4-Hydroxynonenal in Redox Homeostasis of Gastrointestinal Mucosa: Implications for the Stomach in Health and Diseases



Abstract
:1. Introduction
2. Approaches to Determine HNE in Samples of Patients Suffering from Stomach Diseases
3. HNE in the Stomach under Physiological Conditions
4. HNE in Patients with H. pylori-Associated Gastritis and Peptic Ulcer
5. HNE in Gastric Carcinogenesis
6. HNE in Alcohol- and Non-Steroid Anti-Inflammatory Drug (NSAID)-Induced Gastropathy
7. Pharmacological and Non-Pharmacological Approaches to Reduce Redox Imbalance in GM
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sherwood, L. Human Physiology: From Cells to Systems; Cengage Learning: Boston, MA, USA, 2015. [Google Scholar]
- Kanner, J.; Lapidot, T. The stomach as a bioreactor: Dietary lipid peroxidation in the gastric fluid and the effects of plant-derived antioxidants. Free Radic. Biol. Med. 2001, 31, 1388–1395. [Google Scholar] [CrossRef]
- Mentis, A.; Lehours, P.; Mégraud, F. Epidemiology and Diagnosis of Helicobacter pylori infection. Helicobacter 2015, 20 (Suppl. 1), 1–7. [Google Scholar] [CrossRef]
- Sgouras, D.N.; Trang, T.T.H.; Yamaoka, Y. Pathogenesis of Helicobacter pylori Infection. Helicobacter 2015, 20 (Suppl. 1), 8–16. [Google Scholar] [CrossRef] [PubMed]
- Pérez, S.; Taléns-Visconti, R.; Rius-Pérez, S.; Finamor, I.; Sastre, J. Redox signaling in the gastrointestinal tract. Free Radic. Biol. Med. 2017, 104, 75–103. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Zarkovic, N. 4-Hydroxynonenal as a bioactive marker of pathophysiological processes. Mol. Asp. Med. 2003, 24, 281–291. [Google Scholar] [CrossRef]
- Zarkovic, N.; Cipak Gasparovic, A.; Cindrić, M.; Waeg, G.; Borović Šunjić, S.; Mrakovčić, L.; Jaganjac, M.; Kolenc, D.; Andrišić, L.; Gverić Ahmetašević, S.; et al. 4-Hydroxynonenal-protein adducts as biomarkers of oxidative stress, lipid peroxidation and oxidative homeostasis. In Proceedings of the Free Radicals Health and Lifestyle, Rome, Italy, 26–29 August 2009; pp. 37–44. [Google Scholar]
- Žarković, N.; Žarković, K.; Schaur, R.J.; Štolc, S.; Schlag, G.; Redl, H.; Waeg, G.; Borović, S.; Lončarić, I.; Jurić, G.; et al. 4-Hydroxynonenal as a second messenger of free radicals and growth modifying factor. Life Sci. 1999, 65, 1901–1904. [Google Scholar] [CrossRef]
- Schaur, R.; Siems, W.; Bresgen, N.; Eckl, P. 4-Hydroxy-nonenal—A Bioactive Lipid Peroxidation Product. Biomolecules 2015, 5, 2247–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirpich, I.A.; Marsano, L.S.; McClain, C.J. Gut-Liver Axis, Nutrition, and Non Alcoholic Fatty Liver Disease. Clin. Biochem. 2015, 48, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Zarkovic, K.; Jakovcevic, A.; Zarkovic, N. Contribution of the HNE-immunohistochemistry to modern pathological concepts of major human diseases. Free Radic. Biol. Med. 2017, 111, 110–126. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S.; Miyake, N.; Hiai, H.; Hagiwara, M.; Kawakishi, S.; Osawa, T.; Uchida, K. The monoclonal antibody specific for the 4-hydroxy-2-nonenal histidine adduct. FEBS Lett. 1995, 359, 189–191. [Google Scholar] [CrossRef] [Green Version]
