The Extract of D. dasycarpus Ameliorates Oxazolone-Induced Skin Damage in Mice by Anti-Inflammatory and Antioxidant Mechanisms



Abstract
:1. Introduction
2. Materials and Methods
2.1. Herb Extraction and Chemicals
2.2. Animals and Treatments
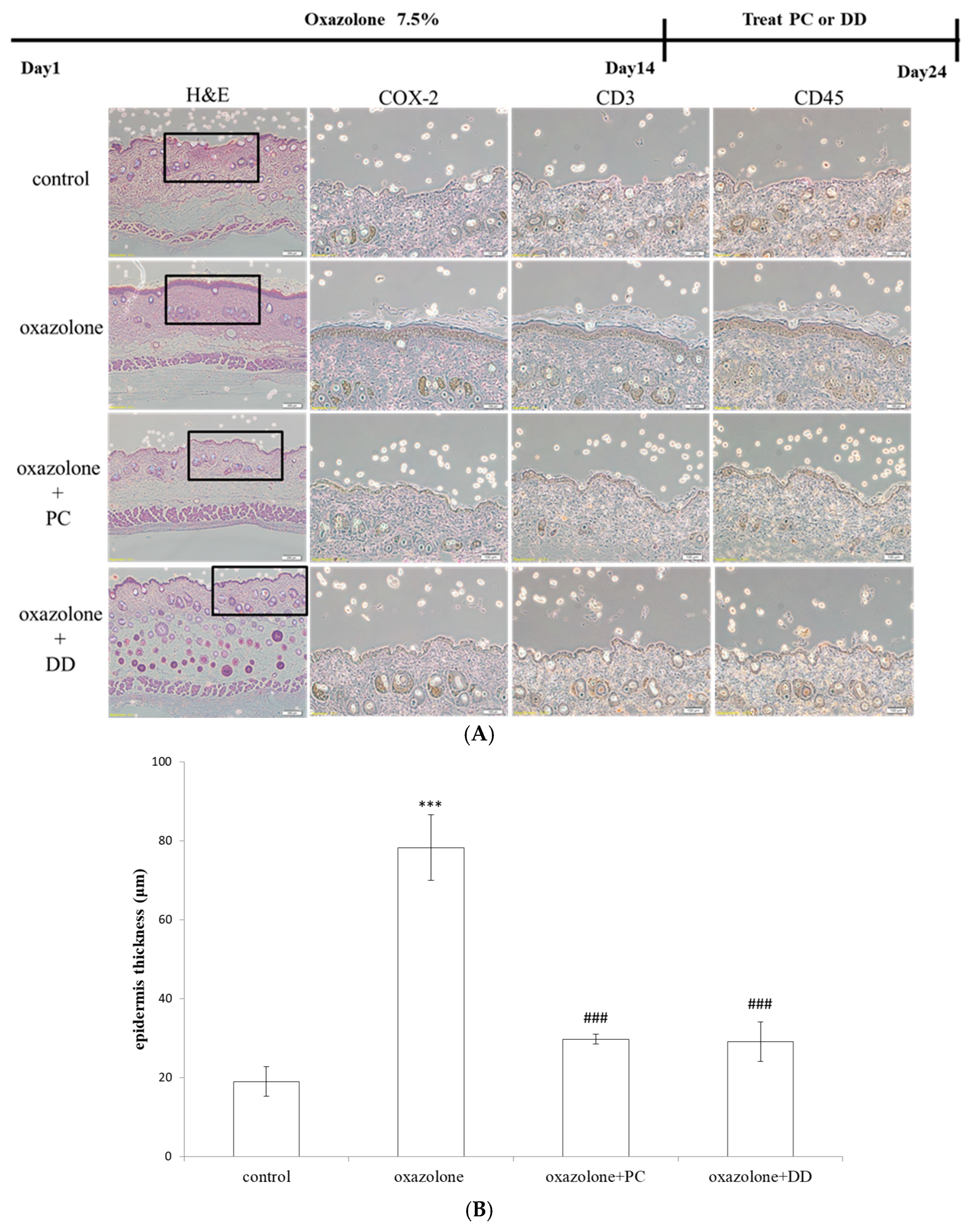
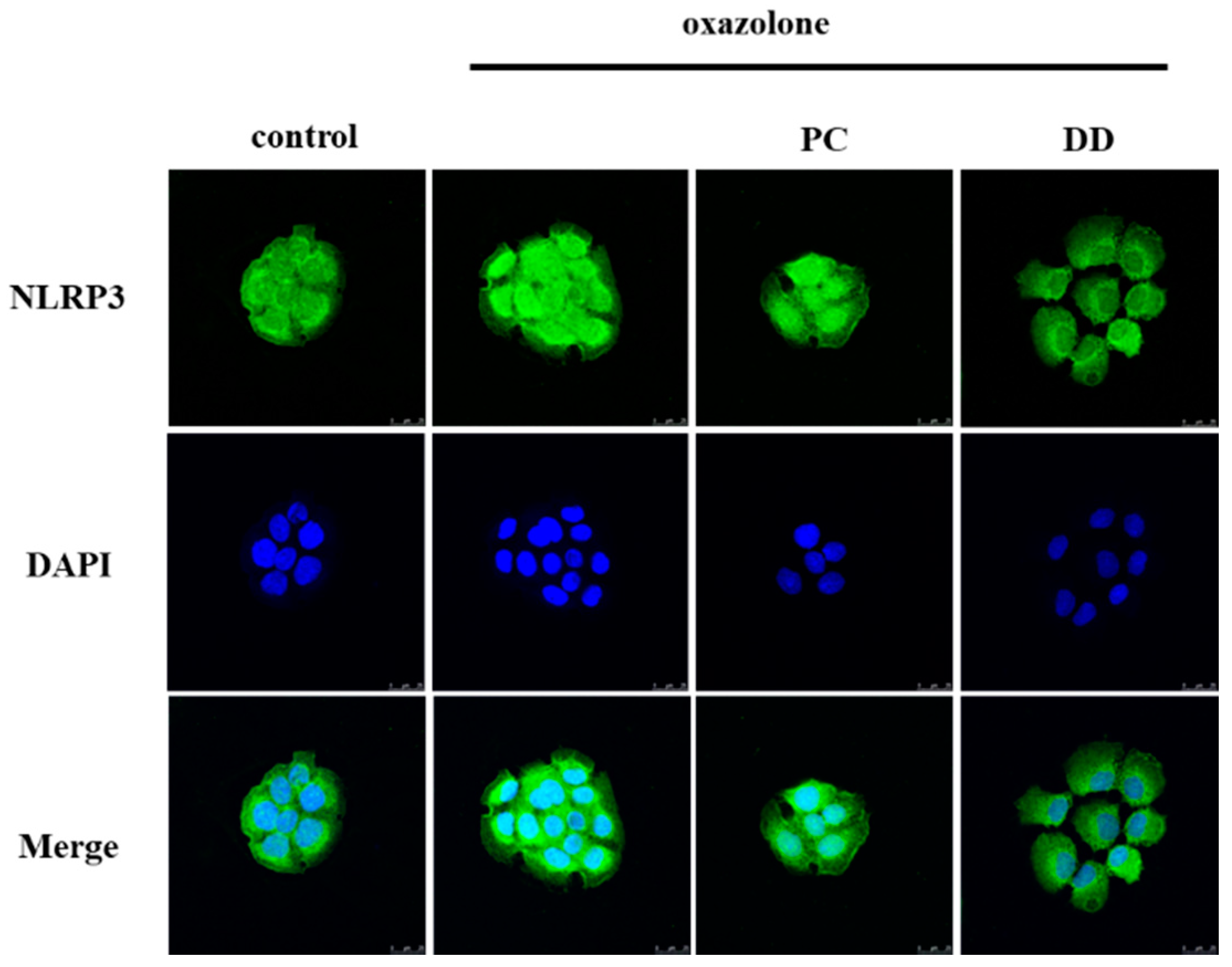
2.3. Specimens and Immunohistochemistry
2.4. Cell Culture
2.5. Cytokines Measurement
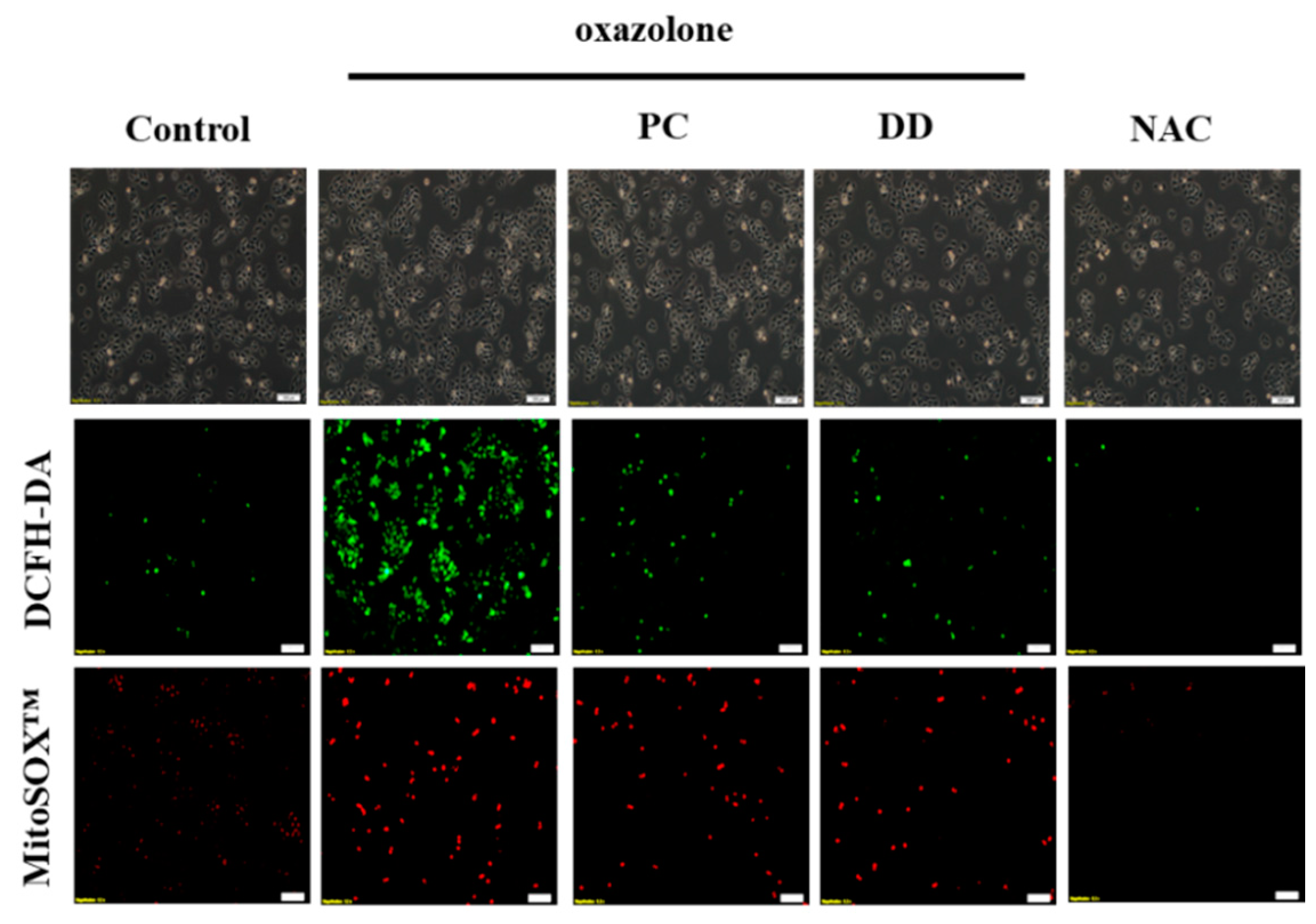
2.6. Reactive Oxygen Species (ROS) Measurement
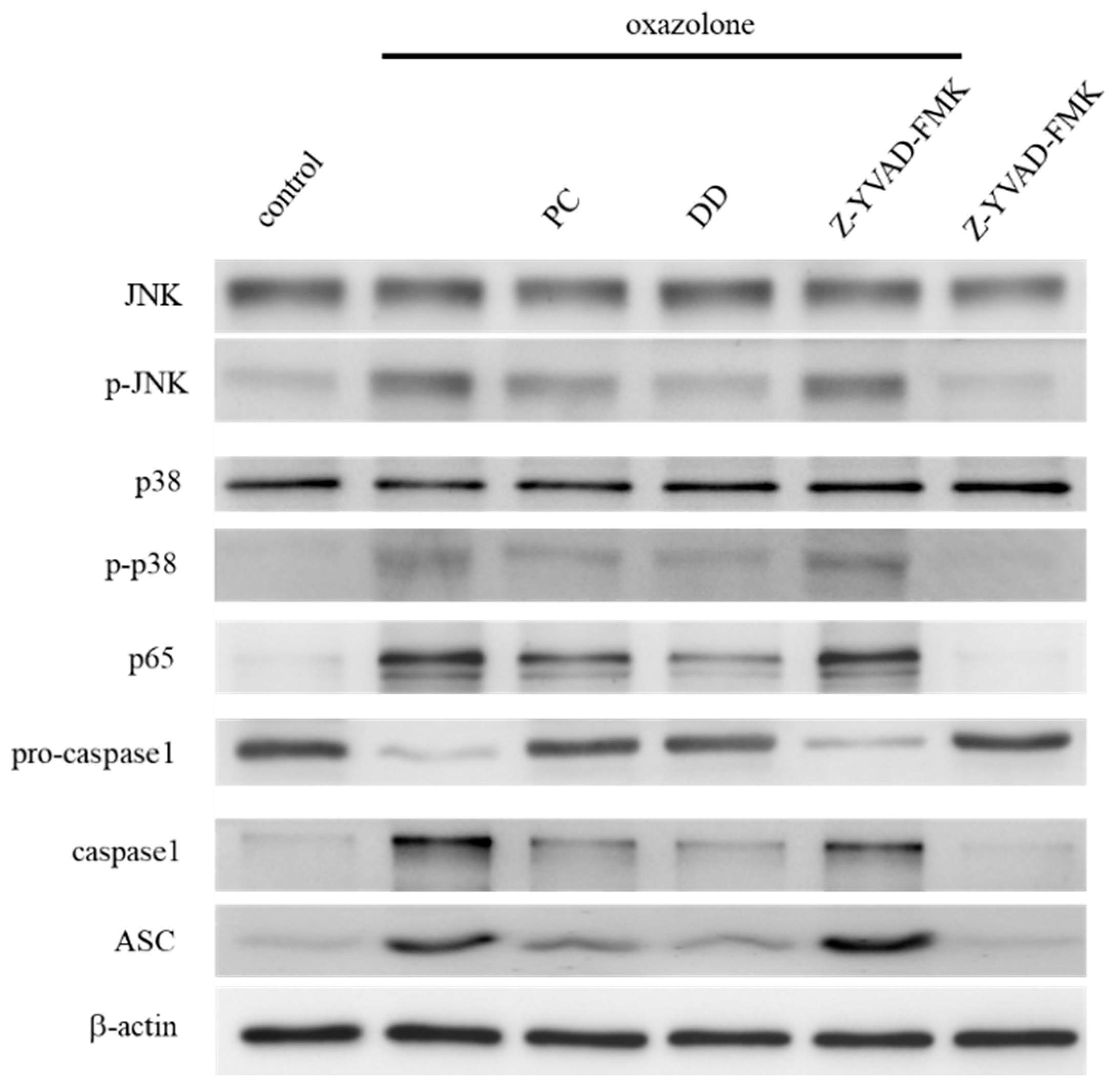
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, F.-P.; Chen, T.-J.; Kung, Y.-Y.; Chen, Y.-C.; Chou, L.-F.; Chen, F.-J.; Hwang, S.-J. Use frequency of traditional Chinese medicine in Taiwan. BMC Health Serv. Res. 2007, 7, 26. [Google Scholar] [CrossRef] [PubMed]
- Weng, S.-W.; Chen, B.-C.; Wang, Y.-C.; Liu, C.-K.; Sun, M.-F.; Chang, C.-M.; Lin, J.-G.; Yen, H.-R. Traditional Chinese medicine use among patients with psoriasis in Taiwan: A nationwide population-based study. Evid. Based Complement. Altern. Med. 2016, 2016, 13. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-F.; Liu, P.-H.; Huang, T.-P.; Lien, A.S.-Y.; Ou, L.-S.; Yu, C.-H.; Yang, S.-L.; Chang, H.-H.; Yen, H.-R. Characteristics and prescription patterns of traditional Chinese medicine in atopic dermatitis patients: Ten-year experiences at a Medical Center in Taiwan. Complement. Ther. Med. 2014, 22, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.-S.; Wang, M.-L.; Shyur, H.-J.; Leu, Y.-L.; Chan, Y.-Y.; Teng, C.-M.; Kuo, S.-C. Chemical constituents and bioactive principles from the root bark of Dictamnus dasycarpus+. Chin. Pharm. J. 1994, 46, 447–455. [Google Scholar]
