Bis-allylic Deuterated DHA Alleviates Oxidative Stress in Retinal Epithelial Cells


 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Chemical Synthesis
2.3. Impact of Deuterium on DHA Toxicity and Free Radical Processes Involved in Cellular Lipid Peroxidation
2.3.1. Cell Culture
2.3.2. Cell Viability
2.3.3. Toxicity of Polyunsaturated Fatty Acid
2.3.4. Protection of Lipophenols against All-trans Retinal
2.3.5. Comparison of Lipid Peroxidation Status in ARPE-19 cells with C11-Bodipy581/591 under Oxidative Conditions
2.3.6. Fluorescence-activated Cell Sorting Analysis
2.3.7. Statistical Analyses for Cellular Biological Tests
2.4. Impact of Deuterium on DHA Oxidation in Non-cellular Media
2.4.1. Oxidation Method of Natural/Deuterated DHAs
2.4.2. Solid Phase Extraction of Oxidized Samples
2.4.3. Liquid Chromatography/Mass Spectrometry for Neuroprostane Analysis
2.4.4. Standard Solutions
2.5. Impact of Deuterium on DHA Enzymatic Oxidation by 15s-LOX
2.5.1. Kinetics Parameters of 15s-LOX
2.5.2. Liquid Chromatography/Mass Spectrometry of 15s-LOX Oxidized Sample
3. Results
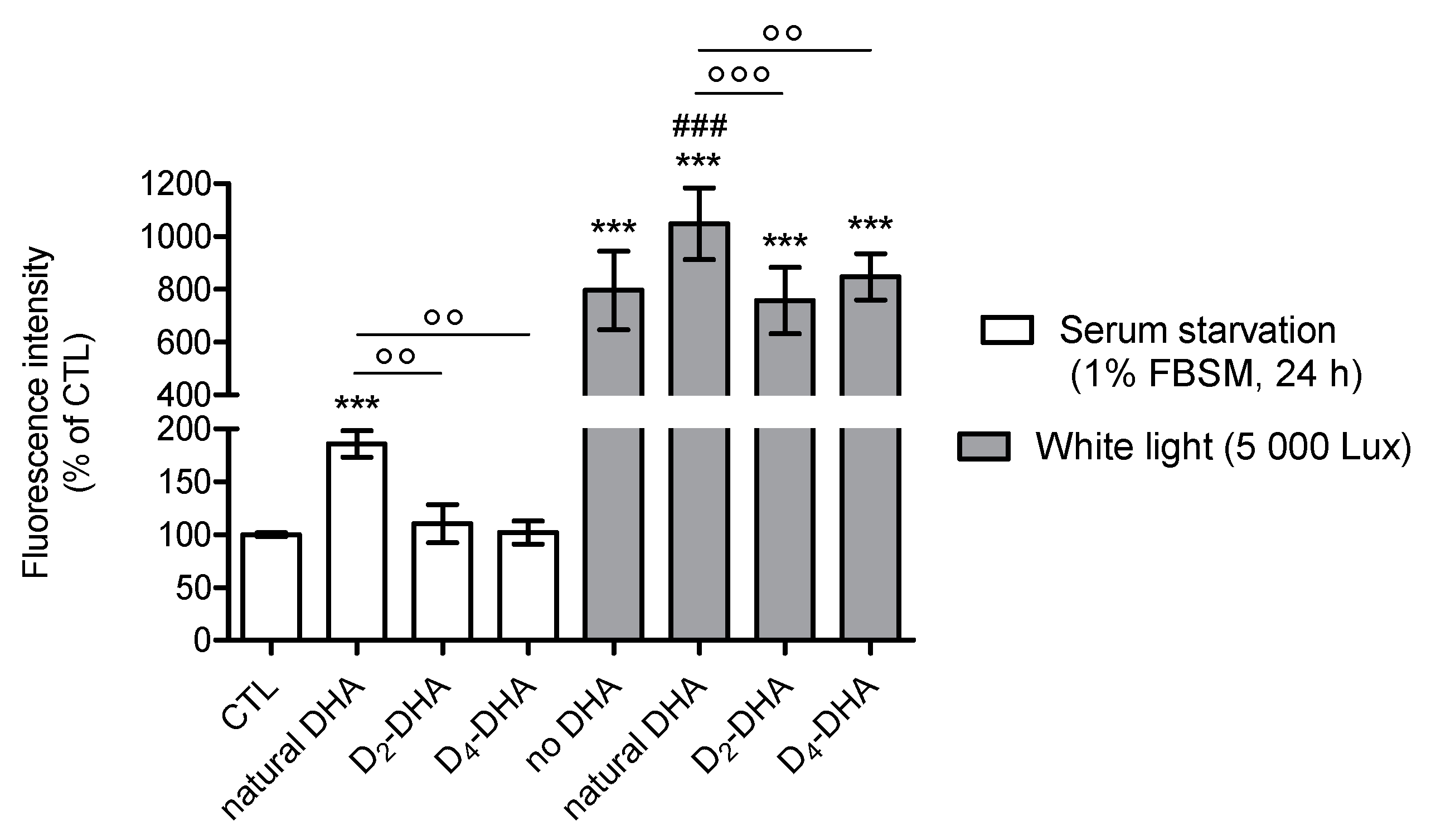
3.1. Deuterium Incorporation at Bis-allylic Positions Decreases DHA Toxicity on ARPE-19 Cell Line
3.2. Deuterium Incorporation at Bis-allylic Positions of DHA Reduces Radical Lipid Peroxidation Status on ARPE-19 Cell Line
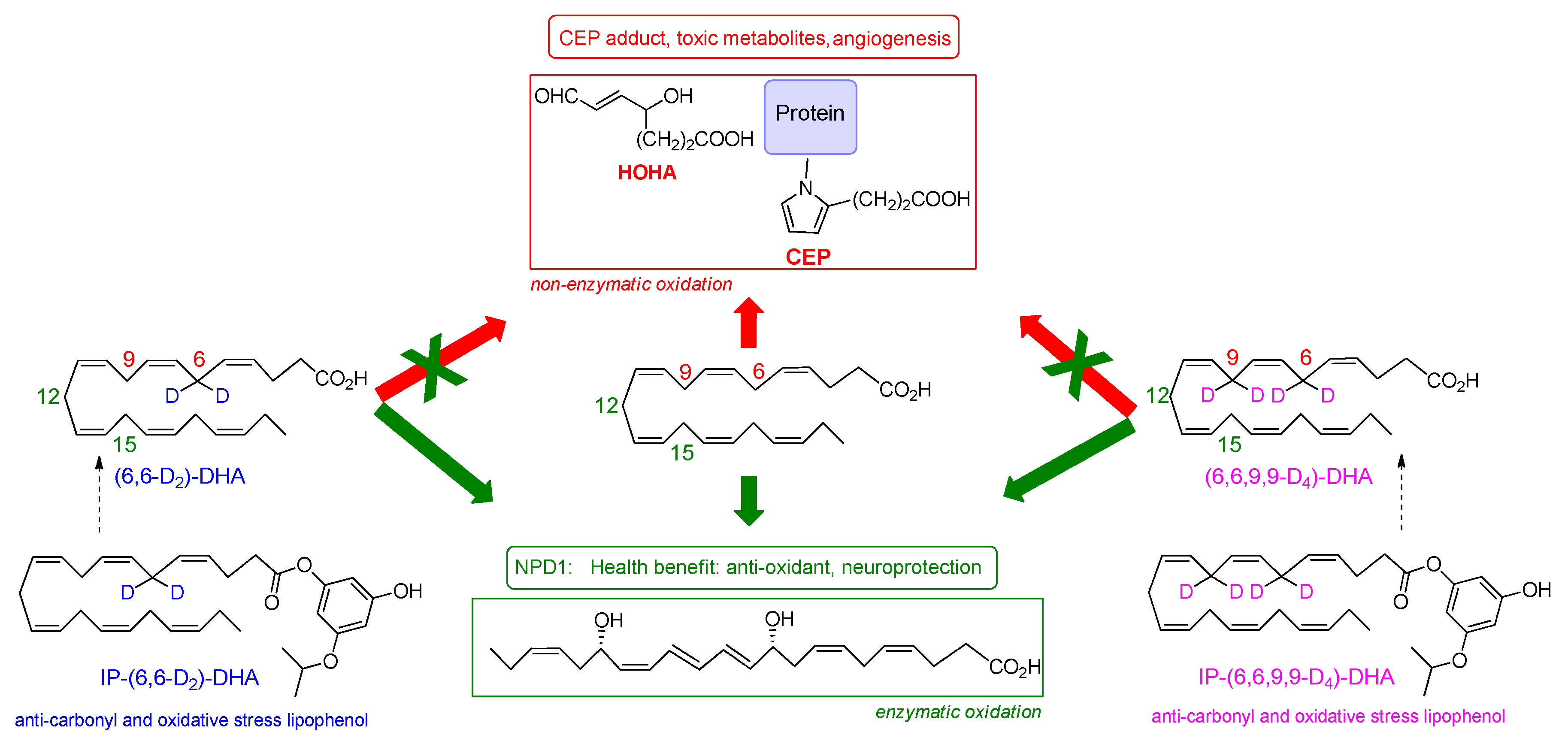
3.3. Deuterium Incorporation at Four Bis-allylic Positions of IP-DHA Lipophenol Reduces Lipid Peroxidation Status on ARPE-19 Cell Line
3.4. Deuterium Incorporation at Bis-allylic positions of IP-DHA Lipophenol Maintains Cytoprotection Against All-trans Retinal in ARPE-19 Cells
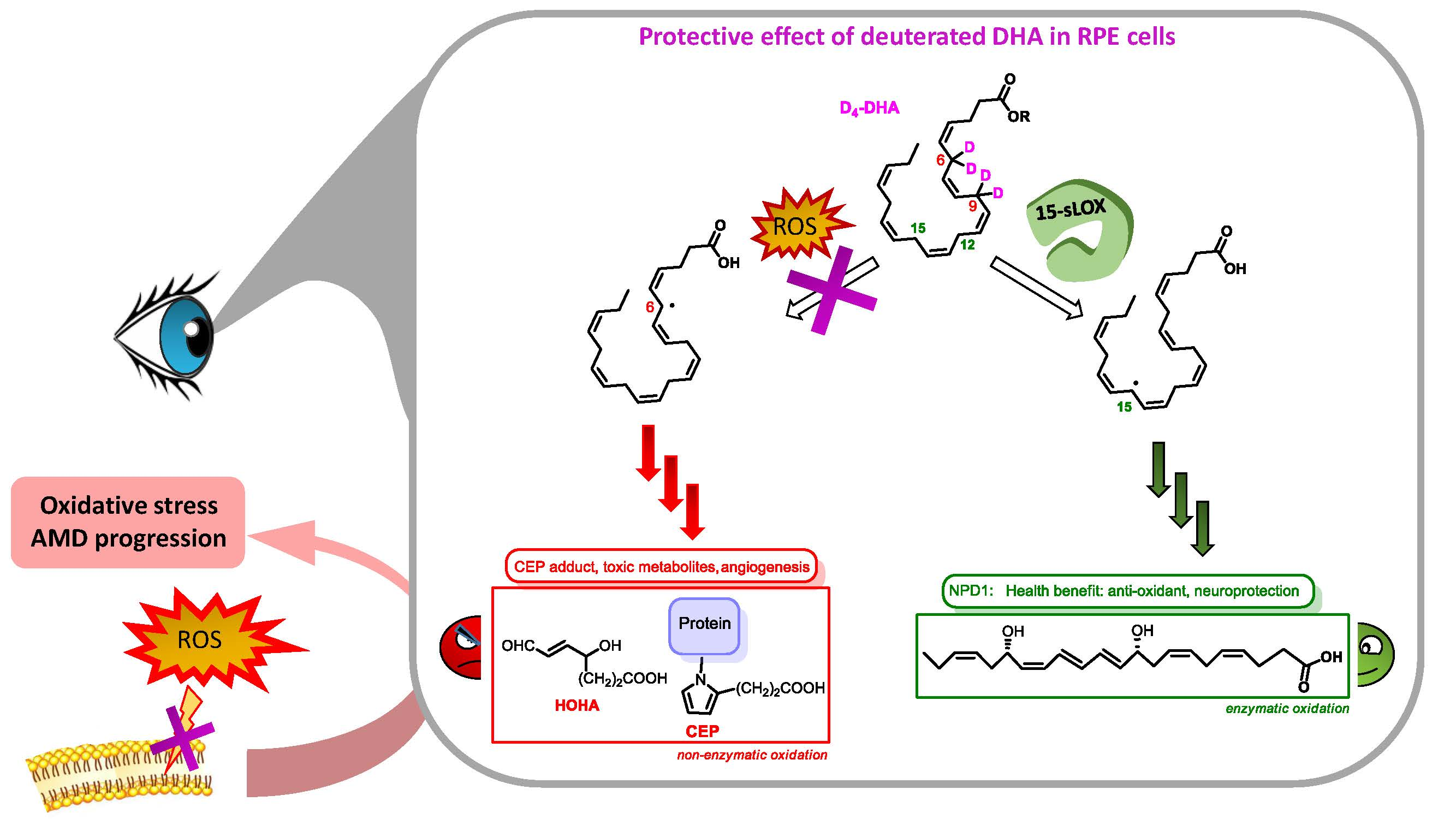
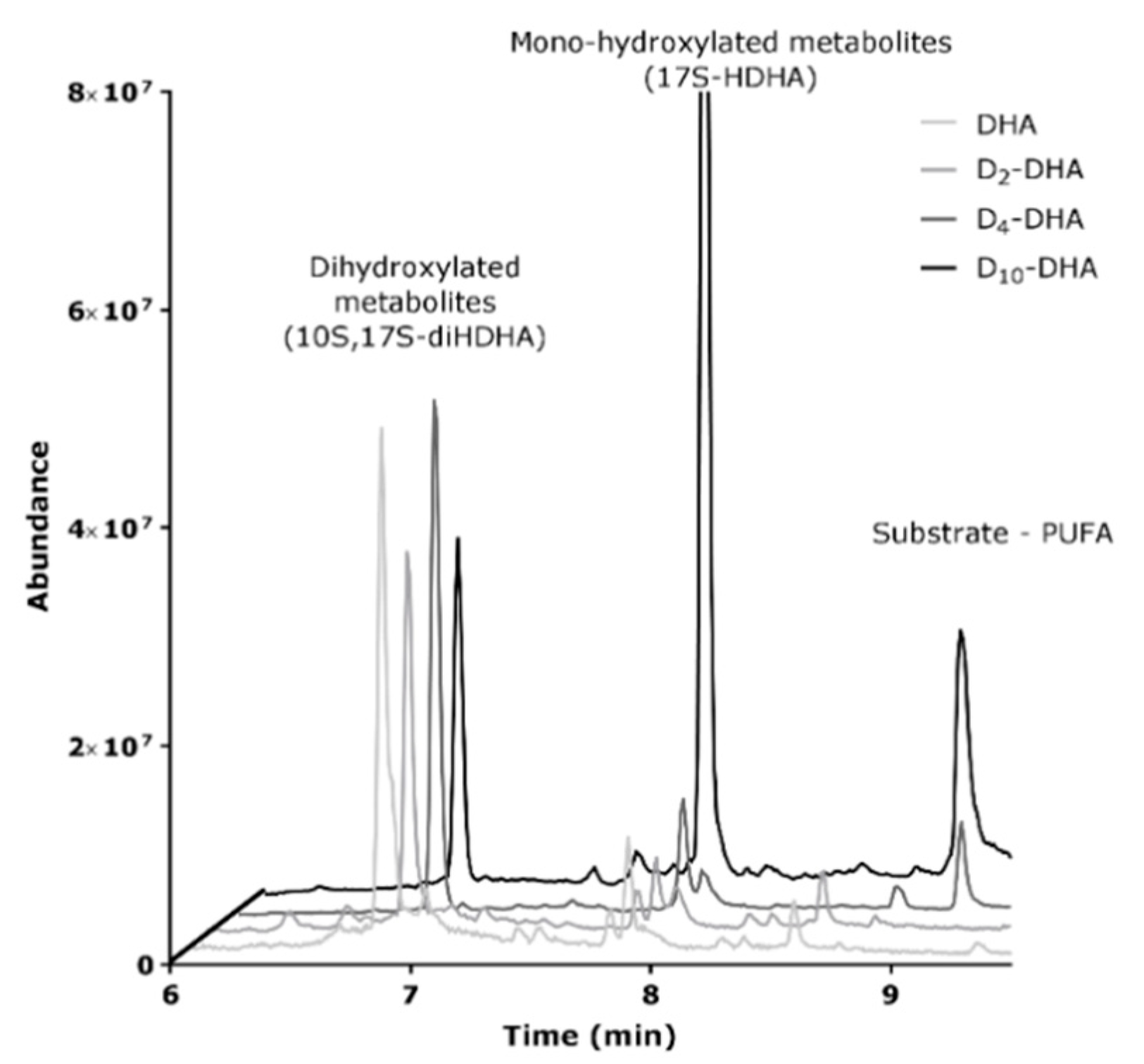
3.5. Deuterium Incorporation at Bis-allylic Positions of DHA Impacts the Formation of Metabolites from Radical Lipid Peroxidation
3.6. 15-sLOX Enzymatic Oxidation of Deuterated-DHA was not Inhibited by Deuterium Incorporation at the 6 and 9 Bis-allylic Positions
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, W.; Esselman, W.J.; Jump, D.B.; Busik, J.V. Anti-inflammatory effect of docosahexaenoic acid on cytokine-induced adhesion molecule expression in human retinal vascular endothelial cells. Invest. Ophthalmol. Vis. Sci. 2005, 46, 4342–4347. [Google Scholar] [CrossRef] [PubMed]
