Endogenous and Exogenous Melatonin Exposure Attenuates Hepatic MT1 Melatonin Receptor Protein Expression in Rat


 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Drugs and Chemicals
2.2. Animals
2.3. Surgical Procedures
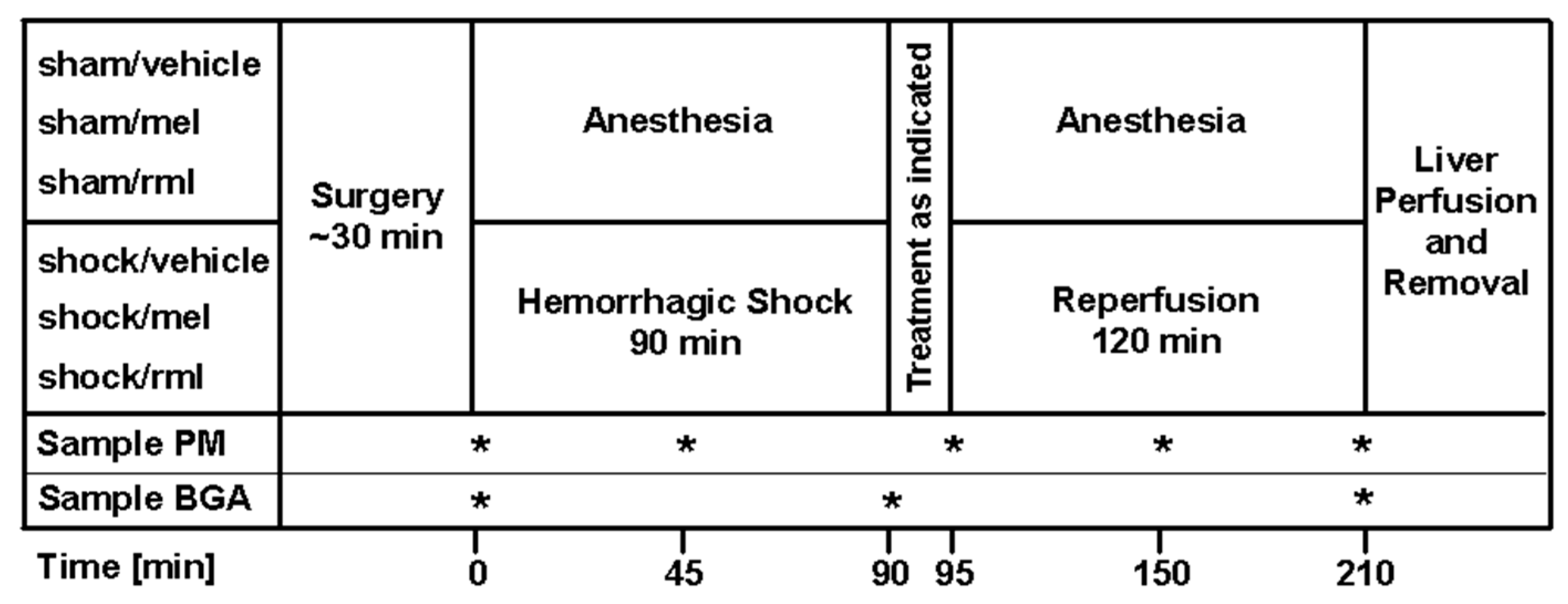
2.4. Experimental Protocol
2.5. Enzyme-Linked Immunosorbent Assay
2.6. RNA Isolation
2.7. RTQ-PCR
2.8. Western Immunoblot Analysis
2.9. Immunohistochemical Staining
2.10. Statistical Analysis
3. Results
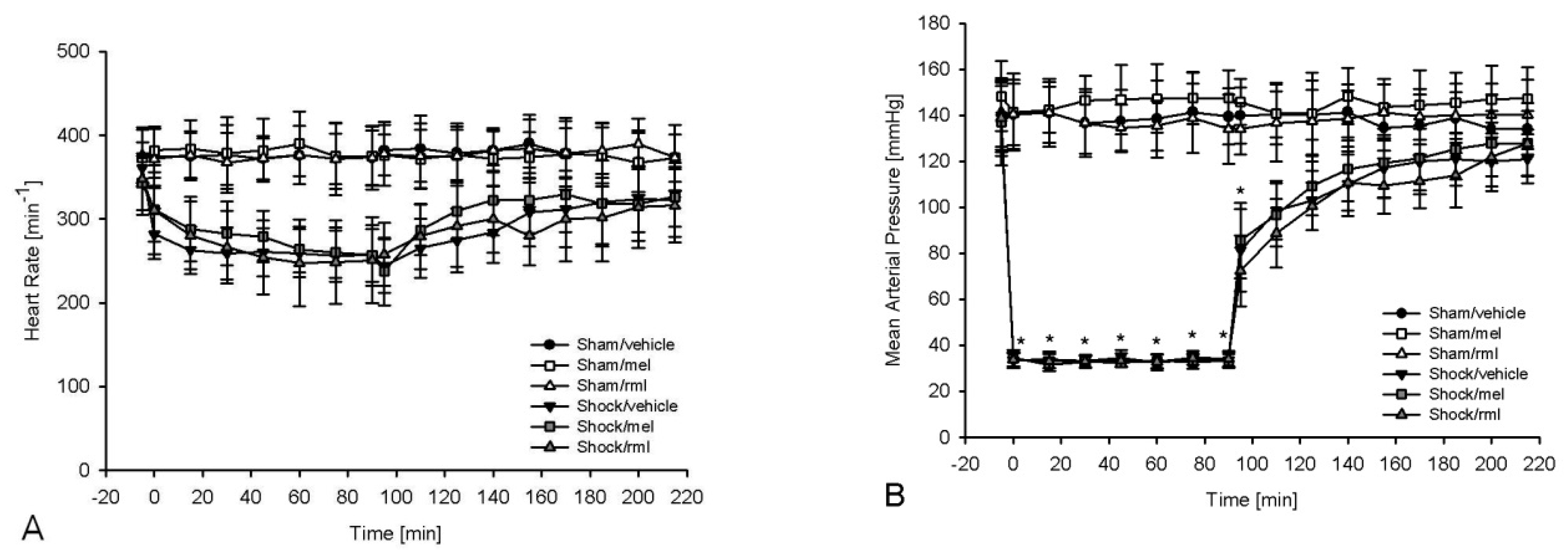
3.1. Hemodynamic Parameters and Analysis of Blood Gases
3.2. Plasma Melatonin Content
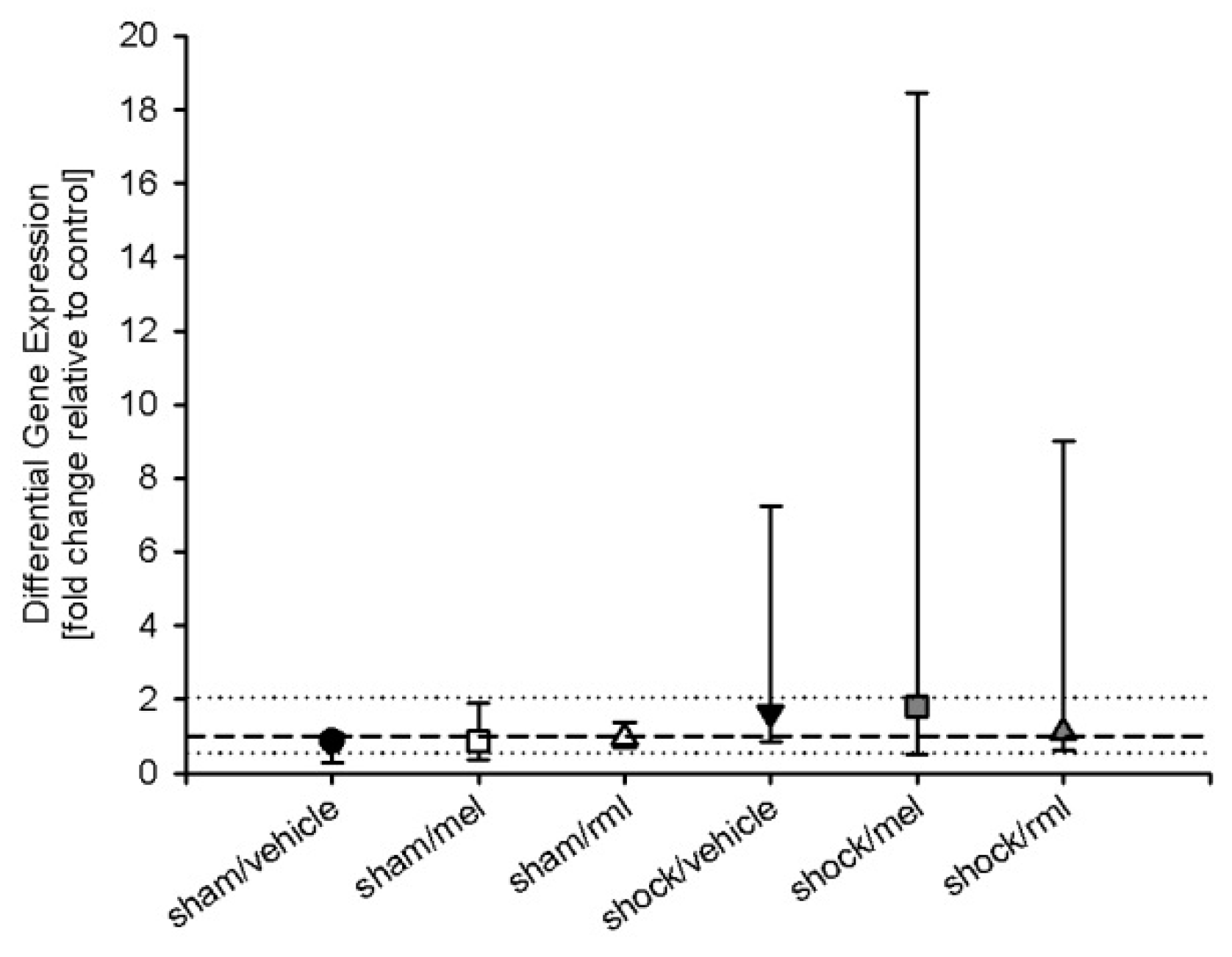
3.3. Melatonin Receptor mRNA
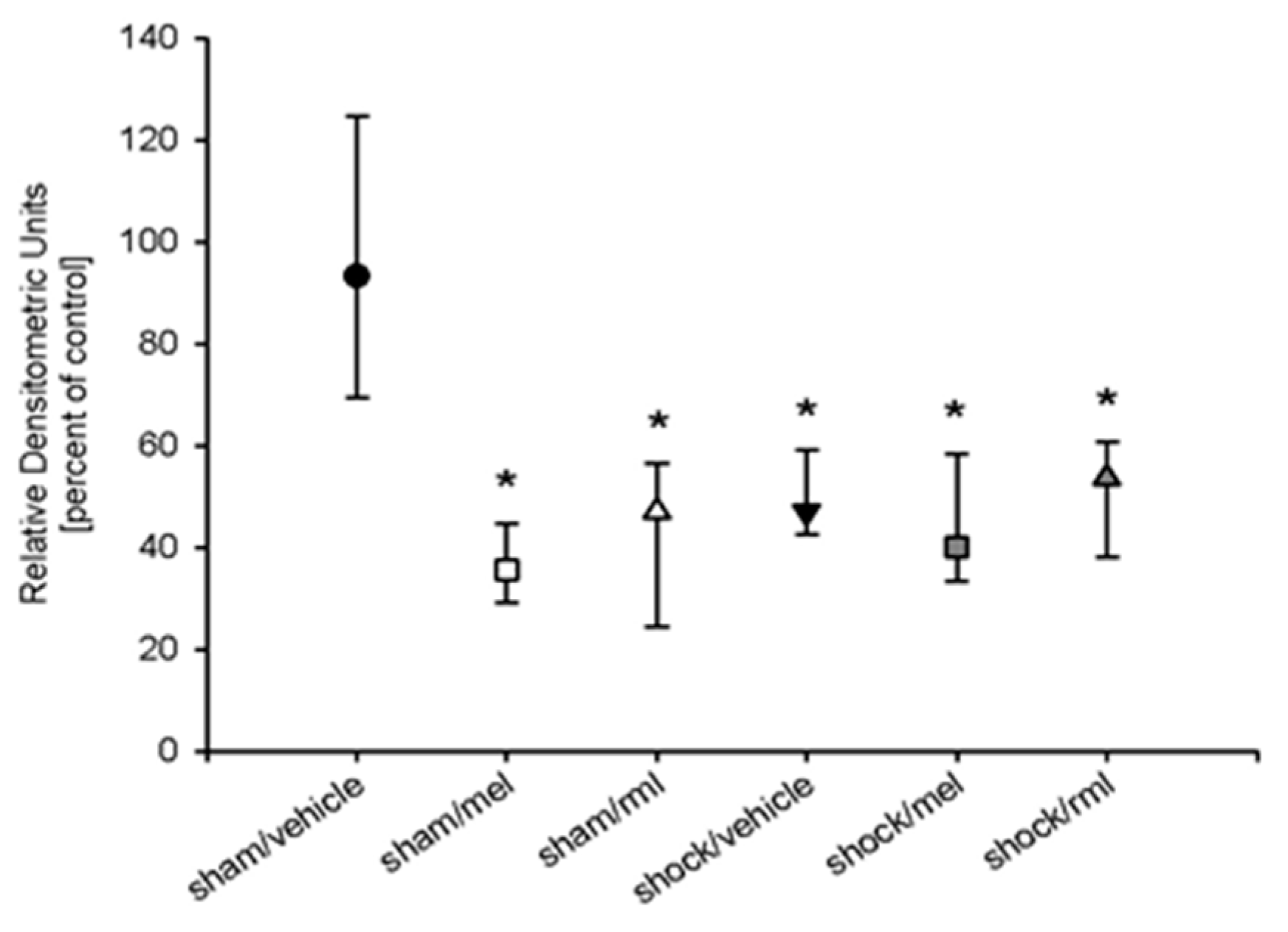
3.4. Melatonin Receptor Protein
3.5. Spatial Expression Pattern of Melatonin Receptors
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mathes, A.M.; Kubulus, D.; Pradarutti, S.; Bentley, A.; Weiler, J.; Wolf, B.; Ziegeler, S.; Bauer, I.; Rensing, H. Melatonin pretreatment improves liver function and hepatic perfusion after hemorrhagic shock. Shock 2008, 29, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Mathes, A.M.; Kubulus, D.; Weiler, J.; Bentley, A.; Waibel, L.; Wolf, B.; Bauer, I.; Rensing, H. Melatonin receptors mediate improvements of liver function but not of hepatic perfusion and integrity after hemorrhagic shock in rat. Crit. Care Med. 2008, 36, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Le, P.H.; Kuo, C.J.; Cheng, H.T.; Wu, R.C.; Chen, T.H.; Lin, C.J.; Chiang, K.C.; Hsu, J.T. Melatonin Attenuates Acute Pancreatitis-Induced Liver Damage Through Akt-Dependent PPAR-γ Pathway. J. Surg. Res. 2018, 236, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Sewerynek, E.; Reiter, R.J.; Melchiorri, D.; Ortiz, G.G.; Lewinski, A. Oxidative damage in the liver induced by ischemia-reperfusion: Protection by melatonin. Hepatogastroenterology 1996, 43, 898–905. [Google Scholar] [PubMed]
