Radioprotective Agents and Enhancers Factors. Preventive and Therapeutic Strategies for Oxidative Induced Radiotherapy Damages in Hematological Malignancies


Abstract
:1. Introduction
Radiotherapy in Hematologic Malignancies
2. Radiotherapy and Oxidative Stress
3. Radioprotective Agents
4. Enhancers of Radiotherapy Activity in Hematologic Neoplasms
5. Future Perspectives
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tseng, Y.D.; Ng, A.K. Hematologic Malignancies. Hematol. Oncol. Clin. N. Am. 2020, 34, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Galunić-Bilić, L.; Šantek, F. Infradiaphragmal Radiotherapy in Patients with Lymphoma: Volume Definition and Side Effects. Acta Clin. Croat. 2018, 57, 554–560. [Google Scholar] [CrossRef] [Green Version]
- Bosch, A.; Frias, Z. Radiotherapy in the treatment of multiple myeloma. Int. J. Radiat. Oncol. Biol. Phys. 1988, 15, 1363–1369. [Google Scholar] [CrossRef]
- Kilciksiz, S.; Celik, O.K.; Pak, Y.; Demiral, A.N.; Pehlivan, M.; Orhan, O.; Tokatli, F.; Agaoglu, F.; Zincircioglu, B.; Atasoy, B.M.; et al. Clinical and prognostic features of plasmacytomas: A multicenter study of Turkish Oncology Group–Sarcoma Working Party. Am. J. Hematol. 2008, 83, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Dispenzieri, A.; Witzig, T.E.; Russell, S.J. Enhancing the therapeutic index of radiation in multiple myeloma. Drug Discov. Today Dis. Mech. 2006, 3, 515–522. [Google Scholar] [CrossRef]
- Chatterjee, M.; Chakraborty, T.; Tassone, P. Multiple myeloma: Monoclonal antibodies–based immunotherapeutic strategies and targeted radiotherapy. Eur. J. Cancer 2006, 42, 1640–1652. [Google Scholar] [CrossRef]
- Wong, J.Y.; Liu, A.; Schultheiss, T.; Popplewell, L.; Stein, A.; Rosenthal, J.; Essensten, M.; Forman, S.; Somlo, G. Targeted total marrow irradiation using three dimensional image-guided tomographic intensity–modulated radiation therapy: An alternative to standard total body irradiation. Biol. Blood Marrow Transplant. 2006, 12, 306–315. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.Y.; Rosenthal, J.; Liu, A.; Schultheiss, T.; Forman, S.; Somlo, G. Image-guided total-marrow irradiation using helical tomotherapy in patients with multiple myeloma and acute leukemia undergoing hematopoietic cell transplantation. Int. J. Radiat. Oncol. Biol. Phys. 2009, 73, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Anderson, P.M.; Wiseman, G.A.; Dispenzieri, A.; Arndt, C.A.; Hartmann, L.C.; Smithson, W.A.; Mullan, B.P.; Bruland, O.S. High-dose samarium-153 ethylene diamine tetramethylene phosphonate: Low toxicity of skeletal irradiation in patients with osteosarcoma and bone metastases. J. Clin. Oncol. 2002, 20, 189–196. [Google Scholar] [CrossRef]
- Dispenzieri, A.; Wiseman, G.A.; Lacy, M.Q.; Geyer, S.; Litzow, M.R.; Tefferi, A.; Inwards, D.J.; Micallef, I.N.; Ansell, S.; Gastineau, D.A.; et al. A phase II study of high dose 153-samarium EDTMP (153-sm EDMTP) and melphalan for peripheral stem cell transplantation (PBSCT) in multiple myeloma (MM). Blood 2003, 102, 982. [Google Scholar]
- Wilky, B.A.; Loeb, D.M. Beyond Palliation: Therapeutic Applications of 153Samarium-EDTMP. Clin. Exp. Pharmacol. 2013, 3, 1000131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abruzzese, E.; Iuliano, F.; Trawinska, M.M.; DiMaio, M. 153Sm: Its use in multiple myeloma and report of a clinical experience. Expert Opin. Investig. Drugs 2008, 17, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Marchand, V.; Decaudin, D.; Servois, V.; Kirova, Y.M. Concurrent radiation therapy and lenalidomide in myeloma patient. Radiother. Oncol. 2008, 87, 152–153. [Google Scholar] [CrossRef] [PubMed]