- Connor, K.M.; SanGiovanni, J.P.; Lofqvist, C.; Aderman, C.M.; Chen, J.; Higuchi, A.; Hong, S.; Pravda, E.A.; Majchrzak, S.; Carper, D.; et al. Increased dietary intake of ω-3-polyunsaturated fatty acids reduces pathological retinal angiogenesis. Nat. Med. 2007, 13, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Rotstein, N.P.; Politi, L.E.; German, O.L.; Girotti, R. Protective effect of docosahexaenoic acid on oxidative stress-induced apoptosis of retina photoreceptors. Invest. Ophthalmol. Vis. Sci. 2003, 44, 2252–2259. [Google Scholar] [CrossRef] [PubMed]
- Suh, M.; Wierzbicki, A.A.; Lien, E.; Clandinin, M.T. Relationship between dietary supply of long-chain fatty acids and membrane composition of long- and very long chain essential fatty acids in developing rat photoreceptors. Lipids 1996, 31, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Fliesler, A.J.; Anderson, R.E. Chemistry and metabolism of lipids in the vertebrate retina. Prog. Lipid Res. 1983, 22, 79–131. [Google Scholar] [CrossRef]
- Querques, G.; Forte, R.; Souied, E.H. Retina and Omega-3. J. Nutr. Metab. 2011, 2011, 74836. [Google Scholar] [CrossRef] [PubMed]
- Bazan, N.G.; Calandria, J.M.; Serhan, C.N. Rescue and repair during photoreceptor cell renewal mediated by docosahexaenoic acid-derived neuroprotectin D1. J. Lipid Res. 2010, 51, 2018–2031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazan, N.G.; Marcheselli, V.L.; Cole-Edwards, K. Brain response to injury and neurodegeneration: Endogenous neuroprotective signaling. Ann. N. Y. Acad. Sci. 2005, 1053, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Marcheselli, V.L.; Serhan, C.N.; Bazan, N.G. Neuroprotectin D1: A docosahexaenoic acid-derived docosatriene protects human retinal pigment epithelial cells from oxidative stress. Proc. Natl. Acad. Sci. USA 2004, 101, 8491–8496. [Google Scholar] [CrossRef] [Green Version]
- Bannenberg, G.L.; Chiang, N.; Ariel, A.; Arita, M.; Tjonahen, E.; Gotlinger, K.H.; Hong, S.; Serhan, C.N. Molecular circuits of resolution: Formation and actions of resolvins and protectins. J. Immunol. 2005, 174, 4345–4355. [Google Scholar] [CrossRef]
- Vistoli, G.; Maddis, D.D.; Cipak, A.; Zarkovic, N.; Carini, M.; Aldini, G. Advanced glycoxidation and lipoxidation end products (AGEs and ALEs): An overview of their mechanisms of formation. Free Radical Res. 2013, 47, 3–27. [Google Scholar] [CrossRef] [PubMed]
- Véricel, E.; Polette, A.; Bacot, S.; Calzada, C.; Lagarde, M. Pro- and antioxidant activities of docosahexaenoic acid on human blood platelets. J. Thromb. Haemost. 2003, 1, 566–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; Xu, L.; Porter, N.A. Free radical lipid peroxidation: Mechanisms and analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef] [PubMed]