- Waeg, G.; Dimsity, G.; Esterbauer, H. Monoclonal antibodies for detection of 4-hydroxynonenal modified proteins. Free Radic. Res. 1996, 25, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.; Milkovic, L.; Bennett, S.J.; Griffiths, H.R.; Zarkovic, N.; Grune, T. Measurement of HNE-protein adducts in human plasma and serum by ELISA-Comparison of two primary antibodies. Redox Biol. 2013, 1, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Sousa, B.C.; Pitt, A.R.; Spickett, C.M. Chemistry and analysis of HNE and other prominent carbonyl-containing lipid oxidation compounds. Free Radic. Biol. Med. 2017, 111, 294–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarkovic, K.; Uchida, K.; Kolenc, D.; Hlupic, L.; Zarkovic, N. Tissue distribution of lipid peroxidation product acrolein in human colon carcinogenesis. Free Radic. Res. 2006, 40, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Cherkas, A.; Yelisyeyeva, O.; Semen, K.; Zarković, K.; Kaminskyy, D.; Gasparović, A.C.; Jaganjac, M.; Lutsyk, A.; Waeg, G.; Zarkovic, N. Persistent accumulation of 4-hydroxynonenal-protein adducts in gastric mucosa after Helicobacter pylori eradication. Coll. Antropol. 2009, 33, 815–821. [Google Scholar] [PubMed]
- Yelisyeyeva, O.; Cherkas, A.; Zarkovic, K.; Semen, K.; Kaminskyy, D.; Waeg, G.; Zarkovic, N. The distribution of 4-hydroxynonenal-modified proteins in gastric mucosa of duodenal peptic ulcer patients. Free Radic. Res. 2008, 42, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Cherkas, A.; Zarkovic, K.; Cipak Gasparovic, A.; Jaganjac, M.; Milkovic, L.; Abrahamovych, O.; Yatskevych, O.; Waeg, G.; Yelisyeyeva, O.; Zarkovic, N. Amaranth oil reduces accumulation of 4-hydroxynonenal-histidine adducts in gastric mucosa and improves heart rate variability in duodenal peptic ulcer patients undergoing Helicobacter pylori eradication. Free Radic. Res. 2018, 52, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Spickett, C.M.; Wiswedel, I.; Siems, W.; Zarkovic, K.; Zarkovic, N. Advances in methods for the determination of biologically relevant lipid peroxidation products. Free Radic. Res. 2010, 44, 1172–1202. [Google Scholar] [CrossRef] [PubMed]
- Spickett, C.M. The lipid peroxidation product 4-hydroxy-2-nonenal: Advances in chemistry and analysis. Redox Biol. 2013, 1, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Aslebagh, R.; Pfeffer, B.A.; Fliesler, S.J.; Darie, C.C. Mass spectrometry-based proteomics of oxidative stress: Identification of 4-hydroxy-2-nonenal (HNE) adducts of amino acids using lysozyme and bovine serum albumin as model proteins. Electrophoresis 2016, 37, 2615–2623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldini, G.; Gamberoni, L.; Orioli, M.; Beretta, G.; Regazzoni, L.; Facino, R.M.; Carini, M. Mass spectrometric characterization of covalent modification of human serum albumin by 4-hydroxy-trans-2-nonenal. J. Mass Spectrom. 2006, 41, 1149–1161. [Google Scholar] [CrossRef] [PubMed]
- Tirosh, O.; Shpaizer, A.; Kanner, J. Lipid Peroxidation in a Stomach Medium Is Affected by Dietary Oils (Olive/Fish) and Antioxidants: The Mediterranean versus Western Diet. J. Agric. Food Chem. 2015, 63, 7016–7023. [Google Scholar] [CrossRef] [PubMed]