- Kurcer, Z.; Oğuz, E.; Iraz, M.; Fadillioglu, E.; Baba, F.; Koksal, M.; Olmez, E. Melatonin improves methanol intoxication-induced oxidative liver injury in rats. J. Pineal Res. 2007, 43, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Fang, Y.; Lu, Y.; Tan, D.; Du, C.; Li, Y.; Ma, Q.; Yu, J.; Chen, M.; Zhou, C.; et al. Melatonin alleviates cadmium-induced liver injury by inhibiting the TXNIP-NLRP3 inflammasome. J. Pineal Res. 2017, 62, e12389. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Kongo, M.; Sasaki, E.; Nishida, K.; Ishiguro, I. Therapeutic effect of melatonin on carbon tetrachloride-induced acute liver injury in rats. J. Pineal Res. 2000, 28, 119–126. [Google Scholar] [CrossRef]
- Sigala, F.; Theocharis, S.; Sigalas, K.; Markantonis-Kyroudis, S.; Papalabros, E.; Triantafyllou, A.; Kostopanagiotou, G.; Andreadou, I. Therapeutic value of melatonin in an experimental model of liver injury and regeneration. J. Pineal Res. 2006, 40, 270–279. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species? J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef]
- Bedini, A.; Fraternale, A.; Crinelli, R.; Mari, M.; Bartolucci, S.; Chiarantini, L.; Spadoni, G. Design, Synthesis, and Biological Activity of Hydrogen Peroxide Responsive Arylboronate Melatonin Hybrids. Chem. Res. Toxicol. 2019, 32, 100–112. [Google Scholar] [CrossRef]
- Lochner, A.; Genade, S.; Davids, A.; Ytrehus, K.; Moolman, J.A. Short- and long-term effects of melatonin on myocardial post-ischemic recovery. J. Pineal Res. 2006, 40, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Rezzani, R.; Rodella, L.F.; Bonomini, F.; Tengattini, S.; Bianchi, R.; Reiter, R.J. Beneficial effects of melatonin in protecting against cyclosporine A-induced cardiotoxicity are receptor mediated. J. Pineal Res. 2006, 41, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Brzozowski, T.; Konturek, P.C.; Zwirska-Korczala, K.; Konturek, S.J.; Brzozowska, I.; Drozdowicz, D.; Sliwowski, Z.; Pawlik, M.; Pawlik, W.W.; Hahn, E.G. Importance