- Semchyshyn, H.M. Reactive carbonyl species in vivo: Generation and dual biological effects. Sci. World J. 2014, 2014, 417842. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.Z. Oxidative stress, polyunsaturated fatty acids-derived oxidation products and bisretinoids as potential inducers of CNS diseases: Focus on age-related macular degeneration. Pharmacol. Rep. 2013, 65, 288–304. [Google Scholar] [CrossRef]
- Malinin, N.L.; West, X.Z.; Byzova, T.V. Oxidation as “The Stress of Life”. Aging (Albany NY) 2011, 3, 906–910. [Google Scholar] [CrossRef] [PubMed]
- Hollyfield, J.G. Age-Related Macular Degeneration: The Molecular Link between Oxidative Damage, Tissue-Specific Inflammation and Outer Retinal Disease: The Proctor Lecture. Invest. Ophthalmol. Vis. Sci. 2010, 51, 1276–1281. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Meer, S.G.; Miyagi, M.; Rayborn, M.E.; Hollyfield, J.G.; Crabb, J.W.; Salomon, R.G. Carboxyethylpyrrole Protein Adducts and Autoantibodies, Biomarkers for Age-related Macular Degeneration. J. Biol. Chem. 2003, 278, 42027–42035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahem, Q.; Renganathan, K.; Sears, J.; Vasanji, A.; Gu, X.; Lu, L.; Salomon, R.G.; Crabb, J.W.; Anand-Apte, B. Carboxyethylpyrrole oxidative protein modifications stimulate neovascularization: Implications for age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2006, 103, 13480–13484. [Google Scholar] [CrossRef] [Green Version]
- Hollyfield, J.G.; Perez, V.L.; Salomon, R.G. A Hapten Generated from an Oxidation Fragment of Docosahexaenoic Acid Is Sufficient to Initiate Age-Related Macular Degeneration. Mol. Neurobiol. 2010, 41, 290–298. [Google Scholar] [CrossRef] [Green Version]
- Crauste, C.; Vigor, C.; Brabet, P.; Picq, M.; Lagarde, M.; Hamel, C.; Durand, T.; Vercauteren, J. Synthesis and Evaluation of Polyunsaturated Fatty Acid-Phenol Conjugates as Anti-Carbonyl-Stress Lipophenols. Eur. J. Org. Chem. 2014, 4548–4561. [Google Scholar] [CrossRef]
- Donato, L.; Scimone, C.; Rinaldi, C.; Aragona, P.; Briuglia, S.; D’Ascola, A.; D’Angelo, R.; Sidoti, A. Stargardt Phenotype Associated with Two ELOVL4 Promoter Variants and ELOVL4 Downregulation: New Possible Perspective to Etiopathogenesis? Invest. Ophthalmol. Vis. Sci. 2018, 59, 843–857. [Google Scholar] [CrossRef] [PubMed]
