Polyamine Induction in Postharvest Banana Fruits in Response to NO Donor SNP Occurs via l-Arginine Mediated Pathway and Not via Competitive Diversion of S-Adenosyl-l-Methionine

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples, Treatments, Nitrite and NO Measurements
2.2. Quality and Shelf Life
2.3. Ethylene Measurement by Gas Chromatography
2.4. Extraction and Estimation of Polyamines
2.5. RNA Extraction, DNase Treatment and RNA Quantification
2.6. First Strand cDNA Synthesis
2.7. Primer Designing
2.8. Semi-Quantitative RT-PCR
2.9. Quantitative Real Time PCR
2.10. Statistical Analysis
3. Results
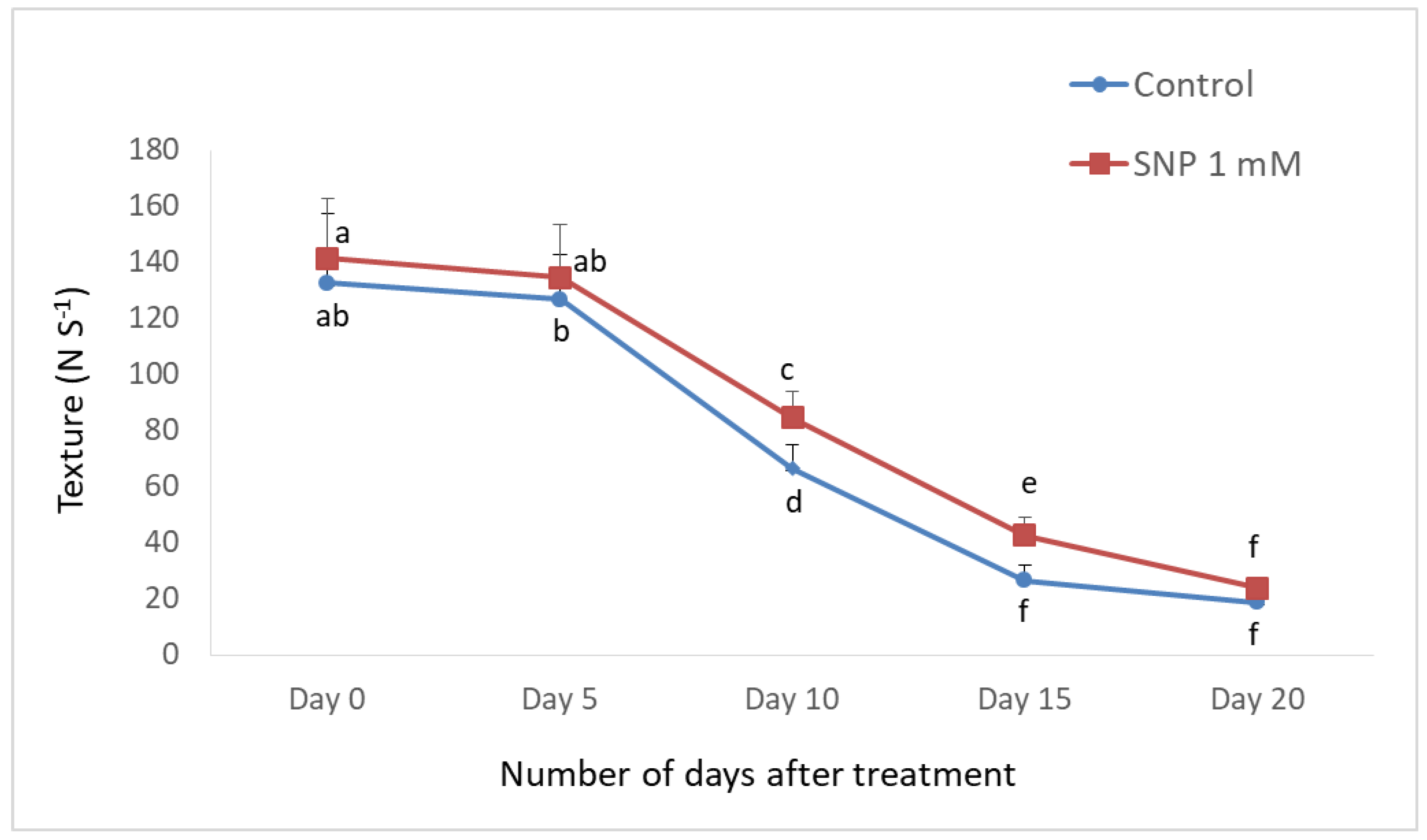
3.1. NO Treatment Differentially Affects the Shelf Life
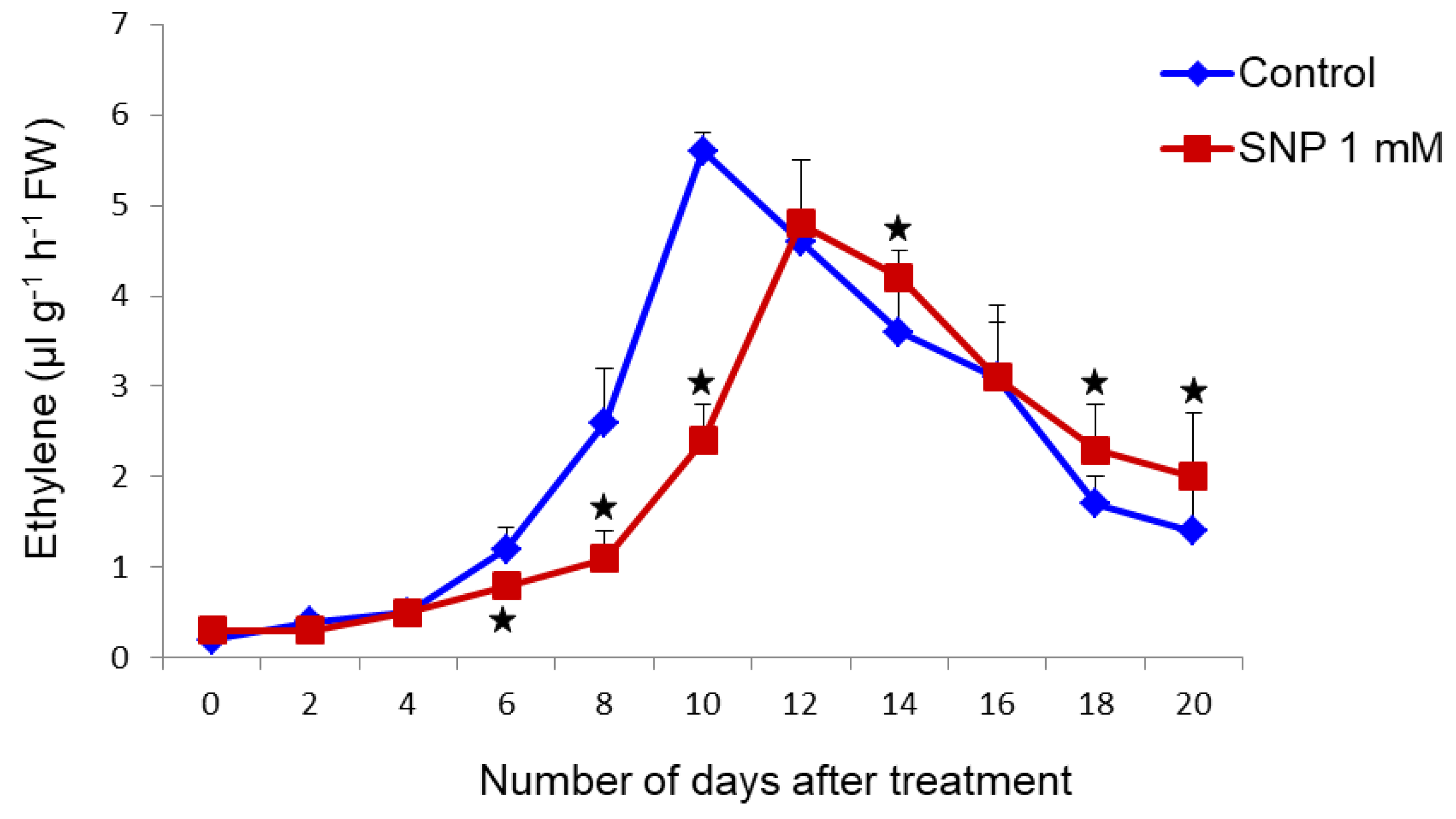
3.2. SNP Treatment Leads to Reduced Ethylene Production
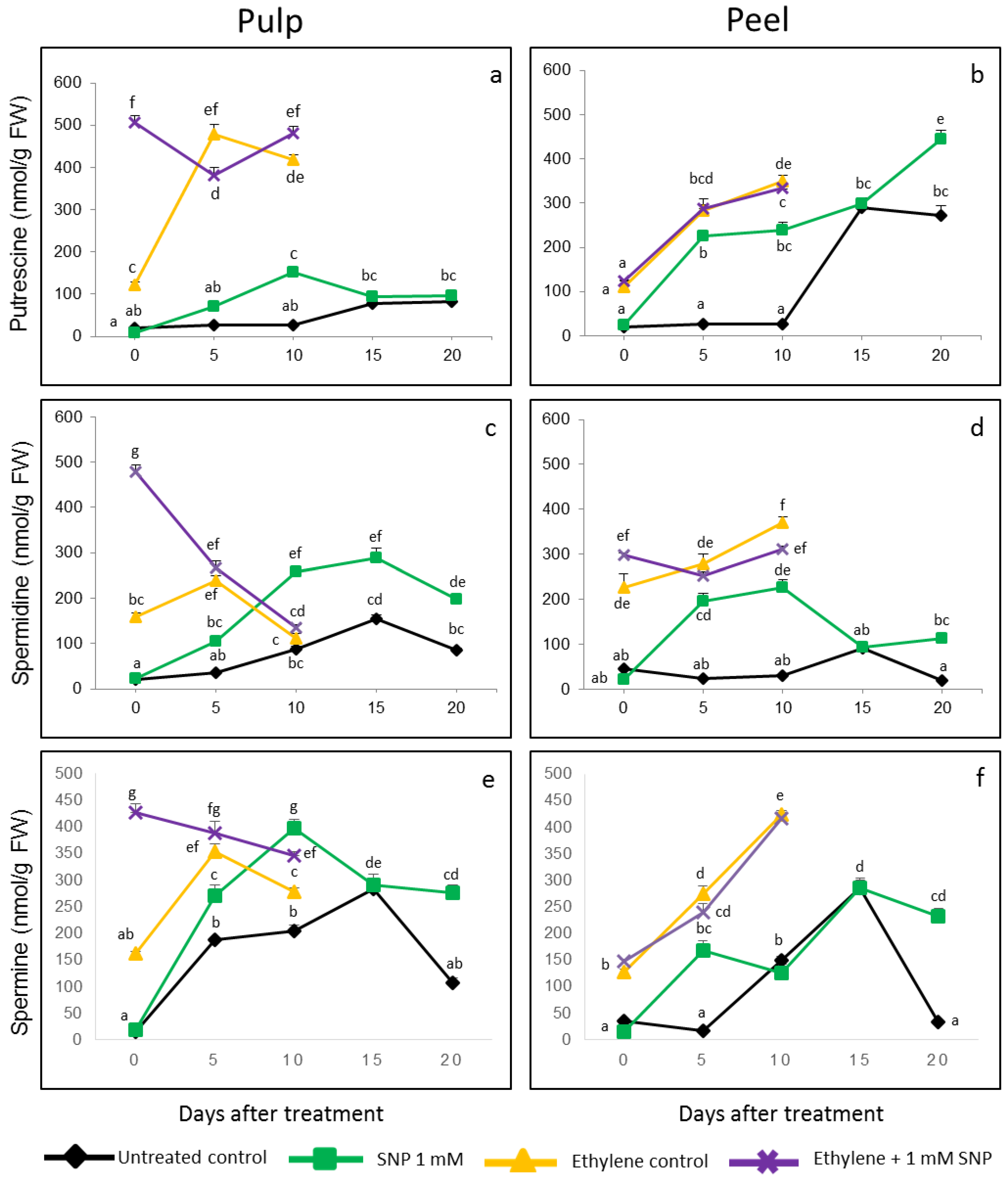
3.3. Acceleration of Putrescine (Put), Spermidine (Spd) and Spermine (Spm) Production in Response to SNP Treatment
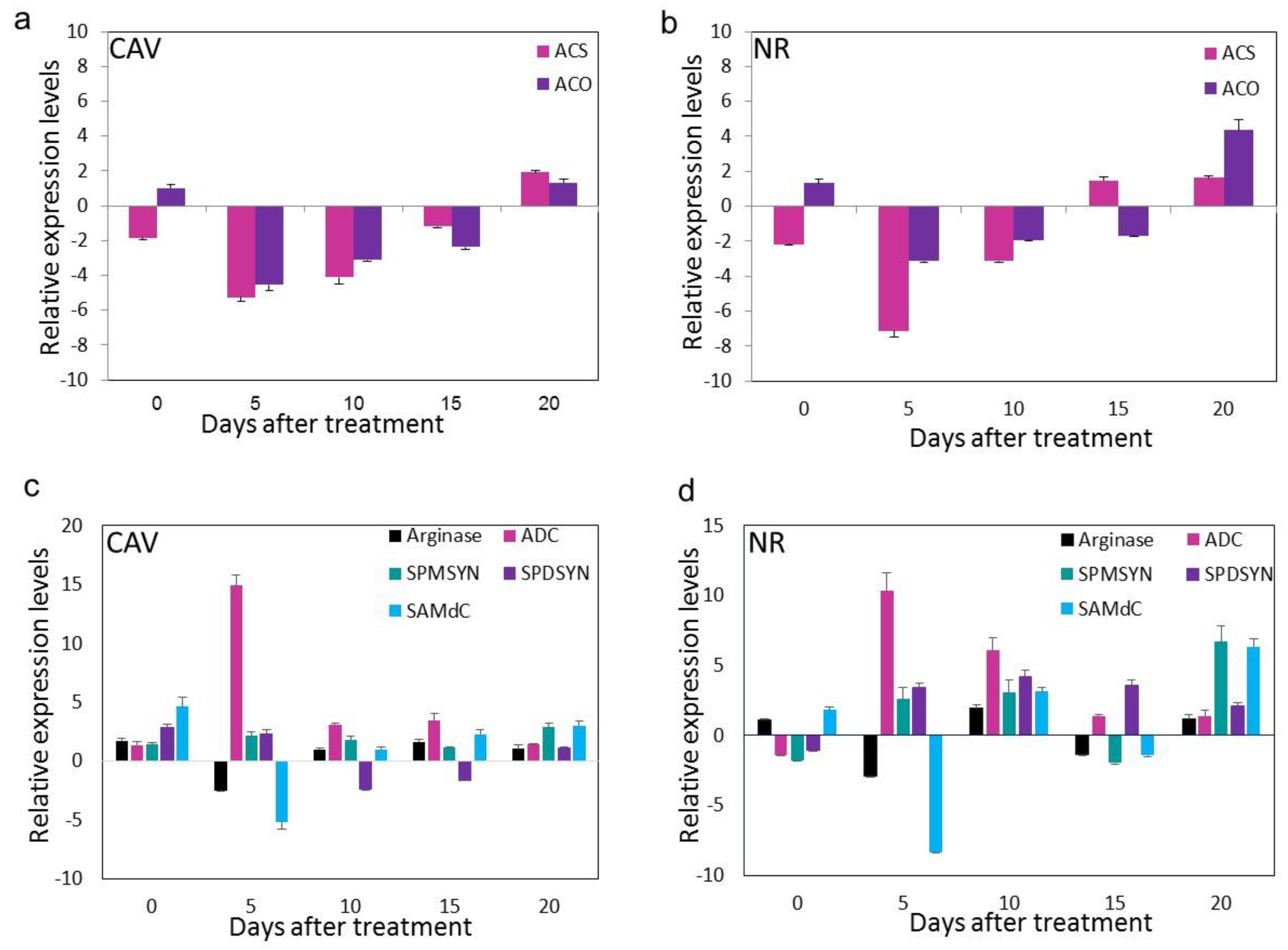
3.4. SNP Down-Regulates Genes Involved in Ethylene Biosynthesis
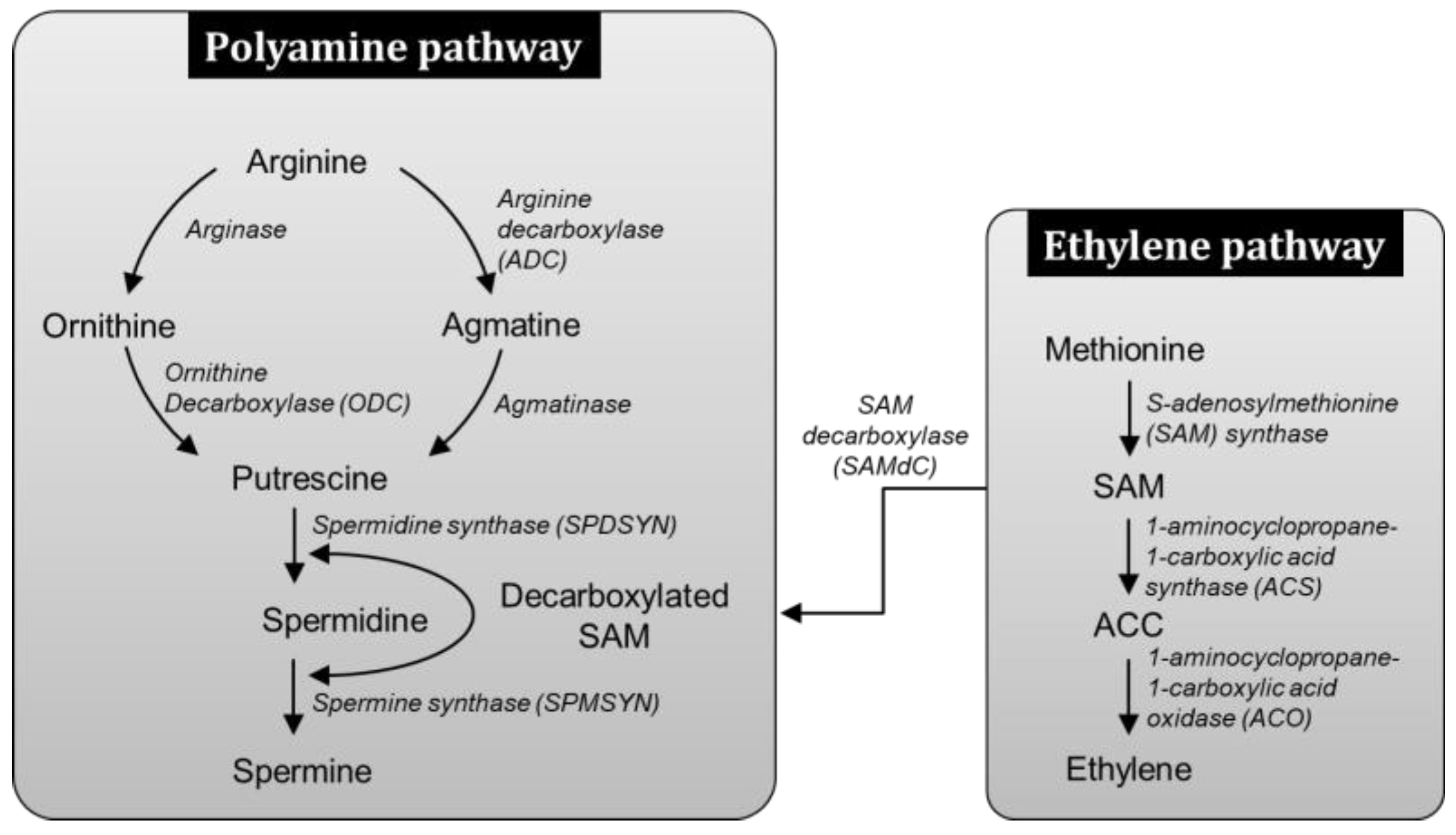
3.5. SNP Increases PA Biosynthetic Genes via Arg-Mediated Route and Not by Precursor SAM Pool Diversion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Manjunatha, G.; Lokesh, V.; Neelwarne, B. Nitric oxide in fruit ripening: Trends and opportunities. Biotechnol. Adv. 2010, 28, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Leshem, Y.A.Y. Nitric oxide in biological systems. Plant Growth Regul. 1996, 18, 155–159. [Google Scholar] [CrossRef]
- Leshem, Y.Y.; Wills, R.B.; Ku, V.V.V. Evidence for the function of the free radical gas—Nitric Oxide (NO•)—As an endogenous maturation and senescence regulating factor in higher plants. Plant Physiol. Biochem. 1998, 36, 825–833. [Google Scholar] [CrossRef]
- Leshem, Y.A.Y.; Pinchasov, Y. Non-invasive photoacoustic spectroscopic determination of relative endogenous nitric oxide and ethylene content stoichiometry during the ripening of strawberries Fragaria anannasa (Duch.) and avocados Persea americana (Mill.). J. Exp. Bot. 2000, 51, 1471–1473. [Google Scholar] [PubMed]
