-inflammatory activity test of astaxan thin extract from Litopenaeus vannamei shrimp waste against the number of neutrophils and lymphocytes in white rats (Rattus norvegicus) injected with carrageenin. Sch. Acad. Biosci. 2021, 9, 123–129. [Google Scholar] [CrossRef]
- McGraw, K.J.; Toomey, M.B. Carotenoid accumulation in the tissues of zebra finches: Predictors of integument tary pigmentation and implications for carotenoid allocation strategies. Physiol. Biochem. Zool. PBZ 2010, 83, 97–109. [Google Scholar] [CrossRef]
- Yao, Q.; Ma, J.; Chen, X.; Zhao, G.; Zang, J. A natural strategy for astaxanthin stabilization and color regulation: Interaction with proteins. Food Chem. 2023, 402, 134343. [Google Scholar] [CrossRef] [PubMed]
- Gowd, V.; Xiao, J.; Wang, M.; Chen, F.; Cheng, K. Multi-mechanistic antidiabetic potential of astaxanthin: An update on preclinical and clinical evidence. Mol. Nutr. Food Res. 2021, 65, 2100252. [Google Scholar] [CrossRef]
- Wang, S.; Qi, X. The putative role of astaxanthin in neuroinflammation modulation: Mechanisms and therapeutic potential. Front. Pharmacol. 2022, 13, 916653. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Wang, M.; Hu, S.; Feng, Y.; Shao, Y.; Xie, Y.; Wu, M.; Chen, Y.; Shi, X. The neuroprotective effect of astaxanthin on pilocarpine-Induced status epilepticus in rats. Front. Cell Neurosci. 2019, 13, 123. [Google Scholar] [CrossRef]
- Niu, T.; Xuan, R.; Jiang, L.; Wu, W.; Zhen, Z.; Song, Y.; Hong, L.; Zheng, K.; Zhang, J.; Xu, Q.; et al. Astaxanthin induces the Nrf2/HO–1 antioxidant pathway in human umbilical vein endothelial cells by generating trace amounts of ROS. J. Agric. Food Chem. 2018, 66, 1551–1559. [Google Scholar] [CrossRef]
- Zarneshan, S.N.; Fakhri, S.; Farzaei, M.H.; Khan, H.; Saso, L. Astaxanthin targets PI3K/Akt signaling pathway toward po tential therapeutic applications. Food Chem. Toxicol. 2020, 145, 111714. [Google Scholar] [CrossRef]
- Long, X.; Wu, X.; Zhao, L.; Liu, J.; Cheng, Y. Effects of dietary supplementation with Haematococcus pluvialis cell powder on coloration, ovarian development and antioxidation capacity of adult female Chinese mitten crab, Eriocheir sinensis. Aquaculture 2017, 473, 545–553. [Google Scholar] [CrossRef]
- Ma, N.; Long, X.; Liu, J.; Chang, G.; Deng, D.; Cheng, Y.; Wu, X. Defatted Haematococcus pluvialis meal can enhance the coloration of adult Chinese mitten crab Eriocheir sinensis. Aquaculture 2019, 510, 371–379. [Google Scholar] [CrossRef]
- Wu, X.; Zhao, L.; Long, X.; Liu, J.; Su, F.; Cheng, Y. Effects of dietary supplementation of Haematococcus pluvialis powder on gonadal development, coloration and antioxidant capacity of adult male Chinese mitten crab (Eriocheir sinensis). Aquac. Res. 2017, 48, 5214–5223. [Google Scholar] [CrossRef]