- Cideciyan, A.V.; Aleman, T.S.; Swider, M.; Schwartz, S.B.; Steinberg, J.D.; Brucker, A.J.; Maguire, A.M.; Bennett, J.; Stone, E.M.; Jacobson, S.G. Mutations in ABCA4 result in accumulation of lipofuscin before slowing of the retinoid cycle: A reappraisal of the human disease sequence. Hum. Mol. Genet. 2004, 13, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, J.R.; Nakanishi, K.; Parish, C.A. The lipofuscin fluorophore A2E mediates blue light-induced damage to retinal pigmented epithelial cells. Invest. Ophthalmol. Vis. Sci. 2000, 41, 1981–1989. [Google Scholar]
- Sparrow, J.R.; Gregory-Roberts, E.; Yamamoto, K.; Blonska, A.; Ghosh, S.K.; Ueda, K.; Zhou, J. The bisretinoids of retinal pigment epithelium. Prog. Retin. Eye Res. 2012, 31, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Yoon, K.D.; Yamamoto, K.; Ueda, K.; Zhou, J.; Sparrow, J.R. A Novel Source of Methylglyoxal and Glyoxal in Retina: Implications for Age-Related Macular Degeneration. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Ben-Shabat, S.; Itagaki, Y.; Jockusch, S.; Sparrow, J.R.; Turro, N.J.; Nakanishi, K. Formation of a Nonaoxirane from A2E, a Lipofuscin Fluorophore related to Macular Degeneration, and Evidence of Singlet Oxygen Involvement. Angew. Chem. Int. Ed. 2002, 41, 814–817. [Google Scholar] [CrossRef]
- Chen, Y.; Okano, K.; Maeda, T.; Chauhan, V.; Golczak, M.; Maeda, A.; Palczewski, K. Mechanism of all-trans-retinal toxicity with implications for stargardt disease and age-related macular degeneration. J. Biol. Chem. 2012, 287, 5059–5069. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, K.; Zhang, K.; Zhou, F.; Zhu, L. Induction of oxidative and nitrosative stresses in human retinal pigment epithelial cells by all-trans-retinal. Exp. Cell Res. 2016, 348, 87–94. [Google Scholar] [CrossRef]
- Brabet, P.; Cubizolle, A.; Crauste, C.; Guillou, L.; Bonnefont, A.-L.; Vercauteren, J.; Durand, T.; Hamel, C.P. A phloroglucinol-DHA derivative protects against light-induced retinal degeneration in an Abca4-deficient mouse model. Invest. Ophthalmol. Vis. Sci. 2017, 58, 2031. [Google Scholar]
- New Lipophenol Compounds and Uses Thereof. Available online: https://patents.google.com/patent/WO2015162265A1/en (accessed on 24 April 2015).
- Schnebelen, C.; Viau, S.; Grégoire, S.; Joffre, C.; Creuzot-Garcher, C.P.; Bron, A.M.; Bretillon, L.; Acar, N. Nutrition for the Eye: Different Susceptibility of the Retina and the Lacrimal Gland to Dietary Omega-6 and Omega-3 Polyunsaturated Fatty Acid Incorporation. ORE 2009, 41, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, T.; Neokleous, A.; Nicolaou, D.; Sears, B. Pilot study for treating dry age-related macular degeneration (AMD) with high-dose omega-3 fatty acids. PharmaNutrition 2014, 2, 8–11. [Google Scholar] [CrossRef]