of the pineal gland, endogenous prostaglandis and sensory nerves in the gastroprotective actions of central and peripheral melatonin against stress-induced damage. J. Pineal Res. 2005, 39, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Husson, I.; Mesplès, B.; Bac, P.; Vamecq, J.; Evrard, P.; Gressens, P. Melatoninergic neuroprotection of the murine periventricular white matter against neonatal excitotoxic challenge. Ann. Neurol. 2002, 51, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Mathes, A.M.; Kubulus, D.; Waibel, L.; Weiler, J.; Heymann, P.; Wolf, B.; Rensing, H. Selective activation of melatonin receptors with ramelteon improves liver function and hepatic perfusion after hemorrhagic shock in rat. Crit. Care Med. 2008, 36, 2863–2870. [Google Scholar] [CrossRef] [PubMed]
- Sallinen, P.; Saarela, S.; Ilves, M.; Vakkuri, O.; Leppäluoto, J. The expression of MT1 and MT2 melatonin receptor mRNA in several rat tissues. Life Sci. 2005, 76, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Hidalgo, M.; Guerrero Montávez, J.M.; Carrascosa-Salmoral Mdel, P.; Naranjo Gutierrez Mdel, C.; Lardone, P.J.; de la Lastra Romero, C.A. Decreased MT1 and MT2 melatonin receptor expression in extrapineal tissues of the rat during physiological aging. J. Pineal Res. 2009, 46, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Naji, L.; Carrillo-Vico, A.; Guerrero, J.M.; Calvo, J.R. Expression of membrane and nuclear melatonin receptors in mouse peripheral organs. Life Sci. 2004, 74, 2227–2236. [Google Scholar] [CrossRef] [PubMed]
- Kokkola, T.; Vaittinen, M.; Laitinen, J.T. Inverse agonist exposure enhances ligand binding and G protein activation of the human MT1 melatonin receptor, but leads to receptor down-regulation. J. Pineal Res. 2007, 43, 255–262. [Google Scholar] [CrossRef]
