An Aromatic Aldehyde Synthase Controls the Synthesis of Hydroxytyrosol Derivatives Present in Virgin Olive Oil


Abstract
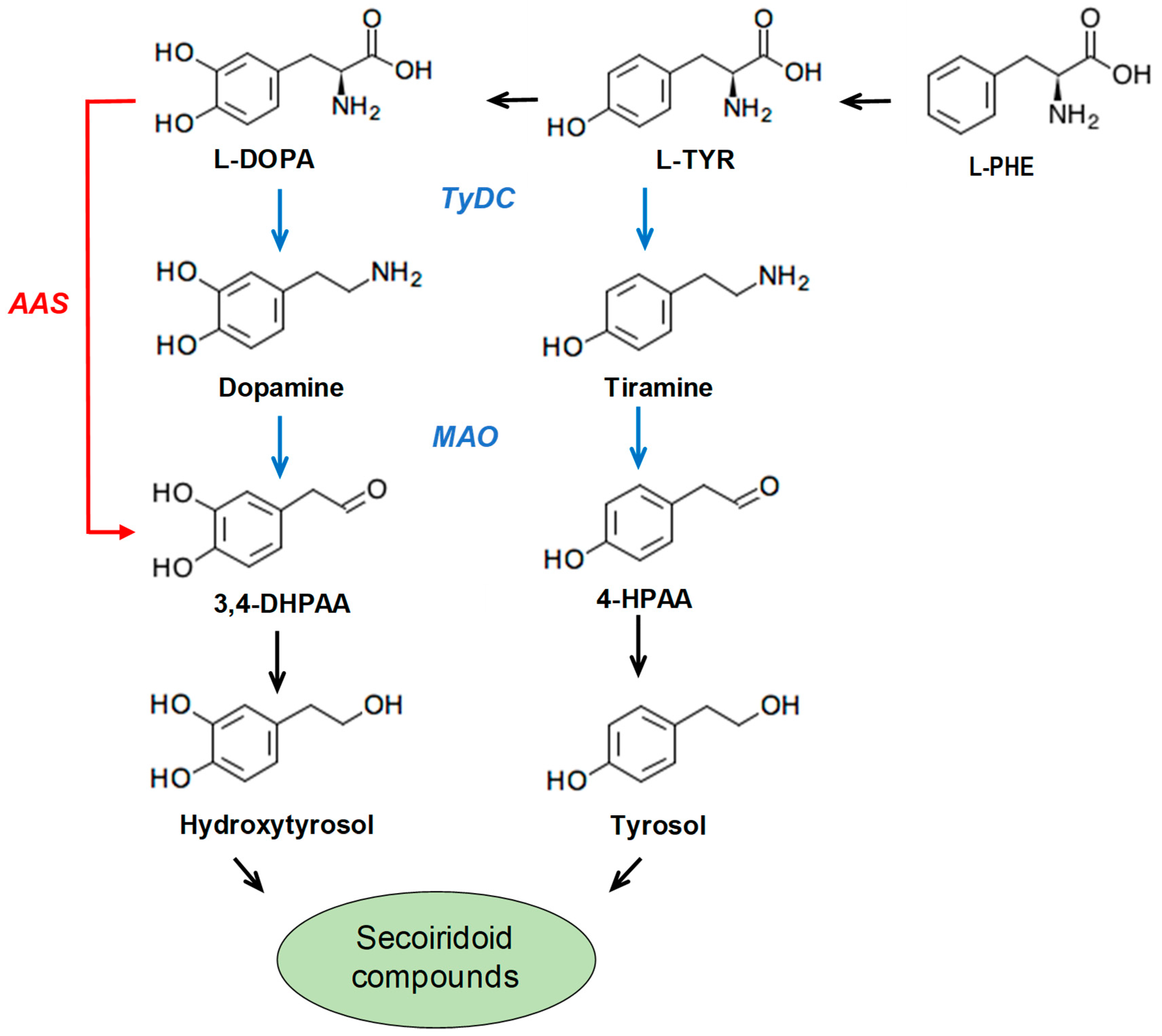
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. RNA Extraction, cDNA Library Construction, and Sequencing
2.3. Isolation of An AAS Full-Length cDNA Clone
2.4. Gene Expression Analysis by RT-QPCR
2.5. Heterologous Expression of OeAAS in Escherichia coli and Purification of OeAAS Protein
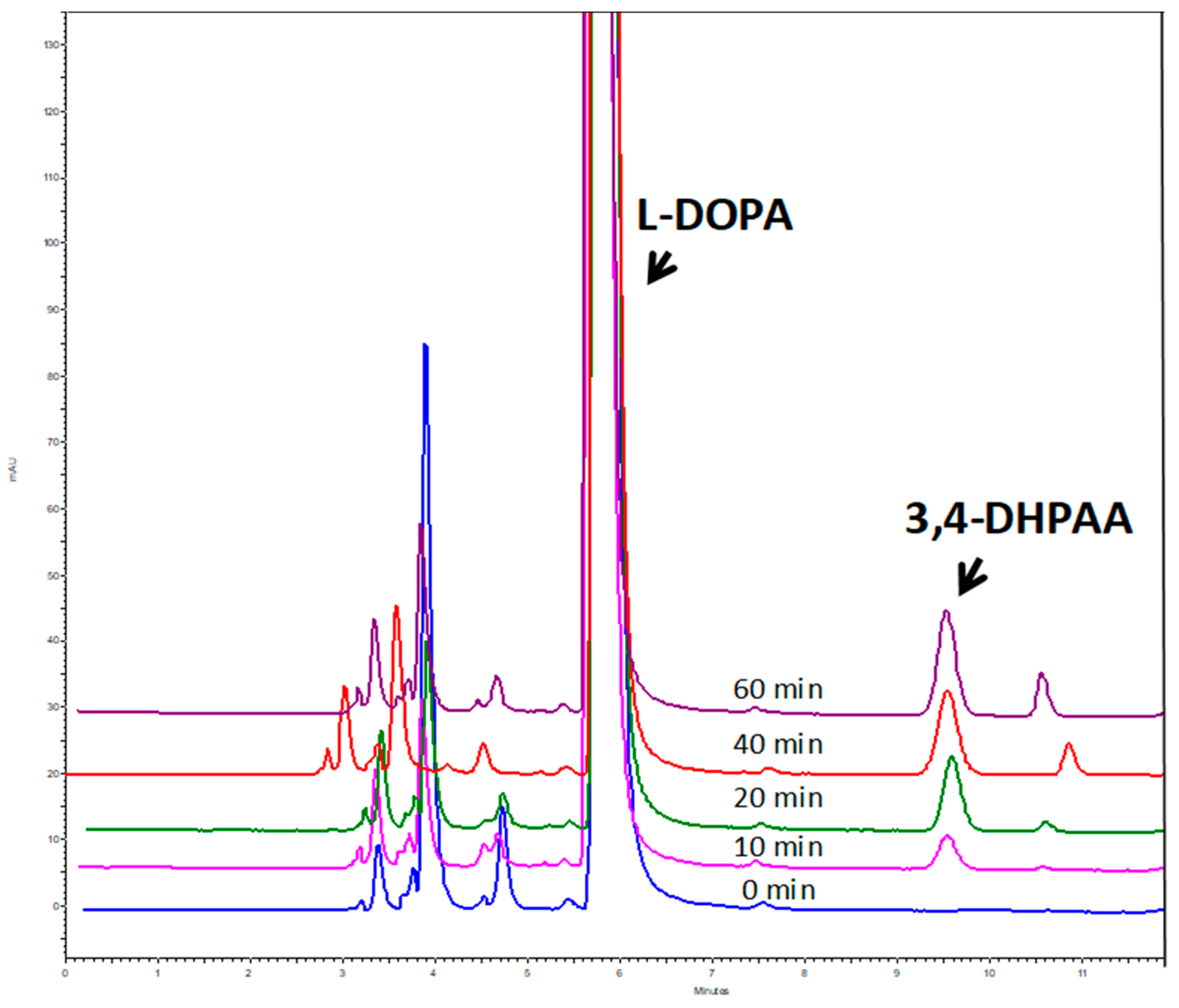
2.6. OeAAS Activity Assay
2.7. Sequence Alignment and Phylogenetic Analysis
2.8. Olive Oil Extraction
2.9. Extraction and Analysis of Fruit and VOO Phenolic Compounds
2.10. Extraction and Analysis of Free Amino Acids in Olive Fruit
3. Results and Discussion
3.1. Identification and Molecular Characterization of An AAS Gene in Olive
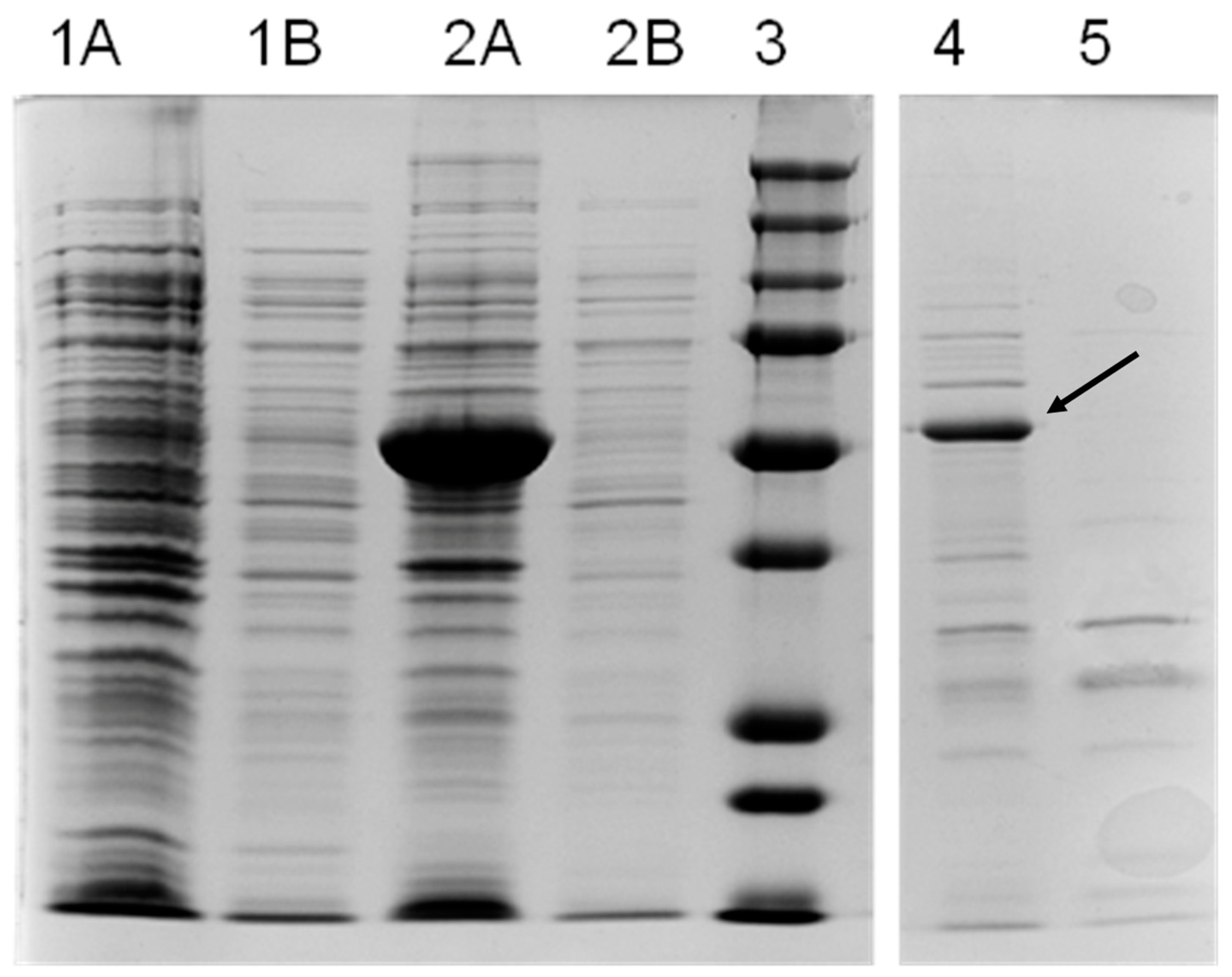
3.2. Purification and Biochemical Characterization of OeAAS
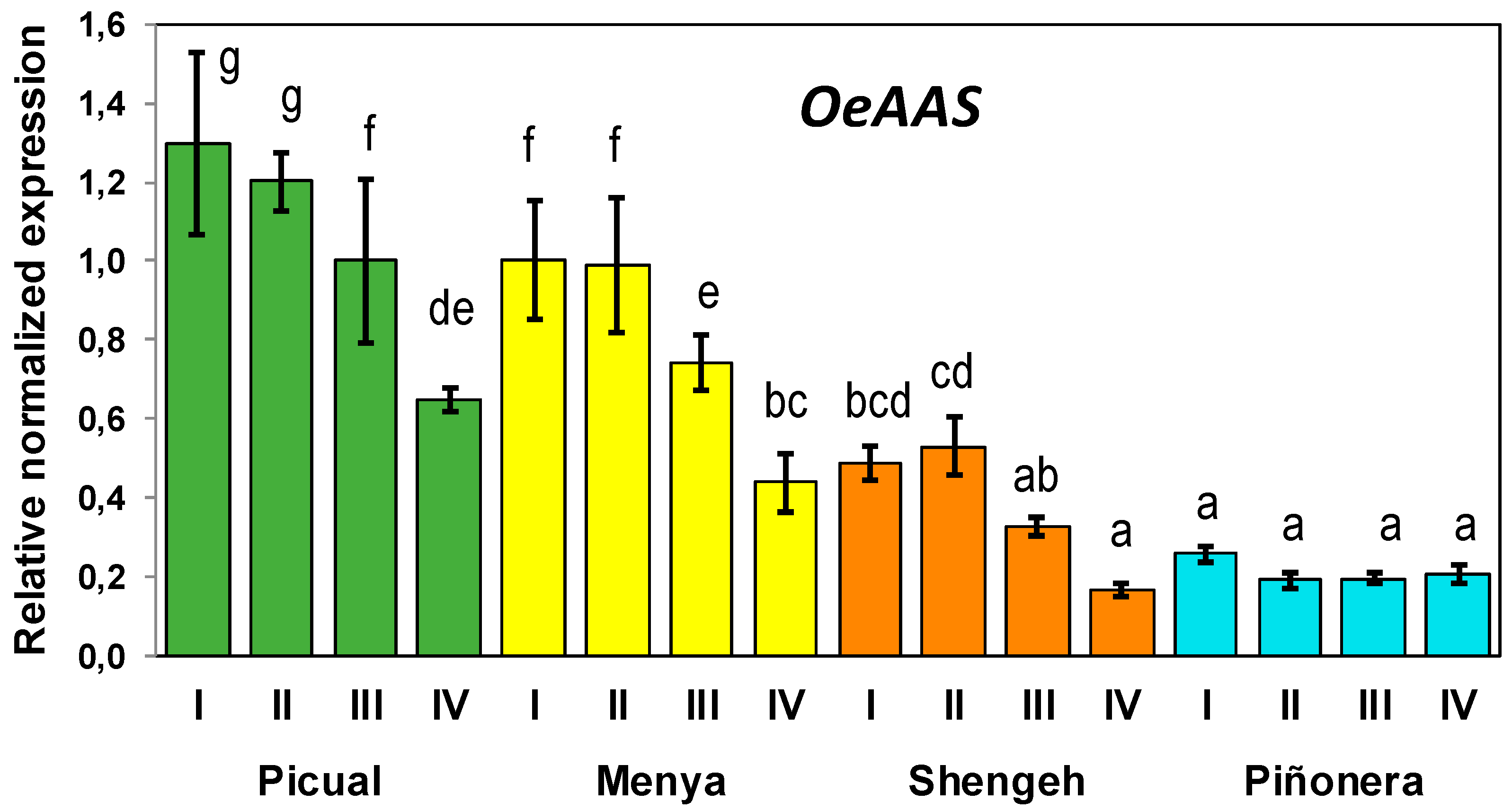
3.3. Developmental Expression of Olive AAS Gene Is Cultivar Dependent
3.4. Correlation of OeAAS Expression with Phenolic Accumulation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andrewes, P.; Busch, J.L.H.C.; de Joode, T.; Groenewegen, A.; Alexandre, H. Sensory properties of virgin olive oil polyphenols: Identification of deacetoxy-ligstroside glycon as a key contributor to pungency. J. Agric. Food Chem. 2003, 51, 1415–1420. [Google Scholar] [CrossRef] [PubMed]
- Mateos, R.; Cert, A.; Pérez-Camino, M.C.; García, J.M. Evaluation of virgin olive oil bitterness by quantification of secoiridoid derivatives. J. Am. Oil Chem. Soc. 2004, 81, 71–75. [Google Scholar] [CrossRef]
- Konstantinidou, V.; Covas, M.I.; Muñoz-Aguayo, D.; Khymenets, O.; de La Torre, R.; Saez, G.; Tormos, M.C.; Toledo, E.; Marti, A.; Ruiz-Gutiérrez, V.; et al. In vivo nutrigenomic effects of virgin olive oil polyphenols within the frame of the Mediterranean diet: A randomized controlled trial. FASEB J. 2010, 24, 2546–2557. [Google Scholar] [CrossRef] [PubMed]
- Serreli, G.; Deiana, M. Biological Relevance of Extra Virgin Olive Oil Polyphenols Metabolites. Antioxidants 2018, 7, 170. [Google Scholar] [CrossRef] [PubMed]
- García-Rodríguez, R.; Romero-Segura, C.; Sanz, C.; Sánchez-Ortiz, A.; Pérez, A. Role of polyphenol oxidase and peroxidase in shaping the phenolic profile of virgin olive oil. Food Res. Int. 2011, 44, 629–635. [Google Scholar] [CrossRef]