- Querques, G.; Benlian, P.; Chanu, B.; Leveziel, N.; Coscas, G.; Soubrane, G.; Souied, E.H. DHA supplementation for late onset Stargardt disease: NAT-3 study. Clin. Ophthalmol. 2010, 4, 575–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosell, M.; Villa, M.; Durand, T.; Galano, J.-M.; Vercauteren, J.; Crauste, C. Total Syntheses of Two bis-Allylic-Deuterated DHA Analogues. Asian J. Org. Chem. 2017, 6, 322–334. [Google Scholar] [CrossRef]
- Hill, S.; Hirano, K.; Shmanai, V.V.; Marbois, B.N.; Vidovic, D.; Bekish, A.V.; Kay, B.; Tse, V.; Fine, J.; Clarke, C.F.; et al. Isotope-reinforced polyunsaturated fatty acids protect yeast cells from oxidative stress. Free Radic. Biol. Med. 2011, 50, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.; Lamberson, C.R.; Xu, L.; To, R.; Tsui, H.S.; Shmanai, V.V.; Bekish, A.V.; Awad, A.M.; Marbois, B.N.; Cantor, C.R.; et al. Small amounts of isotope-reinforced polyunsaturated fatty acids suppress lipid autoxidation. Free Radic. Biol. Med. 2012, 53, 893–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berbée, J.F.P.; Mol, I.M.; Milne, G.L.; Pollock, E.; Hoeke, G.; Lütjohann, D.; Monaco, C.; Rensen, P.C.N.; van der Ploeg, L.H.T.; Shchepinov, M.S. Deuterium-reinforced polyunsaturated fatty acids protect against atherosclerosis by lowering lipid peroxidation and hypercholesterolemia. Atherosclerosis 2017, 264, 100–107. [Google Scholar] [CrossRef]
- Andreyev, A.Y.; Tsui, H.S.; Milne, G.L.; Shmanai, V.V.; Bekish, A.V.; Fomich, M.A.; Pham, M.N.; Nong, Y.; Murphy, A.N.; Clarke, C.F.; et al. Isotope-reinforced polyunsaturated fatty acids protect mitochondria from oxidative stress. Free Radic. Biol. Med. 2015, 82, 63–72. [Google Scholar] [CrossRef]
- Raefsky, S.M.; Furman, R.; Milne, G.; Pollock, E.; Axelsen, P.; Mattson, M.P.; Shchepinov, M.S. Deuterated polyunsaturated fatty acids reduce brain lipid peroxidation and hippocampal amyloid β-peptide levels, without discernable behavioral effects in an APP/PS1 mutant transgenic mouse model of Alzheimer’s disease. Neurobiol. Aging 2018, 66, 165–176. [Google Scholar] [CrossRef]
- Shchepinov, M.S.; Roginsky, V.A.; Brenna, J.T.; Molinari, R.J.; To, R.; Tsui, H.; Clarke, C.F.; Manning-Boğ, A.B. Chapter 31—Deuterium Protection of Polyunsaturated Fatty Acids against Lipid Peroxidation: A Novel Approach to Mitigating Mitochondrial Neurological Diseases. In Omega-3 Fatty Acids in Brain and Neurological Health; Watson, R.R., De Meester, F., Eds.; Academic Press: Boston, MA, USA, 2014; pp. 373–383. [Google Scholar]