- Gerdin, M.J.; Masana, M.I.; Ren, D.; Miller, R.J.; Dubocovich, M.L. Short-term exposure to melatonin differentially affects the functional sensitivity and trafficking of the hMT1 and hMT2 melatonin receptors. J. Pharmacol. Exp. Ther. 2003, 304, 931–939. [Google Scholar] [CrossRef]
- Gerdin, M.J.; Masana, M.I.; Dubocovich, M.L. Melatonin-mediated regulation of human MT (1) melatonin receptors expressed in mammalian cells. Biochem. Pharmacol. 2004, 67, 2023–2030. [Google Scholar] [CrossRef] [PubMed]
- Kleber, A.; Ruf, C.; Wolf, A.; Fink, T.; Glas, M.; Wolf, B.; Volk, T.; Abend, M.; Mathes, A. Melatonin or ramelteon therapy differentially affects hepatic gene expression profiles after haemorrhagic shock in rat—A microarray analysis. Exp. Mol. Pathol. 2015, 99, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, M.W.; Zellweger, R.; DeMaso, C.M.; Ayala, A.; Chaudry, I.H. Increased melatonin levels after hemorrhagic shock in male and female C3H/HeN mice. Experientia 1996, 52, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Cao, J.; Wang, Z.; Dong, Y.; Wang, W.; Chen, Y. Melatonin mediates monochromatic light-induced insulin-like growth factor 1 secretion of chick liver: Involvement of membrane receptors. Photochem. Photobiol. 2016, 92, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Bynum, T.E.; Boitnott, J.K.; Maddrey, W.C. Ischemic hepatitis. Dig. Dis. Sci. 1979, 24, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Mayumi, T.; Chan, C.K.; Clemens, M.G.; Bulkley, G.B. Zonal heterogeneity of hepatic injury following shock/resuscitation: Relationship of xanthine oxidase activity to localisation of neutrophil accumulation and central lobular necrosis. Shock 1996, 5, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Paxian, M.; Rensing, H.; Geckeis, K.; Bauer, I.; Kubulus, D.; Spahn, D.R.; Bauer, M. Perflubron emulsion in prolonged hemorrhagic shock: Influence on hepatocellular energy metabolism and oxygen-dependent