- Yang, B.; Lee, H.-B.; Kim, S.; Park, Y.C.; Kim, K.; Kim, H. Decoction of Dictamnus Dasycarpus Turcz. Root Bark Ameliorates Skin Lesions and Inhibits Inflammatory Reactions in Mice with Contact Dermatitis. Pharmacogn. Mag. 2017, 13, 483–487. [Google Scholar] [PubMed]
- Gasparrini, M.; Forbes-Hernandez, T.Y.; Afrin, S.; Reboredo-Rodriguez, P.; Cianciosi, D.; Mezzetti, B.; Quiles, J.L.; Bompadre, S.; Battino, M.; Giampieri, F. Strawberry-based cosmetic formulations protect human dermal fibroblasts against UVA-induced damage. Nutrients 2017, 9, 605. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, K.C.M.; Chibli, L.A.; Santos, B.C.S.; Temponi, V.S.; Pinto, N.C.C.; Scio, E.; Del-Vechio-Vieira, G.; Alves, M.S.; Sousa, O.V. Evidence of bioactive compounds from Vernonia polyanthes leaves with topical anti-inflammatory potential. Int. J. Mol. Sci. 2016, 17, 1929. [Google Scholar] [CrossRef] [PubMed]
- Man, M.-Q.; Hatano, Y.; Lee, S.H.; Man, M.; Chang, S.; Feingold, K.R.; Leung, D.Y.M.; Holleran, W.; Uchida, Y.; Elias, P.M. Characterization of a hapten-induced, murine model with multiple features of atopic dermatitis: Structural, immunologic, and biochemical changes following single versus multiple oxazolone challenges. J. Investig. Dermatol. 