- Oger, C.; Bultel-Ponce, V.; Guy, A.; Balas, L.; Rossi, J.C.; Durand, T.; Galano, J.M. The handy use of Brown’s P2-Ni catalyst for a skipped diyne deuteration: Application to the synthesis of a [D4]-labeled F4t-neuroprostane. Chem. Eur. J. 2010, 16, 13976–13980. [Google Scholar] [CrossRef]
- Guy, A.; Oger, C.; Heppekausen, J.; Signorini, C.; De Felice, C.; Furstner, A.; Durand, T.; Galano, J.M. Oxygenated Metabolites of n-3 Polyunsaturated Fatty Acids as Potential Oxidative Stress Biomarkers: Total Synthesis of 8-F3t -IsoP, 10-F4t -NeuroP and [D4]-10-F4t -NeuroP. Chemistry 2014, 20, 6374–6380. [Google Scholar] [CrossRef] [PubMed]
- Giera, M.; Ioan-Facsinay, A.; Toes, R.; Gao, F.; Dalli, J.; Deelder, A.M.; Serhan, C.N.; Mayboroda, O.A. Lipid and lipid mediator profiling of human synovial fluid in rheumatoid arthritis patients by means of LC–MS/MS. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2012, 1821, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, M.; Körner, A.; Kaussen, T.; Knausberg, U.; Gerber, C.; Hansmann, G.; Jónasdóttir, H.S.; Giera, M.; Mirakaj, V. Inhibition of neogenin fosters resolution of inflammation and tissue regeneration. J. Clin. Invest. 2018, 128, 4711–4726. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damage in vivo and in cell culture: How should you do it and what do the results mean? Br. J. Pharmacol. 2004, 142, 231–255. [Google Scholar] [CrossRef]
- Drummen, G.P.C.; van Liebergen, L.C.M.; Op den Kamp, J.A.F.; Post, J.A. C11-BODIPY581/591, an oxidation-sensitive fluorescent lipid peroxidation probe: (micro) spectroscopic characterization and validation of methodology. Free Radic. Biol. Med. 2002, 33, 473–490. [Google Scholar] [CrossRef]
- MacDonald, M.L.; Murray, I.V.J.; Axelsen, P.H. Mass spectrometric analysis demonstrates that BODIPY 581/591 C11 overestimates and inhibits oxidative lipid damage. Free Radic. Biol. Med. 2007, 42, 1392–1397. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Rebolledo, Á.E.; Martínez-Pastor, F.; Fernández-Santos, M.R.; Del Olmo, E.; Bisbal, A.; Ros-Santaella, J.L.; Garde, J.J. Comparison of the TBARS Assay and BODIPY C11 Probes for Assessing Lipid Peroxidation in Red Deer Spermatozoa. Reprod. Domest. Anim. 2010, 45, e360–e368. [Google Scholar] [CrossRef]
- German, O.L.; Agnolazza, D.L.; Politi, L.E.; Rotstein, N.P. Light, lipids and photoreceptor survival: Live or let die? Photochem. Photobiol. Sci. 2015, 14, 1737–1753. [Google Scholar] [CrossRef]