- Romero-Segura, C.; García-Rodrıguez, R.; Sánchez-Ortiz, A.; Sanz, C.; Pérez, A.G. The role of olive β-glucosidase in shaping the phenolic profile of virgin olive oil. Food Res. Int. 2012, 45, 191–196. [Google Scholar] [CrossRef]
- Fregapane, G.; Salvador, M.D. Fate and Prediction of Phenolic Secoiridoid Compounds throughout the Different Stages of the Virgin Olive Oil Making Process. Antioxidants 2017, 6, 61. [Google Scholar] [CrossRef] [PubMed]
- García-Rodríguez, R.; Belaj, A.; Romero-Segura, C.; Sanz, C.; Pérez, A.G. Exploration of genetic resources to improve the functional quality of virgin olive oil. J. Funct. Foods 2017, 38, 1–8. [Google Scholar] [CrossRef]
- Bernini, R.; Crisante, F.; Merendino, N.; Molinari, R.; Soldatelli, M.C.; Velotti, F. Synthesis of a novel ester of hydroxytyrosol and α-lipoic acid exhibiting an antiproliferative effect on human colon cancer HT-29 cells. Eur. J. Med. Chem. 2011, 46, 439–446. [Google Scholar] [CrossRef]
- Pinto, J.; Paiva-Martins, F.; Corona, G.; Debnam, E.S.; Oruna-Concha, M.J.; Vauzour, D.; Gordon, M.H.; Spencer, J.P. Absorption and metabolism of olive oil secoiridoids in the small intestine. British J. Nutr. 2011, 105, 1607–1618. [Google Scholar] [CrossRef] [Green Version]
- Lopez de las Hazas, M.-C.; Piňol, C.; Macia, A.; Romero, M.-P.; Pedret, A.; Solà, R.; Rubio, L.; Motilva, M.J. Differential absorption and metabolism of hydroxytyrosol and its precursors oleuropein and secoiridoids. J. Funct. Foods 2016, 22, 52–63. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Commission Regulation (EU) No 1018/2013 amending Regulation (EU) No 432/2012 establishing a list of permitted health claims made on foods other than those referring to the reduction of disease risk and to children’s development and heal. Off. J. Eur. Union L 2012, 282, 43–45. [Google Scholar]
- Obied, H.K.; Prenzler, P.D.; Ryan, D.; Servili, M.; Taticchi, A.; Esposto, S.; Robards, K. Biosynthesis and biotransformations of phenol-conjugated oleosidic secoiridoids from Olea europaea L. Nat. Prod. Rep. 2008, 25, 1167–1179. [Google Scholar] [CrossRef] [PubMed]
- Allouche, N.; Sayadi, S. Synthesis of hydroxytyrosol, 2-hydroxyphenylacetic acid, and 3-hydroxyphenylacetic acid by differential conversion of tyrosol isomers using Serratia marcescens strain. J. Agric. Food Chem. 2005, 53, 6525–6530. [Google Scholar] [CrossRef] [PubMed]