gene expression. Anesthesiology 2003, 98, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Rajnik, M.; Salkowski, C.A.; Thomas, K.E.; Li, Y.Y.; Rollwagen, F.M.; Vogel, S.N. Induction of early inflammatory gene expression in a murine model of nonresuscitated, fixed-volume hemorrhage. Shock 2002, 17, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Lagoa, C.E.; Bartels, J.; Baratt, A.; Tseng, G.; Clermont, G.; Fink, M.P.; Billiar, T.R.; Vodovotz, Y. The role of initial trauma in the host’s response to injury and hemorrhage: Insights from a correlation of mathematical simulations and hepatic transcriptomic analysis. Shock 2005, 26, 592–600. [Google Scholar] [CrossRef]
- Tait, S.M.; Wang, P.; Ba, Z.F.; Chaudry, I.H. Downregulation of hepatic beta-adrenergic receptors after trauma and hemorrhagic shock. Am. J. Physiol. 1995, 268, G749–G753. [Google Scholar] [CrossRef]
- Chaste, P.; Clement, N.; Mercati, O.; Guillaume, J.L.; Delorme, R.; Botros, H.G.; Pagan, C.; Périvier, S.; Scheid, I.; Nygren, G.; et al. Identification of Pathway-Biased and Deleterious Melatonin Receptor Mutants in Autism Spectrum Disorders and in the General Population. PLoS ONE 2010, 5, e11495. [Google Scholar] [CrossRef] [PubMed]
- Söderquist, F.; Hellström, P.M.; Cunningham, J.L. Human Gastroenteropancreatic Expression of Melatonin and Its Receptors MT1 and MT2. PLoS ONE 2015, 10, e0120195. [Google Scholar] [CrossRef] [PubMed]
- Tunstall, R.R.; Shukla, P.; Grazul-Bilska, A.; Sun, C.; O’Rourke, S.T. MT2 Receptors Mediate the Inhibitory Effects of Melatonin on Nitric Oxide-Induced Relaxation of Porcine Isolated Coronary Arteries. J. Pharmacol. Exp. Ther. 2011, 336, 127–133. [Google Scholar] [CrossRef]