- Galano, J.-M.; Lee, Y.Y.; Oger, C.; Vigor, C.; Vercauteren, J.; Durand, T.; Giera, M.; Lee, J.C.-Y. Isoprostanes, neuroprostanes and phytoprostanes: An overview of 25years of research in chemistry and biology. Prog. Lipid Res. 2017, 68, 83–108. [Google Scholar] [CrossRef]
- Musiek, E.S.; Cha, J.K.; Yin, H.; Zackert, W.E.; Terry, E.S.; Porter, N.A.; Montine, T.J.; Morrow, J.D. Quantification of F-ring isoprostane-like compounds (F4-neuroprostanes) derived from docosahexaenoic acid in vivo in humans by a stable isotope dilution mass spectrometric assay. J. Chromatogr. B 2004, 799, 95–102. [Google Scholar] [CrossRef]
- Tanito, M.; Brush, R.S.; Elliott, M.H.; Wicker, L.D.; Henry, K.R.; Anderson, R.E. High levels of retinal membrane docosahexaenoic acid increase susceptibility to stress-induced degeneration. J. Lipid Res. 2009, 50, 807–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliwell, B. Oxidative stress in cell culture: An under-appreciated problem? FEBS Lett. 2003, 540, 3–6. [Google Scholar] [CrossRef]
- Aryan, N.; Betts-Obregon, B.S.; Perry, G.; Tsin, A.T. Oxidative Stress Induces Senescence in Cultured RPE Cells. Open Neurol. J. 2016, 10, 83–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, D.; Wu, Y.; Ji, B. Docosahexaenoic acid aggravates photooxidative damage in retinal pigment epithelial cells via lipid peroxidation. J. Photochem. Photobiol. B 2014, 140, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Johansson, I.; Monsen, V.T.; Pettersen, K.; Mildenberger, J.; Misund, K.; Kaarniranta, K.; Schønberg, S.; Bjørkøy, G. The marine n-3 PUFA DHA evokes cytoprotection against oxidative stress and protein misfolding by inducing autophagy and NFE2L2 in human retinal pigment epithelial cells. Autophagy 2015, 11, 1636–1651. [Google Scholar] [CrossRef]
- Brunk, U.T.; Wihlmark, U.; Wrigstad, A.; Roberg, K.; Nilsson, S.-E. Accumulation of lipofuscin within retinal pigment epithelial cells results in enhanced sensitivity to photo-oxidation. Gerontology 1995, 41, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Joumard-Cubizolles, L.; Lee, J.C.-Y.; Vigor, C.; Leung, H.H.; Bertrand-Michel, J.; Galano, J.-M.; Mazur, A.; Durand, T.; Gladine, C. Insight into the contribution of isoprostanoids to the health effects of omega 3 PUFAs. Prostag. Other Lipid Mediat. 2017, 133, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Shchepinov, M.S. Reactive Oxygen Species, Isotope Effect, Essential Nutrients, and Enhanced Longevity. Rejuvenation Res. 2007, 10, 47–60. [Google Scholar] [CrossRef]