- Orenes-Pinero, E.; Garcia-Carmona, F.; Sanchez-Ferrer, A. A new process for obtaining hydroxytyrosol using transformed Escherichia coli whole cells with phenol hydroxylase gene from Geobacillus thermoglucosidasius. Food Chem. 2013, 139, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Chung, D.; Kim, S.Y.; Ahn, J.-H. Production of three phenylethanoids, tyrosol, hydroxytyrosol, and salidroside, using plant genes expressing in Escherichia coli. Sci. Rep. 2017, 7, 2578. [Google Scholar] [CrossRef]
- Li, C.; Jia, P.; Bai, Y.; Fan, T.; Zheng, X.; Cai, Y. Efficient Synthesis of Hydroxytyrosol from l-3,4- Dihydroxyphenylalanine Using Engineered Escherichia coli Whole Cells. J. Agric. Food Chem. 2019, 67, 6867–6873. [Google Scholar] [CrossRef]
- Lan, X.; Chang, K.; Zeng, L.; Liu, X.; Qiu, F.; Zheng, W.; Quan, H.; Liao, Z.; Chen, M.; Huang, W.; et al. Engineering salidroside biosynthetic pathway in hairy root cultures of Rhodiola crenulata based on metabolic characterization of tyrosine decarboxylase. PLoS ONE 2013, 8, e75459. [Google Scholar] [CrossRef]
- Torrens-Spence, M.P.; Pluskal, T.; Li, F.-S.; Carballo, V.; Weng, J.-K. Complete Pathway Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis. Mol. Plant. 2018, 11, 205–217. [Google Scholar] [CrossRef]
- Facchini, P.J.; Huber-Allanach, K.L.; Tari, L.W. Plant aromatic l-amino acid decarboxylases: Evolution, biochemistry, regulation, and metabolic engineering applications. Phytochemistry 2000, 54, 121–138. [Google Scholar] [CrossRef]
- Kaminaga, Y.; Schnepp, J.; Peel, G.; Kish, C.M.; Ben-Nissan, G.; Weiss, D.; Orlova, I.; Lavie, O.; Rhodes, D.; Wood, K.; et al. Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation. J. Biol. Chem. 2006, 281, 23357–23366. [Google Scholar] [CrossRef] [PubMed]
- De Luca, V.; Marineau, C.; Brisson, N. Molecular cloning and analysis of cDNA encoding a plant tryptophan decarboxylase: Comparison with animal dopa decarboxylases. Proc. Natl. Acad. Sci. USA 1989, 86, 2582–2586. [Google Scholar] [CrossRef] [PubMed]
- Torrens-Spence, M.P.; Liu, P.; Ding, H.; Harich, K.; Gillaspy, G.; Li, J. Biochemical evaluation of the decarboxylation and decarboxylation-deamination activities of plant aromatic amino acid decarboxylases. J. Biol. Chem. 2013, 288, 2376–2387. [Google Scholar] [CrossRef] [PubMed]
- Torrens-Spence, M.P.; Lazear, M.; von Guggenberg, R.; Ding, H.; Li, J. Investigation of a substrate-specifying residue within Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases. Phytochemistry 2014, 106, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Belaj, A.; Domínguez-Garcia, M.C.; Atienza, S.G.; Martin-Urdiroz, N.; de la Rosa, R.; Satovic, Z.; del Río, C. Developing a core collection of olive (Olea europaea L.) based on molecular markers (DArTs, SSRs, SNPs) and agronomic traits. Tree Genet. Genomes 2012, 8, 365–378. [Google Scholar] [CrossRef]
- Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; MacManes, M.D.; Ott, M.; Orvis, J.; Pochet, N.; Strozzi, F.; Weeks, N.; et al. De novo transcript sequence reconstruction from RNA-Seq: Reference generation and analysis with trinity. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef]
- Nonis, A.; Vezzaro, A.; Ruperti, B. Evaluation of RNA extraction methods and identification of putative reference genes for real-time quantitative polymerase chain reaction expression studies on Olive (Olea europaea L.) fruits. J. Agric. Food Chem. 2012, 60, 6855–6865. [Google Scholar] [CrossRef]
- Ben ali, S.; Guasmi, F.; Mohamed, M.B.; Benhaj, K.; Boussora, F.; Triki, T.; Kammoun, N.G. Identification of internal control genes for gene expression studies in olive mesocarp tissue during fruit ripening. S. Afr. J. Bot. 2018, 117, 11–16. [Google Scholar] [CrossRef]
- Romero-Segura, C.; Sanz, C.; Pérez, A.G. Purification and characterization of an olive fruit β-glucosidase involved in the biosynthesis of virgin olive oil phenolics. J. Agric. Food Chem. 2009, 57, 7983–7988. [Google Scholar] [CrossRef] [PubMed]
- Pérez, A.G.; León, L.; Pascual, M.; Romero-Segura, C.; Sánchez-Ortiz, A.; de la Rosa, R.; Sanz, C. Variability of virgin olive oil phenolic compounds in a segregating progeny from a single cross in Olea europaea L. and sensory and nutritional quality implications. PLoS ONE 2014, 9, e92898. [Google Scholar] [CrossRef] [PubMed]
- Pérez, A.G.; León, L.; Sanz, C.; De la Rosa, R. Fruit Phenolic Profiling: A New Selection Criterion in Olive Breeding Programs. Front. Plant Sci. 2018, 8, 241. [Google Scholar] [CrossRef] [PubMed]
- Alaiz, M.; Navarro, J.L.; Girón, J.; Vioque, E. Amino acid analysis by high-performance liquid chromatography after derivatization with diethyl ethoxymethylenemalonate. J. Chromatogr. 1992, 591, 181–186. [Google Scholar] [CrossRef]
- Torrens-Spence, M.P.; Gillaspy, G.; Zhao, B.; Harich, K.; White, R.H.; Li, J. Biochemical evaluation of a parsley tyrosine decarboxylase results in a novel 4-hydroxyphenylacetaldehyde synthase enzyme. Biochem. Biophys. Res. Commun. 2012, 418, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Alagna, F.; Mariotti, R.; Panara, F.; Caporali, S.; Urbani, S.; Veneziani, G.; Esposto, S.; Taticchi, A.; Rosati, A.; Rao, R.; et al. Olive phenolic compounds: Metabolic and transcriptional profiling during fruit development. BMC Plant. Biol. 2012, 12, 162. [Google Scholar] [CrossRef] [PubMed]