- Cheng, G.; Yang, E.; Lu, W.; Jia, Y.; Jiang, Y.; Duan, X. Effect of nitric oxide on ethylene synthesis and softening of banana fruit slice during ripening. J. Agric. Food Chem. 2009, 57, 5799–5804. [Google Scholar] [CrossRef]
- Manjunatha, G.; Lokesh, V.; Bhagyalakshmi, N. Nitric oxide-induced enhancement of banana fruit attributes and keeping quality. Acta Hort. 2010, 934, 799–806. [Google Scholar] [CrossRef]
- Chaki, M.; Álvarez de Morales, P.; Ruiz, C.; Begara-Morales, J.C.; Barroso, J.B.; Corpas, F.J.; Palma, J.M. Ripening of pepper (Capsicum annuum) fruit is characterized by an enhancement of protein tyrosine nitration. Ann. Bot. 2015, 116, 637–647. [Google Scholar] [CrossRef]
- Rodríguez-Ruiz, M.; Mateos, R.M.; Codesido, V.; Corpas, F.J.; Palma, J.M. Characterization of the galactono-1, 4-lactone dehydrogenase from pepper fruits and its modulation in the ascorbate biosynthesis. Role of nitric oxide. Redox Biol. 2017, 12, 171–181. [Google Scholar] [CrossRef]
- González-Gordo, S.; Bautista, R.; Claros, M.G.; Cañas, A.; Palma, J.M.; Corpas, F.J. Nitric oxide-dependent regulation of sweet pepper fruit ripening. J. Exp. Bot. 2019. [Google Scholar] [CrossRef]
- Duan, X.; Su, X.; You, Y.; Qu, H.; Li, Y.; Jiang, Y. Effect of nitric oxide on pericarp browning of harvested longan fruit in relation to phenolic metabolism. Food Chem. 2007, 104, 571–576. [Google Scholar] [CrossRef]
- Corpas, F.J.; Palma, J.M. Nitric oxide on/off in fruit ripening. Plant Biol. 2018, 20, 805–807. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; Freschi, L.; Rodríguez-Ruiz, M.; Mioto, P.T.; González-Gordo, S.; Palma, J.M. Nitro-oxidative metabolism during fruit ripening. J. Exp. Bot. 2018, 69, 3449–3463. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.J.; Stoimenova, M.; Kaiser, W.M. In higher plants, only root mitochondria, but not leaf mitochondria reduce nitrite to NO, in vitro and in situ. J. Exp. Bot. 2005, 56, 2601–2609. [Google Scholar] [CrossRef] [PubMed]