- Zhang, Y.; Qian, C.; Huang, J.; Li, J.; Jiang, X.; Li, Z.; Cheng, Y.; Li, J. Suitable natural astaxanthin supplementation with Haematococcus pluvialis improves the physiological function and stress response to air exposure of juvenile red swamp crayfish (Procambarus clarkii). Aquaculture 2023, 573, 739577. [Google Scholar] [CrossRef]
- Jiang, X.; Zu, L.; Wang, Z.; Cheng, Y.; Yang, Y.; Wu, X. Micro-algal astaxanthin could improve the antioxidant capability, immunity and ammonia resistance of juvenile Chinese mitten crab, Eriocheir sinensis. Fish. Shellfish Immun. 2020, 102, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, J.; Cao, J.; Liu, P.; Li, J.; Meng, X. Long-term ammonia toxicity in the hepatopancreas of swimming crab Portunus trituberculatus: Cellular stress response and tissue damage. Front. Mar. Sci. 2022, 8, 757602. [Google Scholar] [CrossRef]
- Tsai, M.C.; Huang, S.C.; Chang, W.T.; Chen, S.C.; Hsu, C.L. Effect of astaxanthin on the inhibition of lipid accumulation in 3T3-L1 adipocytes via modulation of lipogenesis and fatty acid transport Pathways. Molecules 2020, 25, 3598. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Jin, M.; Jiao, L.; Monroig, Ó.; Navarro, J.C.; Tocher, D.R.; Betancor, M.B.; Wang, X.; Yuan, Y.; Zhou, Q. Effects of dietary lipid level on growth, fatty acid profiles, antioxidant capacity and expression of genes involved in lipid metabolism in juvenile swimming crab, Portunus trituberculatus. Br. J. Nutr. 2020, 123, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Yin, P.; Tian, L.; Yu, Y.; Liu, Y.; Niu, J. Dietary supplementation of astaxanthin improved the growth performance, antioxidant ability and immune response of juvenile largemouth bass (Micropterus salmoides) fed high-fat diet. Marine Drugs 2020, 18, 642. [Google Scholar] [CrossRef]
- Liao, Z.; Xu, H.; Wei, Y.; Zhang, Q.; Liang, M. Dietary astaxanthin differentially affected the lipid accumulation in the liver and muscle of the marine teleost, tiger puffer Takifugu rubripes. Aquac. Res. 2018, 49, 3421–3433. [Google Scholar] [CrossRef]
- Ritola, O.; Tossavainen, K.; Kiuru, T.; Lindstrom-Seppa, P.; Molsa, H. Effects of continuous and episodic hyperoxia on stress and hepatic glutathione levels in one-summer-old rainbow trout (Oncorhynchus mykiss). J. Appl. Ichthyol. 2002, 18, 159–164. [Google Scholar] [CrossRef]
- Xie, S.; Li, X.; Yang, Y.; Guo, C.; Zhang, X.; Zhu, T.; Luo, J.; Yang, Z.; Zhao, W.; Cui, Y.; et al. Effects of dietary isoleucine level on growth and expression of genes related to nutritional and physiological metabolism of swimming crab (Portunus trituberculatus). Aquaculture 2023, 574, 739700. [Google Scholar] [CrossRef]