- Berenson, J.R.; Yellin, O.; Patel, R.; Duvivier, H.; Nassir, Y.; Mapes, R.; Abaya, C.D.; Swift, R.A. A phase I study of samarium lexidronam/bortezomib combination therapy for the treatment of relapsed or refractory multiple myeloma. Clin. Cancer Res. 2009, 15, 1069–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berges, O.; Decaudin, D.; Servois, V.; Kirova, Y.M. Concurrent radiation therapy and bortezomib in myeloma patient. Radiother. Oncol. 2008, 86, 290–292. [Google Scholar] [CrossRef]
- Bakst, R.L.; Dabaja, B.S.; Specht, L.K.; Yahalom, J. Use of radiation in extramedullary leukemia/chloroma: Guidelines from the International Lymphoma Radiation Oncology Group. Int. J. Radiat. Oncol. Biol. Phys. 2018, 102, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Karihtala, P.; Winqvist, R.; Syvaoja, J.E.; Kinnula, V.L.; Soini, Y. Increasing oxidative damage and loss of mismatch repair enzymes during breast carcinogenesis. Eur. J. Cancer 2006, 42, 2653–2659. [Google Scholar] [CrossRef] [PubMed]
- Mates, J.M.; Segura, J.A.; Alonso, F.J.; Marquez, J. Intracellular redox status and oxidative stress: Implications for cell proliferation, apoptosis, and carcinogenesis. Arch. Toxicol. 2008, 82, 273–299. [Google Scholar] [CrossRef]
- Karihtala, P.; Soini, Y. Reactive oxygen species and antioxidant mechanisms in human tissues and their relation to malignancies. Acta Pathol. Microbiol. Immunol. Scand. 2007, 115, 81–103. [Google Scholar] [CrossRef]
- Karihtala, P.; Kauppila, S.; Soini, Y.; Jukkola-Vuorinen, A. Oxidative stress and counteracting mechanisms in hormone receptor positive, triple-negative and basal-like breast carcinomas. BMC Cancer 2011, 11, 262. [Google Scholar] [CrossRef] [Green Version]
- Salzman, R.; Pacal, L.; Kankova, K.; Tomandl, J.; Horakova, Z.; Tothova, E.; Kostrica, R. High perioperative level of oxidative stress as a prognostic tool for identifying patients with a high risk of recurrence of head and neck squamous cell carcinoma. Int. J. Clin. Oncol. 2010, 15, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Imbesi, S.; Musolino, C.; Allegra, A.; Saija, A.; Morabito, F.; Calapai, G.; Gangemi, S. Oxidative stress in oncohematologic diseases: An update. Expert Rev. Hematol. 2013, 6, 317–325. [Google Scholar] [CrossRef]
- Gangemi, S.; Allegra, A.; Aguennouz, M.; Alonci, A.; Speciale, A.; Cannavò, A.; Cristani, M.; Russo, S.; Spatari, G.; Alibrandi, A.; et al. Relationship between advanced oxidation protein products, advanced glycation end products, and S-nitrosylated proteins with biological risk and MDR-1 polymorphisms in patients affected by B-chronic lymphocytic leukemia. Cancer Investig. 2012, 30, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Musolino, C.; Allegra, A.; Alonci, A.; Saija, A.; Russo, S.; Cannavò, A.; Cristani, M.; Centorrino, R.; Saitta, S.; Alibrandi, A.; et al. Carbonyl group serum levels are associated with CD38 expression in patients with B chronic lymphocytic leukemia. Clin. Biochem. 2011, 44, 1487–1490. [Google Scholar] [CrossRef] [PubMed]
- Peroja, P.; Pasanen, A.K.; Haapasaari, K.M.; Jantunen, E.; Soini, Y.; Turpeenniemi-Hujanen, T.; Bloigu, R.; Lilja, L.; Kuittinen, O.; Karihtala, P. Oxidative stress and redox state-regulating enzymes have prognostic relevance in diffuse large B-cell lymphoma. Exp. Hematol. Oncol. 2012, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F.L.; Zhang, W.G.; Wei, Y.C.; Meng, S.; Bai, G.G.; Wang, B.Y.; Yang, H.Y.; Tian, W.; Meng, X.; Zhang, H.; et al. Involvement of oxidative stress in the relapse of acute myeloid leukemia. J. Biol. Chem. 2010, 285, 15010–15015. [Google Scholar] [CrossRef] [Green Version]
- Bur, H.; Haapasaari, K.M.; Turpeenniemi-Hujanen, T.; Kuittinen, O.; Auvinen, P.; Marin, K.; Koivunen, P.; Sormunen, R.; Soini, Y.; Karihtala, P. Oxidative stress markers and mitochondrial antioxidant enzyme expression are increased in aggressive Hodgkin lymphomas. Histopathology 2014, 65, 319–327. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Rahgoshai, S.; Mohammadi, M.; Refahi, S.; Oladghaffari, M.; Aghamiri, S. Protective effects of IMOD and cimetidine against radiation induced cellular damage. J. Biomed. Phys. Eng. 2017, 8, 133–140. [Google Scholar]