- Modolo, L.V.; Augusto, O.; Almeida, I.M.; Magalhaes, J.R.; Salgado, I. Nitrite as the major source of nitric oxide production by Arabidopsis thaliana in response to Pseudomonas syringae. FEBS Lett. 2005, 579, 3814–3820. [Google Scholar] [CrossRef] [PubMed]
- Planchet, E.; Jagadis Gupta, K.; Sonoda, M.; Kaiser, W.M. Nitric oxide emission from tobacco leaves and cell suspensions: Rate limiting factors and evidence for the involvement of mitochondrial electron transport. Plant J. 2005, 41, 732–743. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; Barroso, J.B. Nitric oxide synthase-like activity in higher plants. Nitric Oxide Biol. Chem. 2017, 68, 5. [Google Scholar] [CrossRef] [PubMed]
- Rümer, S.; Gupta, K.J.; Kaiser, W.M. Plant cells oxidize hydroxylamines to NO. J. Exp. Bot. 2009, 60, 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Tun, N.N.; Santa-Catarina, C.; Begum, T.; Silveira, V.; Handro, W.; Floh, E.I.S.; Scherer, G.F. Polyamines induce rapid biosynthesis of nitric oxide (NO) in Arabidopsis thaliana seedlings. Plant Cell Physiol. 2006, 47, 346–354. [Google Scholar] [CrossRef]
- Feuerstein, B.G.; Williams, L.D.; Basu, H.S.; Marton, L.J. Implications and concepts of polyamine-nucleic acid interactions. J. Cell. Biochem. 1991, 46, 37–47. [Google Scholar] [CrossRef]
- Corley, E.; Wolosiuk, R.A.; Hertig, C.M. Regulation of the activation of chloroplast fructose-1, 6-bisphosphatase: Inhibition by spermidine and spermine. BiochemBiophys. Res. Commun. 1983, 115, 707–714. [Google Scholar] [CrossRef]
- Smith, T.A. The inhibition and activation of polyamine oxidase from oat seedlings. In Polyamines in Plants; Springer: Dordrecht, The Netherlands, 1985; pp. 67–73. [Google Scholar]
- Malik, A.U.; Singh, Z. Endogenous free polyamines of mangos in relation to development and ripening. J. Am. Soc. Hortic. Sci. 2004, 129, 280–286. [Google Scholar] [CrossRef]