- Sethi, S.; Adams, W.; Pollock, J.; Witt-Enderby, P.A. C-terminal domains within human MT1 and MT2 melatonin receptors are involved in internalization processes. J. Pineal Res. 2008, 45, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Mühlbauer, E.; Gross, E.; Labucay, K.; Wolgast, S.; Peschke, E. Loss of melatonin signalling and its impact on circadian rhythms in mouse organs regulating blood glucose. Eur. J. Pharmacol. 2009, 606, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Poon, A.M.; Choy, E.H.; Pang, S.F. Modulation of blood glucose by melatonin: A direct action on melatonin receptors in mouse hepatocytes. Biol. Signals Recept. 2001, 10, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Chao, W.W.; Chen, S.J.; Peng, H.C.; Liao, J.W.; Chou, S.T. Antioxidant activity of graptopetalum paraguayense E. Walther leaf extract counteracts oxidative stress induced by ethanol and carbon tetrachloride co-induced hepatotoxicity in rats. Antioxidants 2019, 8, 251. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Yang, Z.; Zhang, J.; Mu, J.; Zhou, X.; Zhao, X. Liver injury induced by carbon tetrachloride in mice is prevented by the antioxidant capacity of Anji white tea polyphenols. Antioxidants 2019, 8, 64. [Google Scholar] [CrossRef]
- Sukalingam, K.; Ganesan, K.; Xu, B. Protective effect of aqueous extract from the leaves of Justicia tranquebariesis against thioacetamide-induced oxidative stress and hepatic fibrosis in rats. Antioxidants 2018, 7, 78. [Google Scholar] [CrossRef]
- Ganesan, K.; Sukalingam, K.; Xu, B. Solanum trilobatum L. ameliorate thioacetamide-induced oxidative stress and hepatic damage in albino rats. Antioxidants 2017, 6, 68. [Google Scholar] [CrossRef]
- Singh, D.; Arya, P.V.; Aggarwal, V.P.; Gupta, R.S. Evaluation of Antioxidant and Hepatoprotective Activities of Moringa oleifera Lam. Leaves in Carbon Tetrachloride-Intoxicated Rats. Antioxidants 2014, 3, 569–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondi, C.D.; McKeon, R.M.; Bennett, J.M.; Ignatius, P.F.; Brydon, L.; Jockers, R.; Melan, M.A.; Witt-Enderby, P.A. MT1 melatonin receptor internalization underlies melatonin-induced