- Luo, J.; Zhu, T.; Wang, X.; Cheng, X.; Yuan, Y.; Jin, M.; Betancor, M.B.; Tocher, D.R.; Zhou, Q. Toxicological mechanism of excessive copper supplementation: Effects on coloration, copper bioaccumulation and oxidation resistance in mud crab Scylla paramamosain. J. Hazard. Mater. 2020, 395, 122600. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Monroig, Ó.; Lu, Y.; Yuan, Y.; Li, Y.; Ding, L.; Tocher, D.R.; Zhou, Q. Dietary DHA/EPA ratio affected tissue fatty acid profiles, antioxidant capacity, hematological characteristics and expression of lipid-related genes but not growth in juvenile black seabream (Acanthopagrus schlegelii). PLoS ONE 2017, 4, e0176216. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemists Official (AOAC). Methods of Analysis, 16th ed.; AOAC: Arlington, VA, USA, 1995. [Google Scholar]
- Yang, Z.; Guo, C.; Xie, S.; Zhang, Y.; Zhu, T.; Zhao, W.; Luo, J.; Jin, M.; Zhou, Q. Interactive effects of dietary cholesterol and phospholipids on growth and metabolism of juvenile swimming crab, Portunus trituberculatus. Anim. Feed. Sci. Technol. 2022, 294, 115484. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Su, F.; Yu, W.; Liu, J. Comparison of effect of dietary supplementation with Haematococcus pluvialis powder and synthetic astaxanthin on carotenoid composition, concentration, esterification degree and astaxanthin isomers in ovaries, hepatopancreas, carapace, epithelium of adult female Chinese mitten crab (Eriocheir sinensis). Aquaculture 2020, 523, 735146. [Google Scholar] [CrossRef]
- Lim, K.C.; Yusoff, F.M.; Shariff, M.; Kamarudin, M.S. Astaxanthin as feed supplement in aquatic animals. Rev. Aquac. 2018, 10, 738–773. [Google Scholar] [CrossRef]
- Chien, Y.H.; Pan, C.H.; Hunter, B. The resistance to physical stresses by Penaeus monodon juveniles fed diets supplemented with astaxanthin. Aquaculture 2003, 216, 177–191. [Google Scholar] [CrossRef]
- Han, T.; Li, X.; Wang, J.; Wang, C.; Yang, M.; Zheng, P. Effects of dietary astaxanthin (AX) supplementation on pigmentation, antioxidant capacity and nutritional value of swimming crab, Portunus trituberculatus. Aquaculture 2018, 490, 169–177. [Google Scholar] [CrossRef]
- Wade, N.M.; Cheers, S.; Bourne, N.; Irvin, S.; Blyth, D.; Glencross, B.D. Dietary astaxanthin levels affect colour, growth, carotenoid digestibility and the accumulation of specific carotenoid esters in the giant tiger shrimp, Penaeus monodon. Aquac. Res. 2017, 48, 395–406. [Google Scholar] [CrossRef]
- Daly, B.; Swingle, J.S.; Eckert, G.L. Dietary astaxanthin supplementation for hatchery-cultured red king crab, Paralithodes camtschaticus, juveniles. Aquac. Nutr. 2013, 19, 312–320. [Google Scholar] [CrossRef]
- Doolan, B.J.; Booth, M.A.; Allan, G.L.; Jones, P.L. Effects of dietary astaxanthin concentration and feeding period on the skin pigmentation of Australian snapper Pagrus auratus (Bloch & Schneider, 1801). Aquac. Res 2008, 40, 60–68. [Google Scholar] [CrossRef]
- Yi, X.; Xu, W.; Zhou, H.; Zhang, Y.; Luo, Y.; Zhang, W.; Mai, K. Effects of dietary astaxanthin and xanthophylls on the growth and skin pigmentation of large yellow croaker Larimichthys croceus. Aquaculture 2014, 433, 377–383. [Google Scholar] [CrossRef]