- Li, N.; Parsons, B.L.; Liu, D.; Mattoo, A.K. Accumulation of wound-inducible ACC synthase transcript in tomato fruit is inhibited by salicylic acid and polyamines. Plant Mol. Biol. 1992, 18, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Mirdehghan, S.H.; Rahemi, M.; Martínez-Romero, D.; Guillén, F.; Valverde, J.M.; Zapata, P.J.; Serrano, M.; Valero, D. Reduction of pomegranate chilling injury during storage after heat treatment: Role of polyamines. Postharvest Biol. Technol. 2007, 44, 19–25. [Google Scholar] [CrossRef]
- Katoh, Y.; Hasegawa, T.; Suzuki, T.; Fujii, T. Effects of 1-aminocyclopropane-1-carboxylic acid production on the changes in the polyamine levels in Hiproly barley callus after auxin withdrawal. Agric. Biol. Chem. 1987, 51, 2457–2463. [Google Scholar]
- Pandey, S.; Ranade, S.A.; Nagar, P.K.; Kumar, N. Role of polyamines and ethylene as modulators of plant senescence. J. Biosci. 2000, 25, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Purwoko, B.S.; Susanto, S.; Novita, T. Studies on the physiology of polyamines and ethylene during ripening of banana and papaya fruits. Acta Hortic. 2000, 575, 651–657. [Google Scholar] [CrossRef]
- Bouchereau, A.; Aziz, A.; Larher, F.; Martin-Tanguy, J. Polyamines and environmental challenges: Recent development. Plant Sci. 1999, 140, 103–125. [Google Scholar] [CrossRef]
- Keefer, L.K.; Nims, R.W.; Davies, K.M.; Wink, D.A. “NONOates”(1-substituted diazen-1-ium-1, 2-diolates) as nitric oxide donors: Convenient nitric oxide dosage forms. Methods Enzymol. 1996, 268, 281–293. [Google Scholar] [PubMed]
- Kende, H. Ethylene biosynthesis. Annu. Rev. Plant Biol. 1993, 44, 283–307. [Google Scholar] [CrossRef]
- Walden, R.; Cordeiro, A.; Tiburcio, A.F. Polyamines: Small molecules triggering pathways in plant growth and development. Plant Physiol. 1997, 113, 1009. [Google Scholar] [CrossRef]
- Wang, D.; Li, L.; Xu, Y.; Limwachiranon, J.; Li, D.; Ban, Z.; Luo, Z. Effect of exogenous nitro oxide on chilling tolerance, polyamine, proline, and γ-aminobutyric acid in Bamboo Shoots (Phyllostachys praecox f. prevernalis). J. Agric. Food Chem. 2017, 65, 5607–5613. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.F.; Du, C.X.; Guo, S.R. Nitric oxide enhances salt tolerance in cucumber seedlings by regulating free polyamine content. Environ. Exper. Bot. 2013, 86, 52–59. [Google Scholar] [CrossRef]