- Azzam, E.I.; Jay-Gerin, J.P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Popgeorgiev, N.; Jabbour, L.; Gillet, G. Subcellular localization and dynamics of the Bcl-2 family of proteins. Front. Cell Dev. Biol. 2018, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, C.F.A.; Wong-Brown, M.W.; Bowden, N.A. BCL-2 family isoforms in apoptosis and cancer. Cell Death Dis. 2019, 10, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohl, S.Ö.-G.; Agostino, M.; Dharmarajan, A.; Pervaiz, S. Cross talk between cellular redox state and the antiapoptotic protein Bcl-2. Antioxid. Redox Signal. 2018, 29, 1215–1236. [Google Scholar] [CrossRef] [PubMed]
- Iliakis, G.; Wang, H.; Perrault, A.R.; Boecker, W.; Rosidi, B.; Windhofer, F.; Wu, W.; Guan, J.; Terzoudi, G.; Antelias, G. Mechanisms of DNA double strand break repair and chromosome aberration formation. Cytogenet. Genome Res. 2004, 104, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Purkayastha, S.; Milligan, J.R.; Bernhard, W.A. On the chemical yield of base lesions, strand breaks, and clustered damage generated in plasmid DNA by the direct effect of × rays. Radiat. Res. 2007, 168, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Yakes, F.M.; van Houten, B. Mitochondrial DNA damage is more extensive and persists longer than nuclear DNA damage in human cells following oxidative stress. Proc. Natl. Acad. Sci. USA 1997, 94, 514–519. [Google Scholar] [CrossRef] [Green Version]
- Rodemann, H.P.; Blaese, M.A. Responses of normal cells to ionizing radiation. Semin. Radiat. Oncol. 2007, 17, 81–88. [Google Scholar] [CrossRef]
- Larsen, N.B.; Rasmussen, M.; Rasmussen, L.J. Nuclear and mitochondrial DNA repair: Similar pathways? Mitochondrion 2005, 5, 89–108. [Google Scholar] [CrossRef]
- Yang, J.L.; Weissman, L.; Bohr, V.A.; Mattson, M.P. Mitochondrial DNA damage and repair in neurodegenerative disorders. DNA Repair 2008, 7, 1110–1120. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Maguire, D.; Swarts, S.; Sun, W.; Yang, S.; Wang, W.; Liu, C.; Zhang, M.; Zhang, D.; Zhang, L.; et al. Replication of murine mitochondrial DNA following irradiation. Adv. Exp. Med. Biol. 2009, 645, 43–48. [Google Scholar]
- Cortopassi, G.A.; Arnheim, N. Detection of a specific mitochondrial DNA deletion in tissues of older humans. Nucleic Acids Res. 1990, 18, 6927–6933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, K.J.; Reeve, A.K.; Samuels, D.C.; Chinnery, P.F.; Blackwood, J.K.; Taylor, R.W.; Wanrooij, S.; Spelbrink, J.N.; Lightowlers, R.N.; Turnbull, D.M. What causes mitochondrial DNA deletions in human cells? Nat. Genet. 2008, 40, 275–279. [Google Scholar] [CrossRef]
- Gerhard, G.S.; Benko, F.A.; Allen, R.G.; Tresini, M.; Kalbach, A.; Cristofalo, V.J.; Gocke, C.D. Mitochondrial DNA mutation analysis in human skin fibroblasts from fetal, young, and old donors. Mech Ageing Dev. 2002, 123, 155–166. [Google Scholar] [CrossRef]
- Chabi, B.; Mousson de Camaret, B.; Chevrollier, A.; Boisgard, S.; Stepien, G. Random mtDNA deletions and functional consequence in aged human skeletal muscle. Biochem. Biophys. Res. Commun. 2005, 332, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Pavicic, W.H.; Richard, S.M. Correlation analysis between mtDNA 4977-bp deletion and ageing. Mutat. Res. 2009, 670, 99–102. [Google Scholar] [CrossRef]
- Wen, Q.; Hu, Y.; Ji, F.; Qian, G. Mitochondrial DNA alterations of peripheral lymphocytes in acute lymphoblastic leukemia patients undergoing total body irradiation therapy. Radiat. Oncol. 2011, 6, 133. [Google Scholar] [CrossRef] [Green Version]
- Soyal, D.; Jindal, A.; Singh, I.; Goyal, P. Protective capacity of Rosemary extract against radiation induced hepatic injury in mice. Int. J. Radiat. Res. 2007, 4, 161–168. [Google Scholar]