- Naguib, Y.M.A. Antioxidant activities of astaxanthin and related carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef]
- Shimidzu, N.; Goto, M.; Miki, W. Carotenoids as singlet oxygen quenchers in marine organisms. Fish. Sci. 1996, 62, 134–137. [Google Scholar] [CrossRef]
- Niu, J.; Tian, L.X.; Liu, Y.J.; Yang, H.J.; Ye, C.X.; Gao, W.; Mai, K.S. Effect of dietary astaxanthin on growth, survival, and stress tolerance of postlarval shrimp, Litopenaeus vannamei. J. World Aquac. Soc. 2009, 40, 795–802. [Google Scholar] [CrossRef]
- Lei, X.G.; Zhu, J.-H.; Cheng, W.-H.; Bao, Y.; Ho, Y.-S.; Reddi, A.R.; Holmgren, A.; Arnér, E.S.J. Paradoxical Roles of Antioxidant Enzymes: Basic Mechanisms and Health Implications. Physiol. Rev. 2016, 96, 307–364. [Google Scholar] [CrossRef]
- Moren, M.; Opstad, I.; Berntssen, M.; Infante, J.-L.Z.; Hamre, K. An optimum level of vitamin A supplements for Atlantic halibut (Hippoglossus hippoglossus L.) juveniles. Aquaculture 2004, 235, 587–599. [Google Scholar] [CrossRef]
- Hernandez, L.; Teshima, S.-I.; Ishikawa, M.; ALAM, S.; Koshio, S.; Tanaka, Y. Dietary vitamin A requirements of juvenile Japanese flounder Paralichthys olivaceus. Aquac. Nutr. 2005, 11, 3–9. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, X.; Bu, X.; Song, Y.; You, J.; Zhang, C.; Qin, C.; Qin, J.; Chen, L. Dietary vitamin A affects growth performance, immunity, antioxidant capacity, and lipid metabolism of juvenile Chinese mitten crab Eriocheir sinensis. Aquaculture 2022, 548, 737556. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.J.; Tian, L.X.; Yang, H.J.; Liang, G.Y.; Yue, Y.R.; Xu, D.H. Effects of dietary astaxanthin on growth, antiox idant capacity and gene expression in Pacific white shrimp Litopenaeus vannamei. Aquac. Nutr. 2013, 19, 917–927. [Google Scholar] [CrossRef]
- Behringer, D.C. Diseases of wild and cultured juvenile crustaceans: Insights from below the minimum landing size. J. Invertebr. Pathol. 2012, 110, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.X.; Xiong, F. Astaxanthin and its effects in inflammatory responses and inflammation-associated diseases: Recent advances and future directions. Molecules 2020, 25, 5342. [Google Scholar] [CrossRef] [PubMed]
- Muta, T.; Iwanaga, S. The role of hemolymph coagulation in innate immunity. Curr. Opin. Immunol. 1996, 8, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; Renault, T. Enzymatic activities in european flat oyster, ostrea edulis, and Pacific oyster, crassostrea gigas, hemlymph. J. Invertebr. Pathol. 2000, 76, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Lai, Y.C.; Chang, C.L.; Hung, W.C.; Wu, H.M.; Liao, Y.C.; Huang, C.H.; Liu, W.R. Facile and green synthesis of graphene-based conductive adhesives via liquid exfoliation process. Nanomaterials 2018, 9, 38. [Google Scholar] [CrossRef]