- McGuire, R.G. Reporting of objective color measurements. HortScience 1992, 27, 1254–1255. [Google Scholar] [CrossRef]
- Breene, W.M. Application of texture profile analysis to instrumental food texture evaluation. J. Texture Stud. 1975, 6, 53–82. [Google Scholar] [CrossRef]
- Pathak, N.; Asif, M.H.; Dhawan, P.; Srivastava, M.K.; Nath, P. Expression and activities of ethylene biosynthesis enzymes during ripening of banana fruits and effect of 1-MCP treatment. Plant Growth Regul. 2003, 40, 11–19. [Google Scholar] [CrossRef]
- Flores, H.E.; Galston, A.W. Analysis of polyamines in higher plants by high performance liquid chromatography. Plant Physiol. 1982, 69, 701–706. [Google Scholar] [CrossRef]
- Asif, M.H.; Dhawan, P.; Nath, P. A simple procedure for the isolation of high quality RNA from ripening banana fruit. Plant Mol. Biol. Rep. 2000, 18, 109–115. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, Y.; Ge, Y.; Zhao, J.; Wei, M.; Li, C.; Hou, J.; Cheng, Y.; Chen, J. Postharvest sodium nitroprusside treatment maintains storage quality of apple fruit by regulating sucrose metabolism. Postharvest Biol. Technol. 2019, 154, 115–120. [Google Scholar] [CrossRef]
- Ge, Y.; Li, X.; Li, C.; Tang, Q.; Duan, B.; Cheng, Y.; Hou, J.; Li, J. Effect of sodium nitroprusside on antioxidative enzymes and the phenylpropanoid pathway in blueberry fruit. Food Chem. 2019, 295, 607–612. [Google Scholar] [CrossRef]
- Han, S.E.; Seo, Y.S.; Kim, D.; Sung, S.K.; Kim, W.T. Expression of MdCAS1 and MdCAS2, encoding apple β-cyanoalanine synthase homologs, is concomitantly induced during ripening and implicates MdCASs in the possible role of the cyanide detoxification in Fuji apple (Malus domestica Borkh.) fruits. Plant Cell Rep. 2007, 26, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Flores, F.B.; Sánchez-Bel, P.; Valdenegro, M.; Romojaro, F.; Martínez-Madrid, M.C.; Egea, M.I. Effects of a pretreatment with nitric oxide on peach (Prunuspersica L.) storage at room temperature. Eur. Food Res. Technol. 2008, 227, 1599. [Google Scholar] [CrossRef]