- Janssen, L.; Bosman, C.B.; van Duijn, W.; Oostendorp-van de Ruit, M.M.; Kubben, F.J.; Griffioen, G.; Lamers, C.B.; van Krieken, J.H.; van de Velde, C.J.; Verspaget, H.W. Superoxide dismutases in gastric and esophageal cancer and the prognostic impact in gastric cancer. Clin. Cancer Res. 2000, 6, 3183–3192. [Google Scholar]
- Hall, S.; Rudrawar, S.; Zunk, M.; Bernaitis, N.; Arora, D.; McDermott, C.M.; Anoopkumar-Dukie, S. Protection against Radiotherapy-Induced Toxicity. Antioxidants 2016, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Shi, H.; Ren, L.; Gou, H.; Gong, D.; Gao, X.; Huang, N. Enhancing the anti-colon cancer activity of quercetin by self-assembled micelles. Int. J. Nanomed. 2015, 10, 2051–2063. [Google Scholar] [CrossRef] [Green Version]
- Rusin, A.; Krawczyk, Z.; Grynkiewicz, G.; Gogler, A.; Zawisza-Puchałka, J.; Szeja, W. Synthetic derivatives of genistein, their properties and possible applications. Acta Biochim. Pol. 2010, 57, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szejk, M.; Poplawski, T.; Czubatka-Bienkowska, A.; Olejnik, A.K.; Pawlaczyk-Graja, I.P.; Gancarz, R.; Zbikowska, H.M. A comparative study on the radioprotective potential of the polyphenolic glycoconjugates from medicinal plants of Rosaceae and Asteraceae families versus their aglycones. J. Photochem. Photobiol. B 2017, 171, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Szejk-Arendt, M.; Czubak-Prowizor, K.; Macieja, A.; Poplawski, T.; Olejnik, A.K.; Pawlaczyk-Graja, I.; Gancarz, R.; Zbikowska, H.M. Polyphenolic-polysaccharide conjugates from medicinal plants of Rosaceae/Asteraceae family protect human lymphocytes but not myeloid leukemia K562 cells against radiation-induced death. Int. J. Biol. Macromol. 2020, 156, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Mozdarani, H.; Salimi, M.; Froughizadeh, M. Effect of cimetidine and famotidine on survival of lethally gamma irradiated mice. Iran. J. Radiat. Res. 2008, 5, 187–194. [Google Scholar]
- Estaphan, S.; Abdel-Malek, R.; Rashed, L.; Mohamed, E.A. Cimetidine a promising radio-protective agent through modulating Bax/Bcl2 ratio: An in vivo study in male rats. J. Cell Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Yamini, K.; Gopal, V. Natural radioprotective agents against ionizing radiation—An overview. Int. J. Pharm. Tech. Res. 2010, 2, 1421–1426. [Google Scholar]
- Wegener, T.; Wagner, H. The active components and the pharmacological multi-target principle of STW 5 (Iberogast). Phytomedicine 2006, 13, 20–35. [Google Scholar] [CrossRef]
- Khayyal, M.T.; Abdel-Naby, D.H.; Abdel-Aziz, H.; El-Ghazaly, M.A. A multi-component herbal preparation, STW 5, shows anti-apoptotic effects in radiation induced intestinal mucositis in rats. Phytomedicine 2014, 21, 1390–1399. [Google Scholar] [CrossRef] [Green Version]
- Wadie, W.; Abdel-Aziz, H.; Zaki, H.F.; Kelber, O.; Weiser, D.; Khayyal, M.T. STW 5 is effective in dextran sulfate sodium-induced colitis in rats. Int. J. Colorectal Dis. 2012, 27, 1445–1453. [Google Scholar] [CrossRef] [Green Version]
- Michael, S.; Kelber, O.; Hauschildt, S.; Spanel-Borowski, K.; Nieber, K. 2009. Inhibition of inflammation-induced alterations in rat small intestine by the herbal preparations STW 5 and STW 6. Phytomedicine 2009, 16, 161–171. [Google Scholar] [CrossRef]
- Burdelya, L.G.; Krivokrysenko, V.I.; Tallant, T.C.; Strom, E.; Gleiberman, A.S.; Gupta, D.; Kurnasov, O.V.; Fort, F.L.; Osterman, A.L.; Didonato, J.A.; et al. An agonist of toll-like receptor 5 has radioprotective activity in mouse and primate models. Science 2008, 320, 226–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toshkov, I.A.; Gleiberman, A.S.; Mett, V.L.; Hutson, A.D.; Singh, A.K.; Gudkov, A.V.; Burdelya, L.G. Mitigation of Radiation-Induced Epithelial Damage by the TLR5 Agonist Entolimod in a Mouse Model of Fractionated Head and Neck Irradiation. Radiat. Res. 2017, 187, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Leigh, N.D.; Bian, G.; Ding, X.; Liu, H.; Aygun-Sunar, S.; Burdelya, L.G.; Gudkov, A.V.; Cao, X. A flagellin-derived toll-like receptor 5 agonist stimulates cytotoxic lymphocyte-mediated tumor immunity. PLoS ONE 2014, 9, e85587. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Brackett, C.M.; Morales-Tirado, V.M.; Li, Z.; Zhang, Q.; Wilson, M.W.; Benjamin, C.; Harris, W.; Waller, E.K.; Gudkov, A.V.; et al. The Toll-like receptor 5 agonist entolimod suppresses hepatic metastases in a murine model of ocular melanoma via an NK cell-dependent mechanism. Oncotarget 2016, 7, 2936–2950. [Google Scholar] [CrossRef] [Green Version]