- Negre-Salvayre, A.; Auge, N.; Ayala, V.; Basaga, H.; Boada, J.; Brenke, R.; Chapple, S.; Cohen, G.; Feher, J.; Grune, T.; et al. Pathological aspects of lipid peroxidation. Free Radic. Res. 2010, 44, 1125–1171. [Google Scholar] [CrossRef] [PubMed]
- Abadi, A.T.B. Strategies used by helicobacter pylori to establish persistent infection. World J. Gastroenterol. 2017, 23, 2870–2882. [Google Scholar] [CrossRef] [PubMed]
- Backert, S.; Schmidt, T.P.; Harrer, A.; Wessler, S. Exploiting the gastric epithelial barrier: Helicobacter pylori’s attack on tight and adherens junctions. In Current Topics in Microbiology and Immunology; Springer: Berlin, Germany, 2017; ISBN 9783319505206. [Google Scholar]
- Barrera, G.; Pizzimenti, S.; Ciamporcero, E.S.; Daga, M.; Ullio, C.; Arcaro, A.; Cetrangolo, G.P.; Ferretti, C.; Dianzani, C.; Lepore, A.; et al. Role of 4-Hydroxynonenal-Protein Adducts in Human Diseases. Antioxid. Redox Signal. 2015, 22, 1681–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mol, M.; Regazzoni, L.; Altomare, A.; Degani, G.; Carini, M.; Vistoli, G.; Aldini, G. Enzymatic and non-enzymatic detoxification of 4-hydroxynonenal: Methodological aspects and biological consequences. Free Radic. Biol. Med. 2017, 111, 328–344. [Google Scholar] [CrossRef] [PubMed]
- Cid, T.P.; Fernández, M.C.; Benito Martínez, S.; Jones, N.L. Pathogenesis of Helicobacter pylori infection. Helicobacter 2013, 18 (Suppl. 1), 12–17. [Google Scholar] [CrossRef]
- Cherkas, A.; Abrahamovych, O.; Golota, S.; Nersesyan, A.; Pichler, C.; Serhiyenko, V.; Knasmüller, S.; Zarkovic, N.; Eckl, P. The correlations of glycated hemoglobin and carbohydrate metabolism parameters with heart rate variability in apparently healthy sedentary young male subjects. Redox Biol. 2015, 5, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Cherkas, A.; Zhuraev, R. A marked decrease in heart rate variability in marfan syndrome patients with confirmed FBN1 mutations. Cardiol. J. 2016, 23, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Levenstein, S.; Rosenstock, S.; Jacobsen, R.K.; Jorgensen, T. Psychological stress increases risk for peptic ulcer, regardless of Helicobacter pylori infection or use of nonsteroidal anti-inflammatory drugs. Clin. Gastroenterol. Hepatol. 2015, 13, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Haensel, A.; Mills, P.J.; Nelesen, R.A.; Ziegler, M.G.; Dimsdale, J.E. The relationship between heart rate variability and inflammatory markers in cardiovascular diseases. Psychoneuroendocrinology 2008, 33, 1305–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, M.; Yukinaka, M.; Miyajima, H.; Nada, T.; Kondo, Y.; Okahisa, T.; Shibata, H.; Okamura, S.; Honda, H.; Shimizu, I.; et al. Is autonomic dysfunction a necessary condition for chronic peptic ulcer formation? Aliment. Pharmacol. Ther. 2000, 14 (Suppl. 1), 82–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehrens, S.M.T.; Hampton, S.M.; Skene, D.J. Heart rate variability and endothelial function after sleep deprivation and recovery sleep among male shift and non-shift workers. Scand. J. Work Environ. Health 2012, 38, 171–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yelisyeyeva, O.; Cherkas, A.; Semen, K.; Kaminskyy, D.; Lutsyk, A. Study of aerobic metabolism parameters and heart rate variability and their correlations in elite athletes: A modulatory effect of amaranth oil. Clin. Exp. Med. J. 2009, 3, 293–307. [Google Scholar] [CrossRef]