- Li, X.P.; Wu, B.; Guo, Q.; Wang, J.D.; Zhang, P.; Chen, W.X. Effects of nitric oxide on postharvest quality and soluble sugar content in papaya fruit during ripening. J. Food Process Preserv. 2014, 38, 591–599. [Google Scholar] [CrossRef]
- Adão, R.C.; Glória, M.B.A. Bioactive amines and carbohydrate changes during ripening of ‘Prata’banana (Musa acuminata× M. balbisiana). Food Chem. 2005, 90, 705–711. [Google Scholar] [CrossRef]
- Apelbaum, A.; Burgoon, A.C.; Anderson, J.D.; Lieberman, M.; Ben-Arie, R.; Mattoo, A.K. Polyamines inhibit biosynthesis of ethylene in higher plant tissue and fruit protoplast. Plant Physiol. 1981, 68, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Even-Chen, Z.; Mattoo, A.K.; Goren, R. Inhibition of ethylene biosynthesis by aminoethoxyvinylglycine and by polyamines shunts label from 3,4-[14C-methionine] into spermidine in aged orange peel discs. Plant Physiol. 1982, 69, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Park, K.Y.; Lee, S.H. Effects of ethylene and auxin on polyamine levels in suspension-cultured tobacco cells. Physiol. Plant 1994, 90, 382–390. [Google Scholar] [CrossRef]
- Mehta, R.A.; Cassol, T.; Li, N.; Ali, N.; Handa, A.K.; Mattoo, A.K. Engineered polyamine accumulation in tomato enhances phytonutrient content, juice quality, and vine life. Nat. Biotechnol. 2002, 20, 613. [Google Scholar] [CrossRef]
- Bregoli, A.M.; Scaramagli, S.; Costa, G.; Sabatini, E.; Ziosi, V.; Biondi, S.; Torrigiani, P. Peach (Prunuspersica) fruit ripening: Aminoethoxyvinylglycine (AVG) and exogenous polyamines affect ethylene emission and flesh firmness. Physiol. Plant. 2002, 114, 472–481. [Google Scholar] [CrossRef]
- Aziz, A.; Martin-Tanguy, J.; Larher, F. Plasticity of polyamine metabolism associated with high osmotic stress in rape leaf discs and with ethylene treatment. Plant Growth Regul. 1997, 21, 153–163. [Google Scholar] [CrossRef]