2008, 128, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Saint-Mezard, P.; Krasteva, M.; Chavagnac, C.; Bosset, S.; Akiba, H.; Kehren, J.; Nicolas, J.F.; Berard, F.; Kanitakis, J.; Kaiserlian, D. Afferent and efferent phases of allergic contact dermatitis (ACD) can be induced after a single skin contact with haptens: Evidence using a mouse model of primary ACD. J. Investig. Dermatol. 2003, 120, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Pasparakis, M.; Haase, I.; Nestle, F.O. Mechanisms regulating skin immunity and inflammation. Nat. Rev. Immunol. 2014, 14, 289. [Google Scholar] [CrossRef] [PubMed]
- Andor, P.; Istvan, N.; Lajos, K. Innate immunity in the skin: How keratinocytes fight against pathogens. Curr. Immunol. Rev. 2005, 1, 29–42. [Google Scholar]
- McKenzie, R.C.; Sauder, D.N. Keratinocyte cytokines and growth factors. Functions in skin immunity and homeostasis. Dermatol. Clin. 1990, 8, 649–661. [Google Scholar] [PubMed]
- Ansel, J.; Perry, P.; Brown, J.; Damm, D.; Phan, T.; Hart, C.; Luger, T.; Hefeneider, S. Cytokine modulation of keratinocyte cytokines. J. Investig. Dermatol. 1990, 94, s101–s107. [Google Scholar] [CrossRef]
- Feldmeyer, L.; Keller, M.; Niklaus, G.; Hohl, D.; Werner, S.; Beer, H.D. The inflammasome mediates UVB-induced activation and secretion of interleukin-1β by keratinocytes. Curr. Biol. 2007, 17, 1140–1145. [Google Scholar] [CrossRef] [PubMed]