- Gutensohn, M.; Klempien, A.; Kaminaga, Y.; Nagegowda, D.A.; Negre-Zakharov, F.; Huh, J.H.; Luo, H.; Weizbauer, R.; Mengiste, T.; Tholl, D.; et al. Role of aromatic aldehyde synthase in wounding/herbivory response and flower scent production in different Arabidopsis ecotypes. Plant. J. 2011, 66, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Velázquez-Palmero, D.; Romero-Segura, C.; García-Rodríguez, R.; Hernández, M.L.; Vaistij, F.E.; Graham, I.A.; Pérez;A.G.;Martínez-Rivas, J.M. An oleuropein β-glucosidase from olive fruit is involved in determining the phenolic composition of virgin olive oil. Front. Plant. Sci. 2017, 8, 1902. [Google Scholar] [CrossRef]
- Gómez-Rico, A.; Salvador, M.D.; La Greca, M.; Fregapane, G. Phenolic and volatile compounds of extra virgin olive oil (Olea europaea L. Cv. Cornicabra) with regard to fruit ripening and irrigation management. J. Agric. Food Chem. 2006, 54, 7130–7136. [Google Scholar] [CrossRef] [PubMed]
- Hbaieb, H.; Kotti, F.; García-Rodríguez, R.; Gargouri, M.; Sanz, C.; Pérez, A.G. Monitoring endogenous enzymes during olive fruit ripening and storage: Correlation with virgin olive oil phenolic profiles. Food Chem. 2015, 173, 972–978. [Google Scholar] [CrossRef]
- Cirilli, M.; Caruso, G.; Gennai, C.; Urbani, S.; Frioni, E.; Ruzzi, M.; Servili, M.; Gucci, R.; Poerio, E.; Muleo, R.M. The Role of Polyphenoloxidase, Peroxidase, and -Glucosidase in Phenolics Accumulation in Olea europaea L. Fruits under Different Water Regimes. Front. Plant. Sci. 2017, 8, 717. [Google Scholar] [CrossRef] [PubMed]
- Mougiou, N.; Trikkab, F.; Trantasc, E.; Ververidisc, F.; Makrisb, A.; Argirioub, A.; Vlachonasiosa, K.E. Expression of hydroxytyrosol and oleuropein biosynthetic genes are correlated with metabolite accumulation during fruit development in olive, Olea europaea, cv. Koroneiki. Plant. Physiol. Bioch 2018, 128, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Alagna, F.; Geu-Flores, F.; Kries, H.; Panara, F.; Baldoni, L.; O’Connor, S.; Osbourn, A. Identification and Characterization of the Iridoid Synthase Involved in Oleuropein Biosynthesis in Olive (Olea europaea) Fruits. J. Biol. Chem. 2016. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PICUAL Phenolics | ||||
|---|---|---|---|---|
| (µg/g FW) | Stage I | Stage II | Stage III | Stage IV |
| Hydroxytyrosol-G | 480.6 ± 9.7* | 480.7 ± 39.4 | 283.8 ± 4.0 | 975.8 ± 30.6 |