morphologic changes in chinese hamster ovary cells and these processes are depenet on Gi proteins, MEK 1/2 and microtubule modulation. J. Pineal Res. 2008, 44, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Landsman, T.; Waxman, D.J. Role of the cytokine-induced SH2 domain-containing protein CIS in growth hormone receptor internalization. J. Biol. Chem. 2005, 280, 37471–37480. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Posner, B.I.; Balbis, A. Compartmentalization of epidermal growth factor receptor in liver plasma membrane. J. Cell Biochem. 2009, 107, 96–103. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Baseline Values | End of Shock | End of Experiment |
|---|---|---|---|
| Hb [g/dL] | |||
| sham/vehicle | 10.7 ± 0.4 | 10.1 ± 0.4 | 10.1 ± 0.5 |
| sham/mel | 10.9 ± 0.4 | 10.8 ± 0.4 | 10.3 ± 0.6 |
| sham/rml | 11.2 ± 0.4 | 10.6 ± 0.4 | 10.1 ± 0.4 |
| shock/vehicle | 10.3 ± 0.3 | * 6.4 ± 0.5 | # 8.4 ± 0.4 |
| shock/mel | 11.2 ± 0.4 | * 6.3 ± 0.4 | # 8.7 ± 0.4 |
| shock/rml | 10.7 ± 0.3 | * 6.6 ± 0.4 | # 8.5 ± 0.5 |
| Lactate [mmol/L] | |||
| sham/vehicle | 1.7 ± 0.2 | 1.6 ± 0.3 | 1.7 ± 0.1 |
| sham/mel | 1.5 ± 0.3 | 1.8 ± 0.03 | 1.7 ± 0.4 |
| sham/rml | 1.6 ± 0.3 | 1.7 ± 0.3 | 1.5 ± 0.3 |
| shock/vehicle | 1.5 ± 0.3 | * 8.5 ± 0.6 | # 1.9 ± 0.3 |
| shock/mel | 1.9 ± 0.2 | * 9.0 ± 0.6 | # 2.0 ± 0.2 |
| shock/rml | 1.5 ± 0.3 | * 7.7 ± 0.8 | # 1.6 ± 0.4 |
| pH | |||
| sham/vehicle | 7.37 ± 0.02 | 7.39 ± 0.02 | 7.40 ± 0.02 |
| sham/mel | 7.37 ± 0.02 | 7.39 ± 0.02 | 7.42 ± 0.02 |
| sham/rml | 7.36 ± 0.02 | 7.39 ± 0.03 | 7.41 ± 0.03 |
| shock/vehicle | 7.37 ± 0.02 | 7.27 ± 0.02 | 7.34 ± 0.03 |
| shock/mel | 7.37 ± 0.02 | 7.28 ± 0.03 | 7.34 ± 0.03 |
| shock/rml | 7.37 ± 0.02 | 7.26 ± 0.03 | 7.35 ± 0.03 |
| BE [mmol/l] | |||
| sham/vehicle | −1.6 ± 0.8 | −1.8 ± 0.5 | −3.8 ± 1.2 |
| sham/mel | −1.2 ± 1.1 | −1.9 ± 1.0 | −4.1 ± 0.9 |
| sham/rml | −1.0 ± 1.3 | −1.9 ± 1.1 | −3.3 ± 1.7 |