- Feldmeyer, L.; Werner, S.; French, L.E.; Beer, H.D. Interleukin-1, inflammasomes and the skin. Eur. J. Cell Biol. 2010, 89, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 2016, 16, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Zhong, F.L.; Mamai, O.; Sborgi, L.; Boussofara, L.; Hopkins, R.; Robinson, K.; Szeverenyi, I.; Takeichi, T.; Balaji, R.; Lau, A.; et al. Germline NLRP1 mutations cause skin inflammatory and cancer susceptibility syndromes via inflammasome activation. Cell 2016, 167, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Yamanashi, T.; Iwata, M.; Kamiya, N.; Tsunetomi, K.; Kajitani, N.; Wada, N.; Iitsuka, T.; Yamauchi, T.; Miura, A.; Pu, S.; et al. Beta-hydroxybutyrate, an endogenic NLRP3 inflammasome inhibitor, attenuates stress-induced behavioral and inflammatory responses. Sci. Rep. 2017, 7, 7677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, T.M.; Tsen, J.H.; Yen, H.; Yang, T.Y.; Huang, H.C. Extract from Periostracum cicadae inhibits oxidative stress and inflammation induced by ultraviolet B irradiation on HaCaT keratinocytes. Evid. Based Complement. Altern. Med. 2017, 2017, 8325049. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.; Ruben, Z.; Butchko, G. Histology of and quantitative assays for oxazolone-induced allergic contact dermatitis in mice. Am. J. Dermatopathol. 1986, 8, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.K. In vivo pharmacological disease models for psoriasis and atopic dermatitis in drug discovery. Basic Clin. Pharmacol. Toxicol. 