- Matteoli, G.; Boeckxstaens, G.E. The vagal innervation of the gut and immune homeostasis. Gut 2013, 62, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Hoover, D.B. Cholinergic modulation of the immune system presents new approaches for treating inflammation. Pharmacol. Ther. 2017, 179, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Steinhorn, B.; Sartoretto, J.L.; Sorrentino, A.; Romero, N.; Kalwa, H.; Abel, E.D.; Michel, T. Insulin-dependent metabolic and inotropic responses in the heart are modulated by hydrogen peroxide from NADPH-oxidase isoforms NOX2 and NOX4. Free Radic. Biol. Med. 2017, 113, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Contreras, C.; Nogueiras, R.; Diéguez, C.; Rahmouni, K.; López, M. Traveling from the hypothalamus to the adipose tissue: The thermogenic pathway. Redox Biol. 2017, 12, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Cherkas, A.; Yatskevych, O. The amplitude of heart rate oscillations is dependent on metabolic status of sinoatrial node cells. OA Med. Hypothesis 2014, 2, 1–8. [Google Scholar]
- Gao, L.; Zimmerman, M.C.; Biswal, S.; Zucker, I.H. Selective NRF2 Gene Deletion in the Rostral Ventrolateral Medulla Evokes Hypertension and Sympathoexcitation in Mice. Hypertension 2017, 69, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, S.A.; Kountouras, J. Novel Advances in the Association Between Helicobacter pylori Infection, Metabolic Syndrome, and Related Morbidity. Helicobacter 2015, 20, 405–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.-W.; Chien, C.-Y.; Yang, K.-J.; Kuo, S.-F.; Chen, C.-H.; Chien, R.-N. Helicobacter pylori Infection Increases Insulin Resistance and Metabolic Syndrome in Residents Younger than 50 Years Old: A Community-Based Study. PLoS ONE 2015, 10, e0128671. [Google Scholar] [CrossRef] [PubMed]
- Cherkas, A.; Eckl, P.; Gueraud, F.; Abrahamovych, O.; Serhiyenko, V.; Yatskevych, O.; Pliatsko, M.; Golota, S. Helicobacter pylori in sedentary men is linked to higher heart rate, sympathetic activity, and insulin resistance but not inflammation or oxidative stress. Croat. Med. J. 2016, 57, 141–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardaropoli, S.; Rolfo, A.; Todros, T. Helicobacter pylori and pregnancy-related disorders. World J. Gastroenterol. 2014, 20, 654–664. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, F.; Gasbarrini, A.; Polyzos, S.A.; Kountouras, J. Extragastric Diseases and Helicobacter pylori. Helicobacter 2015, 20 (Suppl. 1), 40–46. [Google Scholar] [CrossRef]