- Jiang, M.; Zhao, H.H.; Zai, S.W.; Shepherd, B.; Wen, H.; Deng, D.F. A defatted microalgae meal (Haematococcus pluvialis) as a partial protein source to replace fishmeal for feeding juvenile yellow perch Perca flavescens. J. Appl. Phycol. 2019, 31, 1197–1205. [Google Scholar] [CrossRef]
- Cheng, Y.; Wu, S. Effect of dietary astaxanthin on the growth performance and nonspecific immunity of red swamp crayfish Procambarus clarkii. Aquaculture 2019, 512, 734341. [Google Scholar] [CrossRef]
- Leone, A.M.; Palmer, R.M.; Knowles, R.G.; Francis, P.L.; Ashton, D.S.; Moncada, S. Constitutive and inducible nitric oxide synthases incorporate molecular oxygen into both nitric oxide and citrulline. J. Biol. Chem. 1991, 266, 23790–23795. [Google Scholar] [CrossRef]
- Chakravortty, D.; Hensel, M. Inducible nitric oxide synthase and control of intracellular bacterial pathogens. Microbes Infect. 2003, 5, 621–627. [Google Scholar] [CrossRef]
- Jiang, G.; Yu, R.; Zhou, M. Studies on nitric oxide synthase activity in haemocytes of shrimps Fenneropenaeus chinensis and Marsupenaeus japonicus after white spot syndrome virus infection. Nitric. Oxide. 2006, 14, 219–227. [Google Scholar] [CrossRef]
- Verma, R.; Balakrishnan, L.; Sharma, K.; Khan, A.A.; Advani, J.; Gowda, H.; Tripathy, S.P.; Suar, M.; Pandey, A.; Gandotra, S.; et al. A network map of Interleukin-10 signaling pathway. J. Cell Commun. Signal 2016, 10, 61–67. [Google Scholar] [CrossRef]
- He, Q.; Xiao, S.; Zhang, C.; Zhang, Y.; Shi, H.; Zhang, H.; Lin, F.; Liu, X.; Yang, H.; Wang, Q.; et al. Modulation of the growth performance, biochemical parameters, and non-specific immune responses of the hybrid grouper (Epinephelus fuscoguttatus♀ × E. lanceolatus♂) by two kinds of Chinese herb. Aquac. Rep. 2021, 19, 100604. [Google Scholar] [CrossRef]
- Yin, B.; Liu, H.; Tan, B.; Dong, X.; Chi, S.; Yang, Q.; Zhang, S.; Chen, L. Cottonseed protein concentrate (CPC) suppresses immune function in different intestinal segments of hybrid grouper ♀Epinephelus fuscoguttatus × ♂Epinephelus lanceolatu via TLR-2/MyD88 signaling pathways. Fish. Shellfish Immun. 2018, 81, 318–328. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, Y.H.; Wang, Z.; Wang, W.; Ren, Q. Novel myeloid differentiation factor 88, EsMyD88, exhibits EsTube-binding activity in Chinese mitten crab Eriocheir sinensis. Dev. Comp. Immunol. 2014, 47, 298–308. [Google Scholar] [CrossRef]
- Li, F.; Xiang, J. Recent advances in researches on the innate immunity of shrimp in China. Dev. Comp. Immunol. 2013, 39, 11–26. [Google Scholar] [CrossRef]
- Moynagh, P.N. TLR signalling and activation of IRFs: Revisiting old friends from the NF-κB pathway. Trends Immunol. 2005, 26, 469–476. [Google Scholar] [CrossRef]
- Kanwugu, O.N.; Glukhareva, T.V. Activation of Nrf2 pathway as a protective mechanism against oxidative stress-induced diseases: Potential of astaxanthin. Arch. Biochem. Biophys. 2023, 741, 109601. [Google Scholar] [CrossRef]