- Brackett, C.M.; Kojouharov, B.; Veith, J.; Greene, K.F.; Burdelya, L.G.; Gollnick, S.O.; Abrams, S.I.; Gudkov, A.V. Toll-like receptor-5 agonist, entolimod, suppresses metastasis and induces immunity by stimulating an NK-dendritic-CD8+ T-cell axis. Proc. Natl. Acad. Sci. USA 2016, 113, E874–E883. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ge, C.; Yang, L.; Wang, R.; Lu, Y.; Gao, Y.; Li, Z.; Wu, Y.; Zheng, X.; Wang, Z.; et al. CBLB502, an agonist of Toll-like receptor 5, has antioxidant and scavenging free radicals activities in vitro. Int. J. Biol. Macromol. 2016, 82, 97–103. [Google Scholar] [CrossRef]
- Bai, H.; Sun, F.; Yang, G.; Wang, L.; Zhang, Q.; Zhang, Q.; Zhan, Y.; Chen, J.; Yu, M.; Li, C.; et al. CBLB502, a Toll-like receptor 5 agonist, offers protection against radiation-induced male reproductive system damage in mice. Biol. Reprod. 2019, 100, 281–291. [Google Scholar] [CrossRef]
- Elkady, A.A.; Ibrahim, I.M. Protective effects of erdosteine against nephrotoxicity caused by gamma radiation in male albino rats. Hum. Exp. Toxicol. 2016, 35, 21–28. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Hu, Y.Y.; Hu, X.F.; Cheng, L.X. The conditioned medium of human mesenchymal stromal cells reduces irradiation-induced damage in cardiac fibroblast cells. J. Radiat. Res. 2018, 59, 555–564. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Wang, Z.; Zheng, R. Efficacy of calf spleen extract injection combined with chemotherapy in the treatment of advanced gastric cancer. J. Bengbu Med. Coll. 2010, 779, 781. [Google Scholar]
- Liu, Z.; Zhang, X.; Ma, J.; Wang, L.; Yang, Y. Calf spleen extract injection combined with docetaxel Saijiakapei gemcitabine treatment of patients with advanced breast cancer program in clinical research. Cancer Res. Prev. 2014, 132, 122–130. [Google Scholar]
- Svirnovskiĭ, A.I.; Shimanskaia, T.V.; Bakun, A.V. Protivoluchevoe deĭstvie ékstrakta regeneriruiushcheĭ selezenki. The radioprotective action of an extract of regenerating spleen. Radiobiologiia 1993, 33, 141–147. [Google Scholar] [PubMed]
- Djordjevic, A.; Bogdanovic, G.; Dobric, S. Fullerenes in biomedicine. J. Buon 2006, 11, 391–404. [Google Scholar] [PubMed]
- Trajković, S.; Dobrić, S.; Jaćević, V.; Dragojević-Simić, V.; Milovanović, Z.; Dordević, A. Tissue-protective effects of fullerenol C60(OH)24 and amifostine in irradiated rats. Colloids Surf. B Biointerfaces 2007, 58, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Chapman, T.R.; Kinsella, T.J. Ribonucleotide reductase inhibitors: A new look at an old target for radiosensitization. Front. Oncol. 2012, 1, 56. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.D.; Hammond, E.M.; Higgins, G.S.; Petersson, K. Ultra-High Dose Rate (FLASH) Radiotherapy: Silver Bullet or Fool’s Gold? Front. Oncol. 2020, 9, 1563. [Google Scholar] [CrossRef] [Green Version]
- Jafarzadeh, A.; Nemati, M.; Khorramdelazad, H.; Hassan, Z.M. Immunomodulatory properties of cimetidine: Its therapeutic potentials for treatment of immune-related diseases. Int. Immunopharmacol. 2019, 70, 156–166. [Google Scholar] [CrossRef]
- Little, M.H. Regrow or repair: Potential regenerative therapies for the kidney. J. Am. Soc. Nephrol. 2006, 17, 2390–2401. [Google Scholar] [CrossRef] [Green Version]