- Hardbower, D.M.; de Sablet, T.; Chaturvedi, R.; Wilson, K.T. Chronic inflammation and oxidative stress: The smoking gun for Helicobacter pylori-induced gastric cancer? Gut Microbes 2013, 4, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Handa, O.; Naito, Y.; Yoshikawa, T. Helicobacter pylori: A ROS-inducing bacterial species in the stomach. Inflamm. Res. 2010, 59, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Cherkas, A.; Golota, S.; Guéraud, F.; Abrahamovych, O.; Pichler, C.; Nersesyan, A.; Krupak, V.; Bugiichyk, V.; Yatskevych, O.; Pliatsko, M.; et al. A Helicobacter pylori-associated insulin resistance in asymptomatic sedentary young men does not correlate with inflammatory markers and urine levels of 8-iso-PGF2α or 1,4-dihydroxynonane mercapturic acid. Arch. Physiol. Biochem. 2018, 124, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Yeniova, A.O.; Uzman, M.; Kefeli, A.; Basyigit, S.; Ata, N.; Dal, K.; Guresci, S.; Nazligul, Y. Serum 8 Hydroxydeoxyguanosine and Cytotoxin Associated Gene A as Markers for Helicobacter pylori Infection. Asian Pac. J. Cancer Prev. 2015, 16, 5199–5203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative Stress: An Essential Factor in the Pathogenesis of Gastrointestinal Mucosal Diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwiecien, S.; Jasnos, K.; Magierowski, M.; Sliwowski, Z.; Pajdo, R.; Brzozowski, B.; Mach, T.; Wojcik, D.; Brzozowski, T. Lipid peroxidation, reactive oxygen species and antioxidative factors in the pathogenesis of gastric mucosal lesions and mechanism of protection against oxidative stress—Induced gastric injury. J. Physiol. Pharmacol. 2014, 65, 613–622. [Google Scholar] [PubMed]
- Awasthi, Y.C.; Ramana, K.V.; Chaudhary, P.; Srivastava, S.K.; Awasthi, S. Regulatory roles of glutathione-S-transferases and 4-hydroxynonenal in stress-mediated signaling and toxicity. Free Radic. Biol. Med. 2017, 111, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Cacev, T.; Cipak, A.; Kapitanović, S.; Gall Troselj, K.; Zarković, N. Even stressed cells are individuals: Second messengers of free radicals in pathophysiology of cancer. Croat. Med. J. 2012, 53, 304–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, G.; Zarkovic, N. Revealing mechanisms of selective, concentration-dependent potentials of 4-hydroxy-2-nonenal to induce apoptosis in cancer cells through inactivation of membrane-associated catalase. Free Radic. Biol. Med. 2015, 81, 128–144. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Xiao, M.; Zarkovic, K.; Zhu, M.; Sa, R.; Lu, J.; Tao, Y.; Chen, Q.; Xia, L.; Cheng, S.; et al. Mitochondrial control of apoptosis through modulation of cardiolipin oxidation in hepatocellular carcinoma: A novel link between oxidative stress and cancer. Free Radic. Biol. Med. 2017, 102, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Živković, N.P.; Petrovečki, M.; Lončarić, Č.T.; Nikolić, I.; Waeg, G.; Jaganjac, M.; Žarković, K.; Žarković, N. Positron emission tomography-computed tomography and 4-hydroxynonenal-histidine immunohistochemistry reveal differential onset of lipid peroxidation in primary lung cancer and in pulmonary metastasis of remote malignancies. Redox Biol. 2017, 11, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Zagari, R.M.; Rabitti, S.; Eusebi, L.H.; Bazzoli, F. Treatment of Helicobacter pylori infection: A clinical practice update. Eur. J. Clin. Investig. 2018, 48. [Google Scholar] [CrossRef] [PubMed]