- Zhang, W.; Feng, C.; Jiang, H. Novel target for treating Alzheimer’s Diseases: Crosstalk between the Nrf2 pathway and autophagy. Ageing Res. Rev. 2021, 65, 101207. [Google Scholar] [CrossRef]
- Mu, C.; Zhao, J.; Wang, L.; Song, L.; Song, X.; Zhang, H.; Qiu, L.; Gai, Y.; Cui, Z. A thioredoxin with antioxidant activity identified from Eriocheir sinensis. Fish. Shellfish Immun. 2009, 26, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Mu, C.; Zhao, J.; Wang, L.; Song, L.; Zhang, H.; Li, C.; Qiu, L.; Gai, Y. Molecular cloning and characterization of peroxire doxin 6 from Chinese mitten crab Eriocheir sinensis. Fish. Shellfish Immun. 2009, 26, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Dai, A.; Liu, F.; Guan, Y. Effects of dietary astaxanthin on the immune response, resistance to white spot syn drome virus and transcription of antioxidant enzyme genes in Pacific white shrimp Litopenaeus vannamei. Iran. J. Fish. Sci. 2015, 14, 114476. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar] [CrossRef]
- Barrento, S.; Marques, A.; Teixeira, B.; Mendes, R.; Bandarra, N.; Vaz-Pires, P.; Nunes, M.L. Chemical composition, cholesterol, fatty acid and amino acid in two populations of brown crab Cancer pagurus: Ecological and human health implications. J. Food Compos. Anal. 2010, 23, 716–725. [Google Scholar] [CrossRef]
- Wang, Z.; Cai, C.; Cao, X.; Zhu, J.; He, J.; Wu, P.; Ye, Y. Supplementation of dietary astaxanthin alleviated oxidative damage induced by chronic high pH stress, and enhanced carapace astaxanthin concentration of Chinese mitten crab Eriocheir sinensis. Aquaculture 2018, 483, 230–237. [Google Scholar] [CrossRef]
- Wei, B.; Yang, Z.; Cheng, Y.; Wang, J.; Zhou, J. Effects of the complete replacement of fish oil with linseed oil on growth, fatty acid composition, and protein expression in the Chinese mitten crab (Eriocheir sinensis). Proteome Sci. 2018, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Sun, P.; Jin, M.; Wang, X.; Zhou, Q. Regulation of dietary lipid sources on tissue lipid classes and mitochondrial energy metabolism of juvenile swimming crab, Portunus trituberculatus. Front. Physiol. 2019, 10, 454. [Google Scholar] [CrossRef]
- Yuan, Y.; Xu, F.; Jin, M.; Wang, X.; Hu, X.; Zhao, M.; Cheng, X.; Luo, J.; Jiao, L.; Betancor, M.B.; et al. Untargeted lipidomics reveals metabolic responses to different dietary n-3 PUFA in juvenile swimming crab (Portunus trituberculatus). Food Chem. 2021, 354, 129570. [Google Scholar] [CrossRef]
- Tan, S.J.; Zhang, X.; Jin, X.K.; Li, W.W.; Li, J.Y.; Wang, Q. Fatty acid binding protein FABP3 from Chinese mitten crab Eriocheir sinensis participates in antimicrobial responses. Fish. Shellfish Immun. 2015, 43, 264–274. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (Dry Weight, %) | Dietary Astaxanthin Levels (mg/kg) | |||||
|---|---|---|---|---|---|---|
| 0.0 | 24.2 | 45.8 | 72.4 | 94.2 | 195.0 | |
| Peru fish meal | 30.00 | 30.00 | 30.00 | 30.00 | 30.00 | 30.00 |