- Linard, C.; Ropenga, A.; Vozenin-Brotons, M.C.; Chapel, A.; Mathe, D. Abdominal irradiation increases inflammatory cytokine expression and activates NF-kappa B in ratileal muscularis layer. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, 556–565. [Google Scholar] [CrossRef]
- Wong, P.C.; Dodd, M.J.; Miaskowski, C.; Paul, S.M.; Bank, K.A.; Shiba, G.H.; Facione, N. Mucositis pain induced by radiation therapy: Prevalence, severity and use of self-care behaviors. J. Pain Symptom Manag. 2006, 32, 27–37. [Google Scholar] [CrossRef]
- Duca, Y.; di Cataldo, A.; Russo, G.; Cannata, E.; Burgio, G.; Compagnone, M.; Alamo, A.; Condorelli, R.A.; La Vignera, S.; Calogero, A.E. Testicular Function of Childhood Cancer Survivors: Who Is Worse? J. Clin. Med. 2019, 8, 2204. [Google Scholar] [CrossRef] [Green Version]
- Meistrich, M.L. Effects of chemotherapy and radiotherapy on spermatogenesis. Eur. Urol. 1993, 23, 136–141. [Google Scholar] [CrossRef]
- Hermann, R.M.; Henkel, K.; Christiansen, H.; Vorwerk, H.; Hille, A.; Hess, C.F.; Schmidberger, H. Testicular dose and hormonal changes after radiotherapy of rectal cancer. Radiother. Oncol. 2005, 75, 83–88. [Google Scholar] [CrossRef]
- Shen, H.; Ong, C. Detection of oxidative DNA damage in human sperm and its association with sperm function and male infertility. Free Radic. Biol. Med. 2000, 28, 529–536. [Google Scholar] [CrossRef]
- Ash, P. The influence of radiation on fertility in man. Br. J. Radiol. 1980, 53, 271–278. [Google Scholar] [CrossRef]
- Albuquerque, A.V.; Almeida, F.R.; Weng, C.C.; Shetty, G.; Meistrich, M.L.; Chiarini-Garcia, H. Spermatogonial behavior in rats during radiation-induced arrest and recovery after hormone suppression. Reproduction 2013, 146, 363–376. [Google Scholar] [CrossRef] [Green Version]
- Chuai, Y.; Gao, F.; Li, B.; Zhao, L.; Qian, L.; Cao, F.; Wang, L.; Sun, X.; Cui, J.; Cai, J. Hydrogen-rich saline attenuates radiation-induced male germ cell loss in mice through reducing hydroxyl radicals. Biochem. J. 2012, 442, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Wang, H.; Wu, Z.B.; Zhao, J.; Zhang, S.; Li, W. Protection of murine spermatogenesis against ionizing radiation-induced testicular injury by a green tea polyphenol. Biol. Reprod. 2015, 92, 6. [Google Scholar] [CrossRef]
- Kim, J.; Lee, S.; Jeon, B.; Jang, W.; Moon, C.; Kim, S. Protection of spermatogenesis against gamma ray-induced damage by granulocytecolony-stimulating factor in mice. Andrologia 2011, 43, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.D.; Maquart, F.X.; Monboisse, J.C. Ionizing radiations and collagen metabolism: From oxygen free radicals to radio-induced late fibrosis. Radiat. Phys. Chem. 2005, 72, 381–386. [Google Scholar] [CrossRef]
- Cohen, E.P.; Robbins, M.E. Radiation nephropathy. Semin. Nephrol. 2003, 23, 486–499. [Google Scholar] [CrossRef]
- Teixeira, V.P.; Boim, M.A.; Segreto, H.R.; Schor, N. Acute, subacute, and chronic x-ray effects on glomerular hemodynamics in rats. Ren. Fail. 1994, 16, 457–470. [Google Scholar] [CrossRef]
- Withers, H.R.; Mason, K.A.; Thames, H.D., Jr. Late radiation response of kidney assayed by tubule-cell survival. Br. J. Radiol. 1986, 59, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Flora, S.J. Role of free radicals and antioxidants in health and disease. Cell. Mol. Biol. 2007, 53, 1–2. [Google Scholar]
- Gobé, G.C.; Axelsen, R.A.; Harmon, B.V.; Allan, D.J. Cell death by apoptosis following X-irradiation of the foetal and neonatal rat kidney. Int. J. Radiat. Biol. 1988, 54, 567–576. [Google Scholar] [CrossRef]
- Moulder, J.E.; Fish, B.L.; Regner, K.R.; Cohen, E.P.; Raife, T.J. Retinoic acid exacerbates experimental radiation nephropathy. Radiat. Res. 2002, 157, 199–203. [Google Scholar] [CrossRef]
- Robbins, M.E.; O’Malley, Y.; Zhao, W.; Davis, C.S.; Bonsib, S.M. The role of tubulointerstitium in radiation-induced renal fibrosis. Radiat. Res. 2001, 155, 481–489. [Google Scholar] [CrossRef]