- Buzás, G.M.; Gastroenterologist, C. Metabolic consequences of Helicobacter pylori infection and eradication. World J. Gastroenterol. 2014, 20, 5226–5234. [Google Scholar] [CrossRef] [PubMed]
- Dogan, Z.; Sarikaya, M.; Ergul, B.; Filik, L. The effect of Helicobacter pylori eradication on insulin resistance and HbA1c level in people with normal glucose levels: A prospective study. Biomed. Pap. Med. Fac. Univ. Palacký Olomouc Czechoslov. 2015, 159, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Longo-Mbenza, B.; Nkondi Nsenga, J.; Vangu Ngoma, D. Prevention of the metabolic syndrome insulin resistance and the atherosclerotic diseases in Africans infected by Helicobacter pylori infection and treated by antibiotics. Int. J. Cardiol. 2007, 121, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Anjan, D.; Dattagupta, S.; Ahuja, V.; Mathur, M.; Sharma, M.P. Pre and post eradication gastric inflammation in Helicobacter pylori-associated duodenal ulcer. Indian J. Gastroenterol. 2002, 21, 7–10. [Google Scholar] [PubMed]
- Lanas, A.; Chan, F.K.L. Peptic ulcer disease. Lancet 2017, 390, 613–624. [Google Scholar] [CrossRef]
- Hardbower, D.M.; Peek, R.M.; Wilson, K.T. At the Bench: Helicobacter pylori, dysregulated host responses, DNA damage, and gastric cancer. J. Leukoc. Biol. 2014, 96, 201–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, S.E.; Choi, K.D.; Choe, J.; Kim, S.O.; Na, H.K.; Choi, J.Y.; Ahn, J.Y.; Jung, K.W.; Lee, J.; Kim, D.H.; et al. The effect of eradication of Helicobacter pylori on gastric cancer prevention in healthy asymptomatic populations. Helicobacter 2018, 23, e12464. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.J.; Kook, M.-C.; Kim, Y.-I.; Cho, S.-J.; Lee, J.Y.; Kim, C.G.; Park, B.; Nam, B.-H. Helicobacter pylori Therapy for the Prevention of Metachronous Gastric Cancer. N. Engl. J. Med. 2018, 88, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Eckl, P.M. Genotoxicity of HNE. Mol. Asp. Med. 2003, 24, 161–165. [Google Scholar] [CrossRef]
- Gentile, F.; Arcaro, A.; Pizzimenti, S.; Daga, M.; Paolo Cetrangolo, G.; Dianzani, C.; Lepore, A.; Graf, M.; Ames, P.R.J.; Barrera, G. DNA damage by lipid peroxidation products: Implications in cancer, inflammation and autoimmunity. AIMS Genet. 2017, 4, 103–137. [Google Scholar] [CrossRef]
- Gasparovic, A.C.; Milkovic, L.; Sunjic, S.B.; Zarkovic, N. Cancer growth regulation by 4-hydroxynonenal. Free Radic. Biol. Med. 2017, 111, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, L.; Rong, S.; Qu, H.; Zhang, Y.; Chang, D.; Pan, H.; Wang, W. Relation between gastric cancer and protein oxidation, DNA damage, and lipid peroxidation. Oxid. Med. Cell. Longev. 2013, 2013, 543760. [Google Scholar] [CrossRef] [PubMed]
- Kwiecień, S.; Brzozowski, T.; Konturek, S.J. Effects of reactive oxygen species action on gastric mucosa in various models of mucosal injury. J. Physiol. Pharmacol. 2002, 53, 39–50. [Google Scholar] [PubMed]
- Wallace, J.; Pshyk-Titko, I.; Muscara, M.N.; Bula, N.; Pavlovsky, Y.; Gavriluk, E.; Zayachkivska, O. Influence of hydrogen sulphide-releasing Aspirin on mucosal integrity of esophageal and gastric mucosa. Proc. Shevchenko Sci. Soc. 2015, 43, 63–74. [Google Scholar]