| Tyrosol-G | 68.5 ± 2.4 | 68.8 ± 1.8 | 45.0 ± 0.5 | 95.5 ± 1.6 |
| Demethyloleuropein | 242.7 ± 38.9 | 82.6 ± 19.5 | 142.9 ± 0.5 | 16.4 ± 1.8 |
| Demethyligstroside | 159.4 ± 1.2 | 36.9 ± 16.4 | 35.6 ± 2.2 | 22.3 ± 2.6 |
| Oleuropein | 30,838.0± 1472.1 | 27,683.1 ± 1004.0 | 18,793.2 ± 67.3 | 7791.4 ± 84.4 |
| Ligstroside | 2513.0 ± 242.9 | 2210.9 ± 144.0 | 1328.1 ± 19.4 | 755.0 ± 4.4 |
| Verbascoside | 1412.1 ± 253.5 | 1944.7 ± 173.2 | 1256 ± 67.6 | 1816.2 ± 30.6 |
| Luteolin-7-G | 677.8 ± 113.5 | 494.1 ± 69.4 | 452.1 ± 11.0 | 180.7 ± 13.9 |
| total phenolics | 36,154.3 ± 1014.2 | 33,735.6 ± 1037.8 | 22,422.7 ± 122.0 | 11,738.2 ± 120.1 |
| MENYA Phenolics | ||||
| (µg/g FW) | Stage I | Stage II | Stage III | Stage IV |
| Hydroxytyrosol-G | 434.8 ± 27.9 | 570.4 ± 168.7 | 463.8 ± 28.9 | 444 ± 13.7 |
| Tyrosol-G | 60.3 ± 6.8 | 91.1 ± 27.9 | 52.1 ± 0.8 | 99.5 ± 0.1 |
| Demethyloleuropein | 269 ± 1.6 | 125.9 ± 20.3 | 1469.4 ± 385.2 | 8357.8 ± 416.3 |
| Demethyligstroside | 37.9 ± 1.1 | 40.4 ± 3.1 | 32.6 ± 6 | 13.5 ± 6.7 |
| Oleuropein | 33,250.5 ± 3011.9 | 29,839 ± 1634.1 | 24,269 ± 930.4 | 2734.2 ± 51.7 |
| Ligstroside | 2793.6 ± 341.1 | 3313.2 ± 76.2 | 3146.3 ± 247.3 | 370.7 ± 11.5 |
| Verbascoside | 112.1 ± 2.1 | 123.9 ± 41.9 | 252.6 ± 44.6 | 326.6 ± 1.7 |
| Luteolin -7-G | 49.9 ± 1.4 | 146.4 ± 23.1 | 122.4 ± 28.3 | 36.6 ± 3.1 |
| total phenolics | 37,120.1 ± 988.6 | 35,247.9 ± 1410.8 | 30,643.9 ± 1181.9 | 12,635.4 ± 357.0 |
| SHENGEH Phenolics | ||||
| (µg/g FW) | Stage I | Stage II | Stage III | Stage IV |
| Hydroxytyrosol-G | 198.6 ± 15.5 | 485.5 ± 2.9 | 685.4 ± 6.4 | 803.3 ± 92.8 |
| Tyrosol-G | 15.1 ± 2.3 | 28.2 ± 0.5 | 84.6 ± 0.8 | 104.3 ± 9.7 |
| Demethyloleuropein | 149 ± 7.7 | 169.7 ± 16.2 | 25 ± 0.8 | 209.7 ± 5.9 |
| Demethyligstroside | 40.6 ± 0.1 | 59.6 ± 2.4 | 44.5 ± 52.7 | 23.2 ± 0.3 |
| Oleuropein | 11,795.3 ± 619.3 | 6617.5 ± 327.7 | 3802.7 ± 309.4 | 4305.1 ± 1 |
| Ligstroside | 1227.5 ± 65.1 | 1005.1 ± 92.1 | 723.8 ± 44.7 | 653.3 ± 0.8 |
| Verbascoside | 11.7 ± 1.8 | 98.7 ± 8.5 | 45.7 ± 16.0 | 120.5 ± 11.9 |
| Luteolin-7-G | 146.4 ± 17.7 | 163 ± 28.6 | 106.9 ± 9.3 | 79.7 ± 8.4 |
| total phenolics | 13,512.2 ± 354.4 | 8866.7 ± 338.6 | 5738.7 ± 311.2 | 6364.6 ± 92.5 |
| PIÑONERA Phenolics | ||||
| (µg/g FW) | Stage I | Stage II | Stage III | Stage IV |
| Hydroxytyrosol-G | 343.4 ± 78.5 | 296.6 ± 77 | 776.1 ± 201.1 | 690.4 ± 24.7 |
| Tyrosol-G | 94.8 ± 49.4 | 62.2 ± 14.9 | 104.1 ± 25.7 | 104.7 ± 0.1 |
| Demethyloleuropein | 222.8 ± 24.4 | 152.9 ± 29.8 | 1706.2 ± 831.5 | 8312.8 ± 321.1 |