- Hu, S.; Chen, Y.; Li, L.; Chen, J.; Wu, B.; Zhou, X.; Zhi, G.; Li, Q.; Wang, R.; Duan, H.; et al. Effects of adenovirus-mediated delivery of the human hepatocyte growth factor gene in experimental radiation-induced heart disease. Int. J. Radiat. Oncol. Biol. Phys. 2009, 75, 1537–1544. [Google Scholar] [CrossRef]
- Blijlevens, N.; Sonis, S. Palifermin (recombinant keratinocyte growth factor-1): A pleiotropic growth factor with multiple biological activities in preventing chemotherapy- and radiotherapy-induced mucositis. Ann. Oncol. 2007, 18, 817–826. [Google Scholar] [CrossRef]
- Lauritano, D.; Petruzzi, M.; Di Stasio, D.; Lucchese, A. Clinical effectiveness of palifermin in prevention and treatment of oral mucositis in children with acute lymphoblastic leukaemia: A case-control study. Int. J. Oral Sci. 2014, 6, 27–30. [Google Scholar] [CrossRef]
- Spielberger, R.; Stiff, P.; Bensinger, W.; Gentile, T.; Weisdorf, D.; Kewalramani, T.; Shea, T.; Yanovich, S.; Hansen, K.; Noga, S.; et al. Palifermin for oral mucositis after intensive therapy for hematologic cancers. N. Engl. J. Med. 2004, 351, 2590–2598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shankar, B.; Kumar, S.S.; Sainis, K.B. Generation of reactive oxygen species and radiation response in lymphocytes and tumour cells. Radiat. Res. 2003, 160, 478–487. [Google Scholar] [CrossRef]
- Komatsu, W.; Ishihara, K.; Murata, M.; Saito, H.; Shinohara, K. Docosahexaenoic acid suppresses nitric oxide production and inducible nitric oxide synthase expression in interferon-gamma plus lipopolysaccharide stimulated murine macrophages by inhibiting the oxidative stress. Free Radic. Biol. Med. 2003, 34, 1006–1016. [Google Scholar] [CrossRef]
- Sneddon, A.A.; Wu, H.C.; Farquharson, A.; Grant, I.; Arthur, J.R.; Rotondo, D.; Choe, S.-N.; Wahle, K.W.J. Regulation of selenoprotein GP×4 expression and activity in human endothelial cells by fatty acids, cytokines and antioxidants. Atherosclerosis 2003, 171, 57–65. [Google Scholar] [CrossRef]
- Wen, B.; Deutsch, E.; Opolon, P.; Auperin, A.; Frascogna, V.; Connault, E.; Bourhis, J. n-3 polyunsaturated fatty acids decrease mucosal/epidermal reactions and enhance antitumour effect of ionising radiation with inhibition of tumour angiogenesis. Br. J. Cancer 2003, 89, 1102–1107. [Google Scholar] [CrossRef] [Green Version]
- Minami, Y.; Miyata, H.; Doki, Y.; Yano, M.; Yamasaki, M.; Takiguchi, S.; Fujiwara, Y.; Yasuda, T.; Monden, M. omega-3 Fatty acid-containing diet (Racol) reduces toxicity of chemoradiation therapy for patients with esophageal cancer. Gan Kagaku Ryoho 2008, 35, 437–440. [Google Scholar]
- Briehl, M.M.; Cotgreave, I.A.; Powis, G. Downregulation of the antioxidant defence during glucocorticoid-mediated apoptosis. Cell Death Differ. 1995, 2, 41–46. [Google Scholar]
- Burington, B.; Barlogie, B.; Zhan, F.; Crowley, J.; Shaughnessy, J.D., Jr. Tumor cell gene expression changes following short-term in vivo exposure to single agent chemotherapeutics are related to survival in multiple myeloma. Clin. Cancer Res. 2008, 14, 4821–4829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bera, S.; Greiner, S.; Choudhury, A.; Dispenzieri, A.; Spitz, D.R.; Russell, S.J.; Apollina Goel, A. Dexamethasone-induced oxidative stress enhances myeloma cell radiosensitization while sparing normal bone marrow hematopoiesis. Neoplasia 2010, 12, 980–992. [Google Scholar] [CrossRef] [Green Version]
- Magda, D.; Lepp, C.; Gerasimchuk, N.; Lee, I.; Sessler, J.L.; Lin, A.; Biaglow, J.E.; Miller, R.A. Redox cycling by motexafin gadolinium enhances cellular response to ionizing radiation by forming reactive oxygen species. Int. J. Radiat. Oncol. Biol. Phys. 2001, 51, 1025–1036. [Google Scholar] [CrossRef]