| Soybean meal | 22.00 | 22.00 | 22.00 | 22.00 | 22.00 | 22.00 |
| Soybean protein concentrate | 6.00 | 6.00 | 6.00 | 6.00 | 6.00 | 6.00 |
| Poultry by-product meal | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 |
| Yeast extract | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 |
| Wheat flour | 22.00 | 22.00 | 22.00 | 22.00 | 22.00 | 22.00 |
| Fish oil | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 |
| Soybean lecithin | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 |
| Cholesterol | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 |
| Vitamin premix 1 | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 |
| Mineral premix 2 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Ca(H2PO4) 2 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Choline chloride | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 |
| Sodium alginate | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| BHT | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 |
| Cellulose | 0.50 | 0.475 | 0.45 | 0.425 | 0.40 | 0.30 |
| Astaxanthin 3 (10%) | 0.00 | 0.025 | 0.05 | 0.075 | 0.10 | 0.20 |
| Proximate composition (%) | ||||||
| Dry matter | 95.33 | 95.02 | 95.19 | 94.58 | 95.19 | 95.09 |
| Crude protein | 46.05 | 46.25 | 46.35 | 46.28 | 46.45 | 46.64 |
| Crude lipid | 10.05 | 10.42 | 10.37 | 10.32 | 10.72 | 10.30 |
| Ash | 9.73 | 9.75 | 9.85 | 9.62 | 9.80 | 9.91 |
| Astaxanthin (mg/kg) | 0.00 | 24.20 | 45.80 | 72.40 | 94.20 | 195.00 |
| Fatty Acids (mg/g) | Dietary Astaxanthin Levels (mg/kg) | |||||
|---|---|---|---|---|---|---|
| 0.0 | 24.2 | 45.8 | 72.4 | 94.2 | 195.0 | |
| 12:00 | 0.08 ± 0.01 | 0.06 ± 0.02 | 0.07 ± 0.01 | 0.10 ± 0.02 | 0.09 ± 0.01 | 0.05 ± 0.00 |
| 14:00 | 4.36 ± 0.54 a | 4.72 ± 0.46 b | 4.03 ± 0.52 ab | 5.06 ± 0.65 b | 5.33 ± 0.08 b | 2.79 ± 0.05 a |
| 16:00 | 41.07 ± 1.43 b | 38.70 ± 0.96 b | 37.08 ± 1.91 b | 40.60 ± 3.03 b | 37.28 ± 3.09 b | 28.34 ± 0.27 a |
| 18:00 | 17.28 ± 0.91 b | 15.04 ± 0.85 b | 15.24 ± 0.62 b | 17.31 ± 1.31 b | 15.15 ± 1.35 b | 11.37 ± 0.44 a |
| 20:00 | 0.92 ± 0.07 c | 0.81 ± 0.03 bc | 0.76 ± 0.08 bc | 0.79 ± 0.04 bc | 0.66 ± 0.09 ab | 0.47 ± 0.03 a |
| ∑SFA 1 | 63.71 ± 2.95 b | 59.35 ± 2.3 b | 57.18 ± 2.9 b | 63.86 ± 4.79 b | 57.84 ± 5.19 b | 43.02 ± 0.75 a |
| 16:1n-7 | 7.68 ± 0.81 b | 7.99 ± 0.80 b | 8.24 ± 0.50 b | 7.65 ± 0.08 b | 9.17 ± 0.40 b | 4.99 ± 0.33 a |
| 18:1n-9 | 75.41 ± 4.09 b | 66.24 ± 0.09 b | 66.89 ± 3.53 b | 75.87 ± 5.88 b | 66.77 ± 5.46 b | 48.28 ± 1.22 a |
| 20:1n-9 | 3.20 ± 0.31 b | 2.41 ± 0.09 ab | 2.70 ± 0.27 b | 2.81 ± 0.32 b | 2.50 ± 0.21 b | 1.67 ± 0.06 a |
| 22:1n-11 | 0.56 ± 0.03 c | 0.42 ± 0.02 bc | 0.44 ± 0.00 bc | 0.50 ± 0.07 bc | 0.38 ± 0.06 ab | 0.26 ± 0.02 a |
| ∑MUFA 2 | 86.86 ± 5.23 b | 77.07 ± 0.60 b | 77.28 ± 4.37 b | 88.29 ± 7.64 b | 77.62 ± 6.93 b | 55.20 ± 1.54 a |