- Glickman, M.H.; Wiseman, J.S.; Klinman, J.P. Extremely Large Isotope Effects in the Soybean Lipoxygenase-Linoleic Acid Reaction. J. Am. Chem. Soc. 1994, 116, 793–794. [Google Scholar] [CrossRef]
- Navratil, A.R.; Shchepinov, M.S.; Dennis, E.A. Lipidomics Reveals Dramatic Physiological Kinetic Isotope Effects during the Enzymatic Oxygenation of Polyunsaturated Fatty Acids Ex Vivo. J. Am. Chem. Soc. 2018, 140, 235–243. [Google Scholar] [CrossRef]
- Donato, L.; Scimone, C.; Nicocia, G.; D’Angelo, R.; Sidoti, A. Role of oxidative stress in Retinitis pigmentosa: New involved pathways by an RNA-Seq analysis. Cell Cycle 2019, 18, 84–104. [Google Scholar] [CrossRef] [PubMed]
- Donato, L.; Scimone, C.; Nicocia, G.; Denaro, L.; Robledo, R.; Sidoti, A.; D’Angelo, R. GLO1 gene polymorphisms and their association with retinitis pigmentosa: A case–control study in a Sicilian population. Mol. Biol. Rep. 2018, 45, 1349–1355. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pro-oxidant Conditions | IC50 (µM) | ||
|---|---|---|---|
| Natural DHA | D2-DHA | D4-DHA | |
| Serum starvation (1% FBSM) | 65.21 ± 1.03 | 69.48 ± 1.46 | 75.60 ± 1.05 |
| H2O2 (600 µM) | 65.47 ± 1.87 | 66.98 ± 1.89 | 76.24 ± 2.26 |
| Substance | KM | VMax |
|---|---|---|
| Natural DHA | 0.0680 | 0.0160 |
| D2-DHA | 0.0363 | 0.0085 |
| D4-DHA | 0.0574 | 0.0124 |
| D10-DHA | 0.00327 | 0.0007 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosell, M.; Giera, M.; Brabet, P.; Shchepinov, M.S.; Guichardant, M.; Durand, T.; Vercauteren, J.; Galano, J.-M.; Crauste, C. Bis-allylic Deuterated DHA Alleviates Oxidative Stress in Retinal Epithelial Cells. Antioxidants 2019, 8, 447. https://doi.org/10.3390/antiox8100447
Rosell M, Giera M, Brabet P, Shchepinov MS, Guichardant M, Durand T, Vercauteren J, Galano J-M, Crauste C. Bis-allylic Deuterated DHA Alleviates Oxidative Stress in Retinal Epithelial Cells. Antioxidants. 2019; 8(10):447. https://doi.org/10.3390/antiox8100447
Chicago/Turabian StyleRosell, Mélissa, Martin Giera, Philippe Brabet, Mikhail S. Shchepinov, Michel Guichardant, Thierry Durand, Joseph Vercauteren, Jean-Marie Galano, and Céline Crauste. 2019. "Bis-allylic Deuterated DHA Alleviates Oxidative Stress in Retinal Epithelial Cells" Antioxidants 8, no. 10: 447. https://doi.org/10.3390/antiox8100447
APA StyleRosell, M., Giera, M., Brabet, P., Shchepinov, M. S., Guichardant, M., Durand, T., Vercauteren, J., Galano, J. -M., & Crauste, C. (2019). Bis-allylic Deuterated DHA Alleviates Oxidative Stress in Retinal Epithelial Cells. Antioxidants, 8(10), 447. https://doi.org/10.3390/antiox8100447