- Filippou, P.; Antoniou, C.; Fotopoulos, V. The nitric oxide donor sodium nitroprusside regulates polyamine and proline metabolism in leaves of Medicagotruncatula plants. Free Radic. Biol. Med. 2013, 56, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Tassoni, A.; Watkins, C.B.; Davies, P.J. Inhibition of the ethylene response by 1-MCP in tomato suggests that polyamines are not involved in delaying ripening, but may moderate the rate of ripening or over-ripening. J. Exp. Bot. 2006, 57, 3313–3325. [Google Scholar] [CrossRef] [PubMed]
- Van de Poel, B.; Bulens, I.; Oppermann, Y.; Hertog, M.L.; Nicolai, B.M.; Sauter, M.; Geeraerd, A.H. S-adenosyl-l-methionine usage during climacteric ripening of tomato in relation to ethylene and polyamine biosynthesis and transmethylation capacity. Physiol. Plant. 2013, 148, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Kushad, M.M.; Yelenosky, G.; Knight, R. Interrelationship of polyamine and ethylene biosynthesis during avocado fruit development and ripening. Plant Physiol. 1988, 87, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Paschalidis, K.A.; Moschou, P.N.; Aziz, A.; Toumi, I.; Roubelakis-Angelakis, K.A. Polyamines in grapevine: An update. In Grapevine Molecular Physiology & Biotechnology; Springer: Dordrecht, The Netherlands, 2009; pp. 207–228. [Google Scholar]
- Agudelo-Romero, P.; Bortolloti, C.; Pais, M.S.; Tiburcio, A.F.; Fortes, A.M. Study of polyamines during grape ripening indicate an important role of polyamine catabolism. Plant Physiol. Biochem. 2013, 67, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Fortes, A.M.; Teixeira, R.T.; Agudelo-Romero, P. Complex interplay of hormonal signals during grape berry ripening. Molecules 2015, 20, 9326–9343. [Google Scholar] [CrossRef]
- Tsaniklidis, G.; Kotsiras, A.; Tsafouros, A.; Roussos, P.A.; Aivalakis, G.; Katinakis, P.; Delis, C. Spatial and temporal distribution of genes involved in polyamine metabolism during tomato fruit development. Plant Physiol. Biochem. 2016, 100, 27–36. [Google Scholar] [CrossRef]
- Hanfrey, C.; Sommer, S.; Mayer, M.J.; Burtin, D.; Michael, A.J. Arabidopsis polyamine biosynthesis: Absence of ornithine decarboxylase and the mechanism of arginine decarboxylase activity. Plant J. 2001, 27, 551–560. [Google Scholar] [CrossRef]