| Demethyligstroside | 224.4 ± 97.4 | 15.3 ± 0.1 | 9.5 ± 10.3 | 3.5 ± 2.9 |
| Oleuropein | 25,061.3 ± 2354.7 | 19,747.3 ± 68.3 | 15,778.4 ± 1973.9 | 2537.1 ± 18.7 |
| Ligstroside | 3356.9 ± 1208.3 | 2208.1 ± 10.2 | 1735.7 ± 399.6 | 416.9 ± 55.5 |
| Verbascoside | 1151.4 ± 95.9 | 1308.9 ± 405.1 | 1350.9 ± 72.2 | 1351.5 ± 52.4 |
| Luteolin-7-G | 146.5 ± 34.1 | 90.1 ± 46.0 | 169.5 ± 34.3 | 74.5 ± 10.8 |
| total phenolics | 36,488.2 ± 1208.9 | 24,206.9 ± 460.6 | 23,404.7 ± 2509.3 | 13,734.6 ± 343.8 |
| Cultivar | PICUAL | MENYA | SHENGEH | PIÑONERA | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Phenolics (µg/g) | II | III | IV | II | III | IV | II | III | IV | II | III | IV |
| Hydroxytyrosol | 1.8 ± 0.2* | 1.5 ± 0.3 | 3.2 ± 2.0 | 1.1 ± 0.2 | 1.1 ± 0.1 | 3.4 ± 2.4 | 0.8 ± 0.1 | 1.0 ± 0.2 | 1.5 ± 0.0 | 0.5 ± 0.5 | 1.0 ± 0.1 | 1.6 ± 0.0 |
| Tyrosol | 3.4 ± 1.0 | 1.9 ± 0.1 | 3.6 ± 0.7 | 4.1 ± 0.2 | 3.8 ± 0.1 | 6.2 ± 0.8 | 2.50 ± 0.5 | 3.4 ± 0.7 | 4.1 ± 1.0 | 3.8 ± 0.0 | 3.4 ± 0.1 | 5.5 ± 0.2 |
| Vanillic acid | 0.5 ± 0.1 | 0.2 ± 0.0 | 0.4 ± 0.4 | 0.5 ± 0.0 | 0.4 ± 0.0 | 0.6 ± 0.0 | 0.6 ± 0.1 | 0.7 ± 0.2 | 0.6 ± 0.1 | 1.5 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 |
| Vainillin | 0.2 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.1 | 0.3 ± 0.2 | 0.3 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.3 ± 0.1 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.1 ± 0.0 | 0.3 ± 0.0 |
| p-Coumaric acid | 1.2 ± 0.1 | 0.9 ± 0.1 | 0.4 ± 0.1 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.1 ± 0.0 | 0.6 ± 0.1 | 0.4 ± 0.1 | 0.4 ± 0.1 | 0.6 ± 0.0 | 0.5 ± 0.1 | 0.4 ± 0.0 |
| Hydroxytyrosol ac. | 0.8 ± 0.1 | 0.3 ± 0.0 | 1.6 ± 0.3 | 1.3 ± 0.1 | 2.4 ± 0.6 | 7.5 ± 0.1 | 1.3 ± 0.1 | 2.4 ± 0.8 | 0.3 ± 0.3 | 2.6 ± 0.1 | 3.8 ± 0.0 | 7.8 ± 0.3 |
| 3.4-DHPEA-EDA | 40.9 ± 3.4 | 39.2 ± 1.1 | 19.5 ± 1.4 | 83.69 ± 1.6 | 121.3 ± 13.4 | 256.6 ± 4.7 | 15.6 ± 0.9 | 15.2 ± 1.1 | 6.4 ± 1.5 | 68.9 ± 1.6 | 59.6 ± 4.4 | 98.0 ± 3.6 |
| p-HPEA-EDA | 46.3 ± 0.1 | 36.9 ± 4.6 | 16.7 ± 2.9 | 90.0 ± 10.1 | 87.6 ± 2.7 | 171.5 ± 2.4 | 20.7 ± 1.3 | 20.6 ± 2.7 | 5.1 ± 1.2 | 102.7 ± 6.7 | 60.7 ± 2.0 | 69.4 ± 6.3 |
| Pinoresinol | 12.7 ± 0.5 | 9.3 ± 0.7 | 3.4 ± 0.7 | 7.7 ± 0.2 | 6.2 ± 0.2 | 3.6 ± 0.1 | 2.8 ± 0.3 | 2.9 ± 0.2 | 1.7 ± 0.4 | 2.5 ± 0.1 | 2.1 ± 0.0 | 2.4 ± 0.1 |
| Cinnamic acid | 1.7 ± 0.2 | 1.3 ± 0.3 | 0.3 ± 0.1 | 2.2 ± 0.6 | 2.2 ± 0.2 | 0.9 ± 0.0 | 0.2 ± 0.0 | 0.3 ± 0.0 | 0.1 ± 0.0 | 0.4 ± 0.0 | 0.4 ± 0.0 | 0.6 ± 0.0 |