2006, 99, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, S.P.; Chang, H.T.; Wang, Y.T.; Tu, P.F. Pressurized liquid extraction followed by high-performance liquid chromatography for determination of seven active compounds in Cortex Dictamni. J. Chromatogr. A 2006, 1108, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Zhou, L.; Zhang, J.; Yu, B.Y.; Li, C.Y.; Liu, Z.X.; Ju, W.Z. Determination of limonin in dog plasma by liquid chromatography–tandem mass spectrometry and its application to a pharmacokinetic study. Biomed. Chromatogr. 2013, 27, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Patlolla, J.M.; Rao, C.V. Triterpenoids for cancer prevention and treatment: Current status and future prospects. Curr. Pharm. Biotechnol. 2012, 13, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Radice, M.; Manfredini, S.; Ziosi, P.; Dissette, V.; Buso, P.; Fallacara, A.; Vertuani, S. Herbal extracts, lichens and biomolecules as natural photo-protection alternatives to synthetic UV filters. A systematic review. Fitoterapia 2016, 114, 144–162. [Google Scholar] [CrossRef] [PubMed]
- Itani, S.; Watanabe, T.; Nadatani, Y.; Sugimura, N.; Shimada, S.; Takeda, S.; Otani, K.; Hosomi, S.; Nagami, Y.; Tanaka, F. NLRP3 inflammasome has a protective effect against oxazolone-induced colitis: A possible role in ulcerative colitis. Sci. Rep. 2016, 6, 39075. [Google Scholar] [CrossRef] [PubMed]
- Fabio, M. Signaling by ROS drives inflammasome activation. Eur. J. Immunol. 2010, 40, 616–619. [Google Scholar] [Green Version]