| shock/vehicle | −1.6 ± 1.2 | * −11.4 ± 2.0 | # −6.1 ± 1.8 |
| shock/mel | −.2 ± 1.1 | * −11.9 ± 1.8 | # −5.9 ± 1.2 |
| shock/rml | −1.8 ± 0.9 | * −12.3 ± 1.8 | # −6.4 ± 1.5 |
| Group | Baseline | 45 min | 95 min | 150 min | 210 min |
|---|---|---|---|---|---|
| sham/vehicle | 0.07 ± 0.04 | 0.05 ± 0.05 | 0.15 ± 0.10 | 0.12 ± 0.05 | 0.05 ± 0.04 |
| sham/mel | 0.06 ± 0.04 | 0.04 ± 0.02 | 17.90 ± 1.81 # | 11.39 ± 1.6 # | 1.19 ± 0.52 # |
| sham/rml | 0.09 ± 0.10 | 0.08 ± 0.05 | 0.10 ± 0.04 | 0.11 ± 0.04 | 0.08 ± 0.06 |
| shock/vehicle | 0.04 ± 0.04 | 0.10 ± 0.05 | 1.62 ± 0.76 * | 0.05 ± 0.05 | 0.04 ± 0.01 |
| shock/mel | 0.07 ± 0.06 | 0.05 ± 0.04 | 18.47 ± 1.43 # | 11.80 ± 1.30 # | 2.25 ± 0.70 # |
| shock/rml | 0.05 ± 0.04 | 0.04 ± 0.04 | 1.90 ± 0.54 * | 0.04 ± 0.05 | 0.10 ± 0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mathes, A.M.; Heymann, P.; Ruf, C.; Huhn, R.; Hinkelbein, J.; Volk, T.; Fink, T. Endogenous and Exogenous Melatonin Exposure Attenuates Hepatic MT1 Melatonin Receptor Protein Expression in Rat. Antioxidants 2019, 8, 408. https://doi.org/10.3390/antiox8090408
Mathes AM, Heymann P, Ruf C, Huhn R, Hinkelbein J, Volk T, Fink T. Endogenous and Exogenous Melatonin Exposure Attenuates Hepatic MT1 Melatonin Receptor Protein Expression in Rat. Antioxidants. 2019; 8(9):408. https://doi.org/10.3390/antiox8090408
Chicago/Turabian StyleMathes, Alexander M., Paul Heymann, Christian Ruf, Ragnar Huhn, Jochen Hinkelbein, Thomas Volk, and Tobias Fink. 2019. "Endogenous and Exogenous Melatonin Exposure Attenuates Hepatic MT1 Melatonin Receptor Protein Expression in Rat" Antioxidants 8, no. 9: 408. https://doi.org/10.3390/antiox8090408
APA StyleMathes, A. M., Heymann, P., Ruf, C., Huhn, R., Hinkelbein, J., Volk, T., & Fink, T. (2019). Endogenous and Exogenous Melatonin Exposure Attenuates Hepatic MT1 Melatonin Receptor Protein Expression in Rat. Antioxidants, 8(9), 408. https://doi.org/10.3390/antiox8090408