- D’Hont, A.; Denoeud, F.; Aury, J.M.; Baurens, F.C.; Carreel, F.; Garsmeur, O.; Noel, B.; Bocs, S.; Droc, G.; Rouard, M.; et al. The banana (Musa acuminata) genome and the evolution of monocotyledonous plants. Nature 2012, 488, 213–217. [Google Scholar] [CrossRef]
- Freschi, L. Nitric oxide and phytohormone interactions: Current status and perspectives. Front. Plant Sci. 2013, 4, 398. [Google Scholar] [CrossRef]
- Yoza, K.I.; Takeda, Y.; Sekiya, K.; Nogata, Y.; Ohta, H. Putrescine accumulation in wounded green banana fruit. Phytochemistry 1996, 42, 331–334. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lokesh, V.; Manjunatha, G.; Hegde, N.S.; Bulle, M.; Puthusseri, B.; Gupta, K.J.; Neelwarne, B. Polyamine Induction in Postharvest Banana Fruits in Response to NO Donor SNP Occurs via l-Arginine Mediated Pathway and Not via Competitive Diversion of S-Adenosyl-l-Methionine. Antioxidants 2019, 8, 358. https://doi.org/10.3390/antiox8090358
Lokesh V, Manjunatha G, Hegde NS, Bulle M, Puthusseri B, Gupta KJ, Neelwarne B. Polyamine Induction in Postharvest Banana Fruits in Response to NO Donor SNP Occurs via l-Arginine Mediated Pathway and Not via Competitive Diversion of S-Adenosyl-l-Methionine. Antioxidants. 2019; 8(9):358. https://doi.org/10.3390/antiox8090358
Chicago/Turabian StyleLokesh, Veeresh, Girigowda Manjunatha, Namratha S. Hegde, Mallesham Bulle, Bijesh Puthusseri, Kapuganti Jagadis Gupta, and Bhagyalakshmi Neelwarne. 2019. "Polyamine Induction in Postharvest Banana Fruits in Response to NO Donor SNP Occurs via l-Arginine Mediated Pathway and Not via Competitive Diversion of S-Adenosyl-l-Methionine" Antioxidants 8, no. 9: 358. https://doi.org/10.3390/antiox8090358
APA StyleLokesh, V., Manjunatha, G., Hegde, N. S., Bulle, M., Puthusseri, B., Gupta, K. J., & Neelwarne, B. (2019). Polyamine Induction in Postharvest Banana Fruits in Response to NO Donor SNP Occurs via l-Arginine Mediated Pathway and Not via Competitive Diversion of S-Adenosyl-l-Methionine. Antioxidants, 8(9), 358. https://doi.org/10.3390/antiox8090358