- He, Y.; Hara, H.; Núñez, G. Mechanism and regulation of NLRP3 inflammasome activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Abais, J.M.; Xia, M.; Zhang, Y.; Boini, K.M.; Li, P.-L. Redox regulation of NLRP3 inflammasomes: ROS as trigger or effector? Antioxid. Redox Signal. 2015, 22, 1111–1129. [Google Scholar] [CrossRef] [PubMed]
- Tschopp, J.; Schroder, K. NLRP3 inflammasome activation: The convergence of multiple signalling pathways on ROS production? Nat. Rev. Immunol. 2010, 10, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Mukaida, N.; Okamoto, S.; Ishikawa, Y.; Matsushima, K. Molecular mechanism of interleukin-8 gene expression. J. Leukoc. Biol. 1994, 56, 554–558. [Google Scholar] [CrossRef] [PubMed]
- Tak, P.P.; Firestein, G.S. NF-κB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Song, X.; Bi, Z.; Chu, W.; Wan, Y. UV-induced NF-κB activation and expression of IL-6 is attenuated by (-)-epigallocatechin-3-gallate in cultured human keratinocytes in vitro. Int. J. Mol. Med. 2005, 16, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-H.; Yoo, J.-M.; Lee, E.; Lee, B.; Cho, W.-K.; Park, K.-I.; Yeul Ma, J. Anti-inflammatory effects of Perillae Herba ethanolic extract against TNF-α/IFN-γ-stimulated human keratinocyte HaCaT cells. J. Ethnopharmacol. 2018, 211, 217–223. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oxazolone | ||||
|---|---|---|---|---|
| Control | PC | DD | ||
| TNF-α pg/mL | 20.64 ± 0.94 | 96.51 ± 9.74 | 87.97 ± 2.35 | 71.85 ± 4.95 a |