| Acetoxypinoresinol | 37.1 ± 0.3 | 32.1 ± 0.7 | 14.6 ± 5.9 | 61.4 ± 05 | 47.7 ± 0.5 | 22.1 ± 2.1 | 9.8 ± 0.7 | 9.6 ± 0.7 | 6.8 ± 1.5 | 4.9 ± 0.0 | 4.4 ± 0.0 | 4.5 ± 0.1 |
| 3.4-DHPEA-EA | 1068.6 ± 15.7 | 854.7 ± 26.4 | 179.7 ± 5.5 | 425.0 ± 41.3 | 388.8 ± 7.3 | 162.9 ± 5.3 | 19.2 ± 3.0 | 37.9 ± 3.4 | 21.6 ± 2.3 | 22.1 ± 1.3 | 11.9 ± 2.0 | 14.5 ± 0.8 |
| p-HPEA-EA | 42.3 ± 1.6 | 37.1 ± 6.6 | 16.8 ± 2.0 | 65.1 ± 3.5 | 40.0 ± 4.3 | 19.2 ± 0.9 | 7.5 ± 0.5 | 9.0 ± 1.1 | 6.6 ± 1.1 | 10.4 ± 0.6 | 5.9 ± 0.0 | 5.6 ± 0.0 |
| Ferulic acid | 0.1 ± 0.0 | 0.0 | 0.2 ± 0.1 | 0.0 | 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 |
| Luteolin | 8.1 ± 1.8 | 8.3 ± 0.1 | 5.8 ± 1.7 | 1.9 ± 0.6 | 1.5 ± 0.0 | 4.4 ± 0.3 | 3.7 ± 0.7 | 4.1 ± 0.2 | 5.0 ± 0.2 | 1.7 ± 0.1 | 2.3 ± 0.0 | 1.9 ± 0.0 |
| Apigenin | 1.9 ± 0.4 | 1.9 ± 0.1 | 1.1 ± 0.4 | 0.4 ± 0.1 | 0.2 ± 0.0 | 0.6 ± 0.0 | 0.7 ± 0.2 | 0.6 ± 0.1 | 0.6 ± 0.0 | 0.2 ± 0.0 | 0.3 ± 0.0 | 0.2 ± 0.0 |
| total phenolics | 1267.7 ± 20.0 | 1025.8 ± 20.9 | 267.5 ± 3.0 | 744.8 ± 69.0 | 703.8 ± 19.6 | 659.8 ± 0.7 | 86.4 ± 2.1 | 108.6 ± 4.8 | 61.1 ± 8.8 | 223.3 ± 10.8 | 156.6 ± 4.4 | 212.9 ± 11.2 |
| total o-phe | 1120.3 ± 17.5 | 904.1 ± 27.9 | 209.9 ± 4.4 | 513.0 ± 55.7 | 515.1 ± 8.8 | 434.8 ± 3.4 | 40.6 ± 3.1 | 60.5 ± 1.0 | 34.8 ± 3.3 | 95.8 ± 3.4 | 78.6 ± 2.4 | 123.8 ± 4.6 |
| secoiridoids | 1198.2 ± 20.6 | 967.9 ± 25.8 | 232.8 ± 3.0 | 663.8 ± 68.3 | 637.7 ± 15.4 | 610.3 ± 0.9 | 63.0 ± 4.6 | 82.7 ± 1.5 | 39.8 ± 6.0 | 204.1 ± 10.1 | 138.1 ± 4.6 | 187.5 ± 10.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez, R.; García-Vico, L.; Sanz, C.; Pérez, A.G. An Aromatic Aldehyde Synthase Controls the Synthesis of Hydroxytyrosol Derivatives Present in Virgin Olive Oil. Antioxidants 2019, 8, 352. https://doi.org/10.3390/antiox8090352
Sánchez R, García-Vico L, Sanz C, Pérez AG. An Aromatic Aldehyde Synthase Controls the Synthesis of Hydroxytyrosol Derivatives Present in Virgin Olive Oil. Antioxidants. 2019; 8(9):352. https://doi.org/10.3390/antiox8090352
Chicago/Turabian StyleSánchez, Rosario, Lourdes García-Vico, Carlos Sanz, and Ana G. Pérez. 2019. "An Aromatic Aldehyde Synthase Controls the Synthesis of Hydroxytyrosol Derivatives Present in Virgin Olive Oil" Antioxidants 8, no. 9: 352. https://doi.org/10.3390/antiox8090352
APA StyleSánchez, R., García-Vico, L., Sanz, C., & Pérez, A. G. (2019). An Aromatic Aldehyde Synthase Controls the Synthesis of Hydroxytyrosol Derivatives Present in Virgin Olive Oil. Antioxidants, 8(9), 352. https://doi.org/10.3390/antiox8090352