- Kwiecien, S.; Magierowska, K.; Magierowski, M.; Surmiak, M.; Hubalewska-Mazgaj, M.; Pajdo, R.; Sliwowski, Z.; Chmura, A.; Wojcik, D.; Brzozowski, T. Role of sensory afferent nerves, lipid peroxidation and antioxidative enzymes in the carbon monoxide-induced gastroprotection against stress ulcerogensis. J. Physiol. Pharmacol. 2016, 67, 717–729. [Google Scholar] [PubMed]
- Abrahamovych, O.; Cherkas, A.; Abrahamovych, U.; Abrahamovych, M.; Serhiyenko, V. Heart Rate Variability: Physiological Bases, Clinical Importance and Peculiarities in Patients with Peptic Ulcer and after Resection of Stomach; Abrahamovych, O., Ed.; Danylo Halytskyi Lviv National Medical University: Lviv, Ukraine, 2014; ISBN 978-966-7175-729. [Google Scholar]
- O’Connor, A.; O’Morain, C.A.; Ford, A.C. Population screening and treatment of Helicobacter pylori infection. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-G.; Bae, J.-H.; Jastrzebski, Z.; Cherkas, A.; Heo, B.-G.; Gorinstein, S.; Ku, Y.-G. Binding, Antioxidant and Anti-proliferative Properties of Bioactive Compounds of Sweet Paprika (Capsicum annuum L.). Plant Foods Hum. Nutr. 2016, 71, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I. Glutathione Homeostasis and Functions: Potential Targets for Medical Interventions. J. Amino Acids 2012, 2012, 736837. [Google Scholar] [CrossRef] [PubMed]
- Semen, K.O.; Yelisyeyeva, O.P.; Kaminskyy, D.V.; Cherkas, A.P.; Zarkovic, K.; Lutsyk, O.; Cipak, A.; Jaganjac, M.; Zarkovic, N. Interval hypoxic training in complex treatment of Helicobacter pylori-associated peptic ulcer disease. Acta Biochim. Pol. 2010, 57, 199–208. [Google Scholar] [PubMed]
- Ielisieieva, O.P.; Semen, K.O.; Cherkas, A.P.; Kaminsky, D.V.; Kurkevych, A.K. Specific mechanisms of individually adjusted interval hypoxic hypercapnic effects on heart rate variability in athletes. Fiziol. Zhurnal 2007, 53, 78–86. [Google Scholar]
- Cherkas, A.; Golota, S. An intermittent exhaustion of the pool of glycogen in the human organism as a simple universal health promoting mechanism. Med. Hypotheses 2014, 82, 387–389. [Google Scholar] [CrossRef] [PubMed]
- Gulevsky, A.K.; Abakumova, Y.S.; Moiseyeva, N.N.; Ivanov, Y.G. Influence of Cord Blood Fraction (below 5 kDa) on Reparative Processes during Subchronic Ulcerative Gastropathy. Ulcers 2011, 2011, 214124. [Google Scholar] [CrossRef] [PubMed]
- Stelmakh, A.; Abrahamovych, O.; Cherkas, A. Highly purified calf hemodialysate (Actovegin®) may improve endothelial function by activation of proteasomes: A hypothesis explaining the possible mechanisms of action. Med. Hypotheses 2016, 95, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Cai, H.; Sasazuki, S.; Tsugane, S.; Zheng, W.; Cho, E.R.; Jee, S.H.; Michel, A.; Pawlita, M.; Xiang, Y.B.; et al. Fruit and vegetable consumption, Helicobacter pylori antibodies, and gastric cancer risk: A pooled analysis of prospective studies in China, Japan, and Korea. Int. J. Cancer 2017, 140, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Shafaghi, A.; Pourkazemi, A.; Khosravani, M.; Fakhrie