| IL-6 pg/mL | 13.20 ± 2.87 | 27.40 ± 1.19 | 14.20 ± 2.43 b | 12.30 ± 2.17 b |
| IL-8 pg/mL | 20.64 ± 0.57 | 198.51 ± 14.24 | 87.97 ± 3.75 b | 51.85 ± 5.21 b |
| Oxazolone | |||||||
|---|---|---|---|---|---|---|---|
| Control | PC | DD | Z-YVAD-FMK | Z-YVAD-FMK | |||
| IL-1β pg/mL | 25.24 ± 0.57 | 180.11 ± 14.24 | 87.67 ± 4.95 a | 72.65 ± 7.21 a | 57.34 ± 4.54 b | 28.84 ± 0.27 b | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, T.-M.; Yang, T.-Y.; Niu, Y.-L.; Huang, H.-C. The Extract of D. dasycarpus Ameliorates Oxazolone-Induced Skin Damage in Mice by Anti-Inflammatory and Antioxidant Mechanisms. Antioxidants 2018, 7, 77. https://doi.org/10.3390/antiox7060077
Chang T-M, Yang T-Y, Niu Y-L, Huang H-C. The Extract of D. dasycarpus Ameliorates Oxazolone-Induced Skin Damage in Mice by Anti-Inflammatory and Antioxidant Mechanisms. Antioxidants. 2018; 7(6):77. https://doi.org/10.3390/antiox7060077
Chicago/Turabian StyleChang, Tsong-Min, Ting-Ya Yang, Yu-Lin Niu, and Huey-Chun Huang. 2018. "The Extract of D. dasycarpus Ameliorates Oxazolone-Induced Skin Damage in Mice by Anti-Inflammatory and Antioxidant Mechanisms" Antioxidants 7, no. 6: 77. https://doi.org/10.3390/antiox7060077
APA StyleChang, T.-M., Yang, T.-Y., Niu, Y.-L., & Huang, H.-C. (2018). The Extract of D. dasycarpus Ameliorates Oxazolone-Induced Skin Damage in Mice by Anti-Inflammatory and Antioxidant Mechanisms. Antioxidants, 7(6), 77. https://doi.org/10.3390/antiox7060077