| 18:2n-6 (LA) | 24.85 ± 1.77 b | 25.30 ± 0.85 b | 23.89 ± 0.66 b | 25.95 ± 1.97 b | 26.35 ± 1.10 b | 16.71 ± 0.36 a |
| 18:3n-6 | 0.09 ± 0.01 ab | 0.11 ± 0.00 b | 0.08 ± 0.01 ab | 0.11 ± 0.02 b | 0.12 ± 0.01 b | 0.06 ± 0.01 a |
| 20:2n-6 | 6.42 ± 0.21 d | 4.78 ± 0.02 b | 5.64 ± 0.33 c | 5.29 ± 0.36 bc | 5.46 ± 0.05 bc | 4.00 ± 0.05 a |
| 20:4n-6 (ARA) | 2.58 ± 0.09 cd | 2.78 ± 0.16 d | 2.37 ± 0.08 bc | 2.70 ± 0.18 cd | 2.08 ± 0.05 ab | 1.89 ± 0.01 a |
| ∑n-6 PUFA 3 | 34.49 ± 2.29 b | 32.87 ± 1.01 b | 31.15 ± 1.36 b | 35.18 ± 2.70 b | 31.70 ± 2.43 b | 22.67 ± 0.39 a |
| 18:3n-3 (ALA) | 3.99 ± 0.15 b | 4.09 ± 0.39 b | 3.23 ± 0.50 ab | 4.03 ± 0.56 b | 4.45 ± 0.21 b | 2.26 ± 0.10 a |
| 18:4n-3 | 0.45 ± 0.01 b | 0.50 ± 0.08 b | 0.41 ± 0.04 b | 0.45 ± 0.05 b | 0.53 ± 0.05 b | 0.24 ± 0.01 a |
| 20:4n3 | 0.37 ± 0.04 | 0.41 ± 0.06 | 0.42 ± 0.08 | 0.59 ± 0.09 | 0.51 ± 0.17 | 0.37 ± 0.04 |
| 20:5n-3 (EPA) | 17.93 ± 1.11 b | 16.75 ± 0.51 b | 16.17 ± 1.00 b | 18.27 ± 1.76 b | 18.48 ± 0.84 b | 11.68 ± 0.48 a |
| 22:5n-3 | 2.61 ± 0.24 b | 2.42 ± 0.11 b | 2.38 ± 0.14 b | 2.88 ± 0.26 b | 2.89 ± 0.08 b | 1.75 ± 0.04 a |
| 22:6n-3 (DHA) | 10.31 ± 0.61 ab | 11.67 ± 0.80 bc | 11.12 ± 0.40 bc | 12.52 ± 1.10 c | 12.57 ± 0.52 c | 8.67 ± 0.11 a |
| ∑n-3 PUFA 4 | 35.69 ± 2.62 b | 36.34 ± 2.24 b | 33.26 ± 2.32 ab | 38.10 ± 4.03 b | 40.24 ± 1.52 b | 24.97 ± 0.69 a |
| DHA/EPA | 1.66 ± 0.03 b | 1.51 ± 0.06 b | 1.52 ± 0.07 b | 1.55 ± 0.07 b | 1.39 ± 0.03 a | 1.35 ± 0.04 a |
| ∑TFA 5 | 220.75 ± 12.95 b | 205.63 ± 6.15 b | 198.87 ± 10.63 b | 225.43 ± 19.10 b | 203.39 ± 18.64 b | 145.86 ± 3.10 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Y.; Xie, S.; Zhan, W.; Peng, H.; Cao, H.; Tang, Z.; Tian, Y.; Zhu, T.; Jin, M.; Zhou, Q. Dietary Astaxanthin Can Promote the Growth and Motivate Lipid Metabolism by Improving Antioxidant Properties for Swimming Crab, Portunus trituberculatus. Antioxidants 2024, 13, 522. https://doi.org/10.3390/antiox13050522
Deng Y, Xie S, Zhan W, Peng H, Cao H, Tang Z, Tian Y, Zhu T, Jin M, Zhou Q. Dietary Astaxanthin Can Promote the Growth and Motivate Lipid Metabolism by Improving Antioxidant Properties for Swimming Crab, Portunus trituberculatus. Antioxidants. 2024; 13(5):522. https://doi.org/10.3390/antiox13050522
Chicago/Turabian StyleDeng, Yao, Shichao Xie, Wenhao Zhan, Hongyu Peng, Haiqing Cao, Zheng Tang, Yinqiu Tian, Tingting Zhu, Min Jin, and Qicun Zhou. 2024. "Dietary Astaxanthin Can Promote the Growth and Motivate Lipid Metabolism by Improving Antioxidant Properties for Swimming Crab, Portunus trituberculatus" Antioxidants 13, no. 5: 522. https://doi.org/10.3390/antiox13050522
APA StyleDeng, Y., Xie, S., Zhan, W., Peng, H., Cao, H., Tang, Z., Tian, Y., Zhu, T., Jin, M., & Zhou, Q. (2024). Dietary Astaxanthin Can Promote the Growth and Motivate Lipid Metabolism by Improving Antioxidant Properties for Swimming Crab, Portunus trituberculatus. Antioxidants, 13(5), 522. https://doi.org/10.3390/antiox13050522