Asl, S.; Amir Maafi, A.; Atrkar Roshan, Z.; Abaspour Rahimabad, J. The Effect of Probiotic Plus Prebiotic Supplementation on the Tolerance and Efficacy of Helicobacter Pylori Eradication Quadruple Therapy: A Randomized Prospective Double Blind Controlled Trial. Middle East J. Dig. Dis. 2016, 8, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Ingold, K.U.; Pratt, D.A. Advances in radical-trapping antioxidant chemistry in the 21st century: A kinetics and mechanisms perspective. Chem. Rev. 2014, 114, 9022–9046. [Google Scholar] [CrossRef] [PubMed]
- Gargallo, C.J.; Sostres, C.; Lanas, A. Prevention and Treatment of NSAID Gastropathy. Curr. Treat. Opt. Gastroenterol. 2014, 12, 398–413. [Google Scholar] [CrossRef] [PubMed]



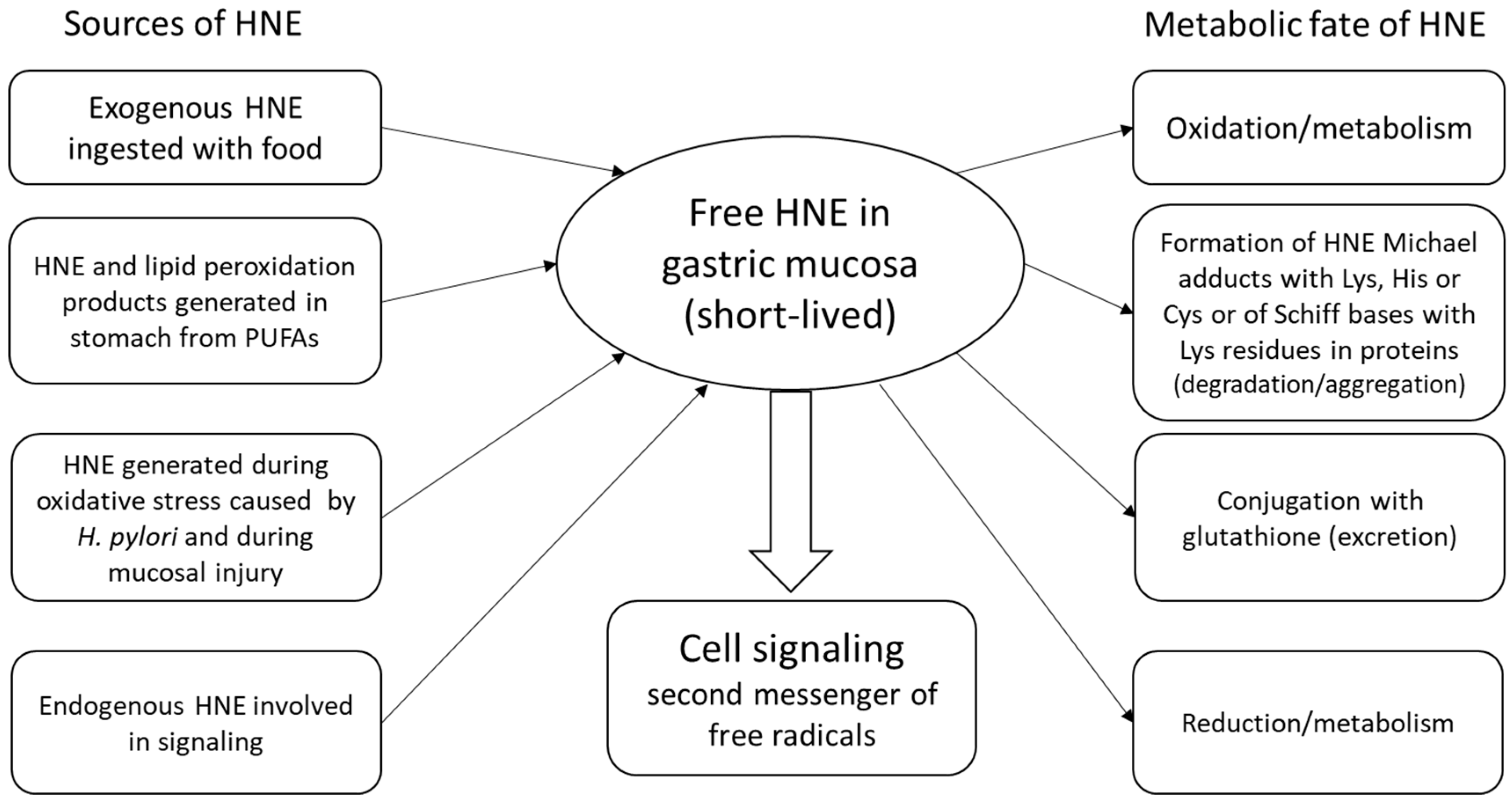{kind=link}
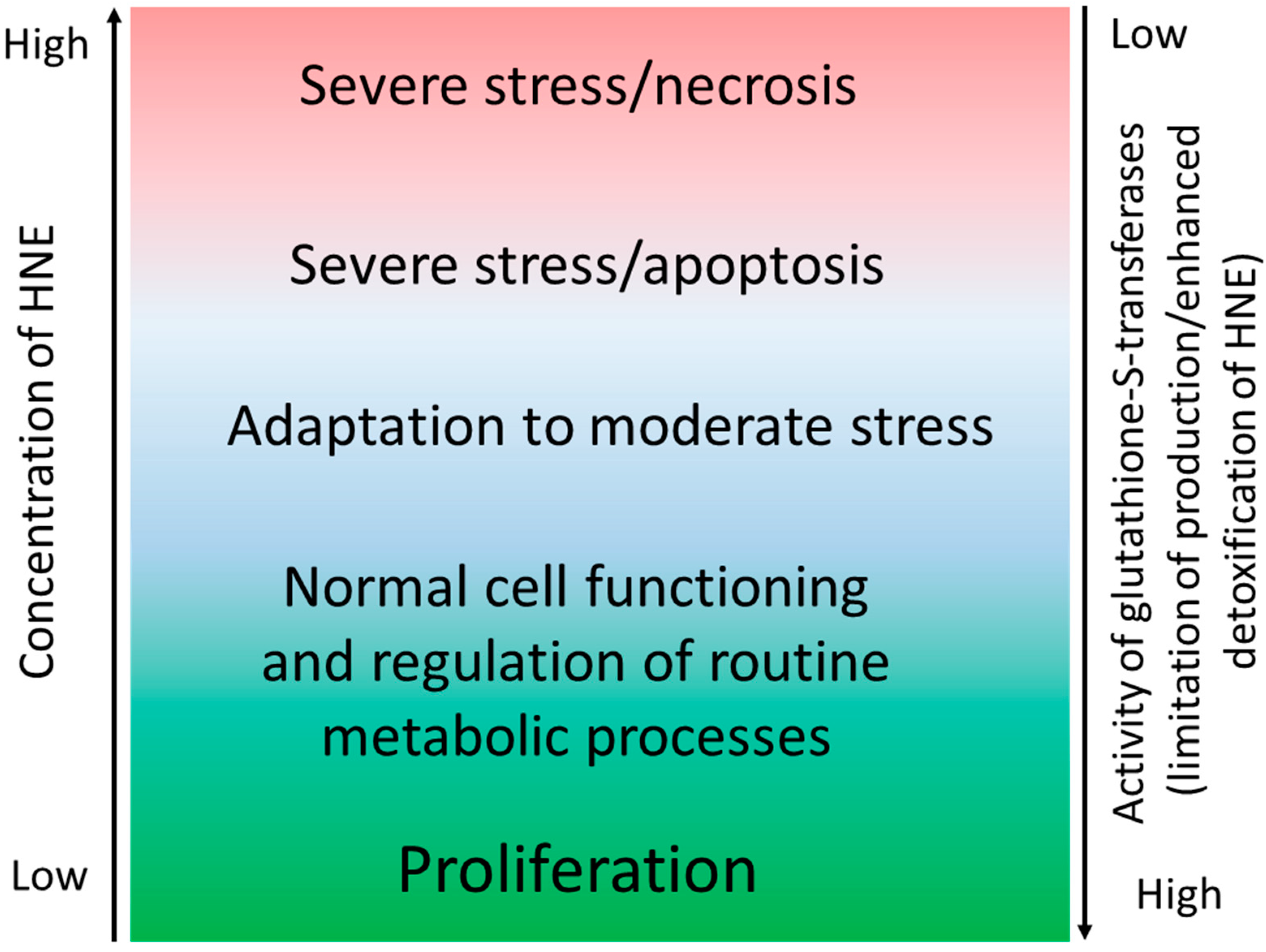{kind=link}
| Intervention | Target Process/Pharmacological Effect | References |
|---|---|---|
| Proton pump inhibitors, H2 histamine receptor inhibitors | Reduction of acidity, decreased proteolytic activity of gastric juice/decreased gastric injury (production of HNE) | [61,66] |
| Antibiotics | H. pylori eradication/decreased gastric injury (production of HNE) | [61,66] |
| NO, CO, H2S-releasing NSAIDs | Release of CO, NO, and/or H2S modulates redox signaling, improves endothelial function, and improves microcirculation/reduced production and improved utilization of HNE | [75,76] |
| Antioxidants/polyphenols present in food | Reduced lipid peroxidation of PUFAs in stomach/reduced absorption of exogenous HNE | [2,79] |
| Phytochemical and phytotoxins with moderate prooxidant action | Nrf-2 activators induce expression of antioxidant genes and increase detoxification of HNE | [20,80] |
| Interval hypoxic training | Improvement of autonomic control of microcirculation and function of internal organs | [81,82] |
| Exercise, intermittent fasting, caloric restriction | Activation of autophagy, reduction of systemic inflammatory response, improvement of protein quality control and autonomic regulation | [83] |
| Ulcer-healing drugs (actovegin, solcoseryl etc.) | Mechanism unknown, suggested influence on microcirculation and/or endothelial function | [84,85] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cherkas, A.; Zarkovic, N. 4-Hydroxynonenal in Redox Homeostasis of Gastrointestinal Mucosa: Implications for the Stomach in Health and Diseases. Antioxidants 2018, 7, 118. https://doi.org/10.3390/antiox7090118
Cherkas A, Zarkovic N. 4-Hydroxynonenal in Redox Homeostasis of Gastrointestinal Mucosa: Implications for the Stomach in Health and Diseases. Antioxidants. 2018; 7(9):118. https://doi.org/10.3390/antiox7090118
Chicago/Turabian StyleCherkas, Andriy, and Neven Zarkovic. 2018. "4-Hydroxynonenal in Redox Homeostasis of Gastrointestinal Mucosa: Implications for the Stomach in Health and Diseases" Antioxidants 7, no. 9: 118. https://doi.org/10.3390/antiox7090118
APA StyleCherkas, A., & Zarkovic, N. (2018). 4-Hydroxynonenal in Redox Homeostasis of Gastrointestinal Mucosa: Implications for the Stomach in Health and Diseases. Antioxidants, 7(9), 118. https://doi.org/10.3390/antiox7090118