- Skvortsova, I.; Popper, B.A.; Skvortsov, S.; Saurer, M.; Auer, T.; Moser, R.; Kamleitner, H.; Zwierzina, H.; Lukas, P. Pretreatment with rituximab enhances radiosensitivity of non-Hodgkin’s lymphoma cells. J. Radiat Res. 2005, 46, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skvortsova, I.; Skvortsov, S.; Popper, B.A.; Haidenberger, A.; Saurer, M.; Gunkel, A.R.; Zwierzina, H.; Lukas, P. Rituximab enhances radiation-triggered apoptosis in non-Hodgkin’s lymphoma cells via caspase-dependent and -independent mechanisms. J. Radiat. Res. 2006, 47, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Kapadia, N.S.; Engles, J.M.; Wahl, R.L. In vitro evaluation of radioprotective and radiosensitizing effects of rituximab. J. Nucl. Med. 2008, 49, 674–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fengling, M.; Fenju, L.; Wanxin, W.; Lijia, Z.; Jiandong, T.; Zu, W.; Xin, Y.; Qingxiang, G. Rituximab sensitizes a Burkitt lymphoma cell line to cell killing by X-irradiation. Radiat. Environ. Biophys. 2009, 48, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Zimta, A.A.; Cenariu, D.; Irimie, A.; Magdo, L.; Nabavi, S.M.; Atanasov, A.G.; Berindan-Neagoe, I. The Role of Nrf2 Activity in Cancer Development and Progression. Cancers 2019, 11, 1755. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Lu, H.; Bai, Y. Nrf2 in cancers: A double-edged sword. Cancer Med. 2019, 8, 2252–2267. [Google Scholar] [CrossRef]
- Wang, X.J.; Hayes, J.D.; Henderson, C.J.; Wolf, C.R. Identification of retinoic acid as an inhibitor of transcription factor Nrf2 through activation of retinoic acid receptor alpha. Proc. Natl. Acad. Sci. USA 2007, 104, 19589–19594. [Google Scholar] [CrossRef] [Green Version]
- Do, M.T.; Kim, H.G.; Khanal, T.; Choi, J.H.; Kim, D.H.; Jeong, T.C.; Jeong, H.G. Metformin inhibits heme oxygenase-1 expression in cancer cells through inactivation of Raf-ERK-Nrf2 signaling and AMPK-independent pathways. Toxicol. Appl. Pharmacol. 2013, 271, 229–238. [Google Scholar] [CrossRef]
- Allegra, A.; Innao, V.; Russo, S.; Gerace, D.; Alonci, A.; Musolino, C. Anticancer Activity of Curcumin and Its Analogues: Preclinical and Clinical Studies. Cancer Investig. 2017, 35, 1–22. [Google Scholar] [CrossRef]
- Krishnan, S.; Sandur, S.K.; Shentu, S.; Aggarwal, B.B. Curcumin enhances colorectal cancer cell radiosensitivity by suppressing the radiation-induced nuclear factorkappaB (NF-kB) pathway. Int. J. Radiat. Oncol. Biol. Phys. 2006, 66, S547. [Google Scholar] [CrossRef]
- Jayakumar, S.; Patwardhan, R.S.; Pal, D.; Sharma, D.; Sandur, S.K. Dimethoxycurcumin, a metabolically stable analogue of curcumin enhances the radiosensitivity of cancer cells: Possible involvement of ROS and thioredoxin reductase. Biochem. Biophys. Res. Commun. 2016, 478, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Adams, G.E.; Dewey, D.L. Hydrated electrons and radiobiological sensitization. Biochem. Biophys. Res. Commun. 1963, 12, 473–477. [Google Scholar] [CrossRef]
- Biaglow, J.E.; Varnes, M.E.; Clark, E.P.; Epp, E.R.R. The role of thiols in cellular response to radiation and drugs. Radiat. Res. 1983, 95, 437–455. [Google Scholar] [CrossRef] [PubMed]
- Biaglow, J.E.; Varnes, M.E.; Epp, E.R.; Clark, E.P.; Tuttle, S.; Held, K.D. Role of glutathione in the aerobic radiation response. Int. J. Radiat. Oncol. Biol. Phys. 1989, 16, 1311–1314. [Google Scholar] [CrossRef]
- Biaglow, J.E.; Mitchell, J.B.; Held, K. The importance of peroxide and superoxide in the x-ray response. Int. J. Radiat. Oncol. Biol. Phys. 1992, 22, 665–669. [Google Scholar] [CrossRef]
- Siim, B.G.; van Zijl, P.L.; Brown, J.M. Tirapazamine-induced DNA damage measured using the comet assay correlates with cytotoxicity towards tumour cells in vitro. Br. J. Cancer 1996, 73, 952–960. [Google Scholar] [CrossRef] [Green Version]
- Coleman, C.N.; Beard, C.J.; Hlatky, L. Biochemical modifiers: Hypoxic cell sensitizers. In Radiation Oncology: Technology and Biology; Mauch, P.M., Loeffler, J.S., Eds.; W.B. Saunders: Philadelphia, PA, USA, 1994; pp. 56–89. [Google Scholar]
- Biaglow, J.E.; Varnes, M.E.; Roizen-Towle, L.; Clark, E.P.; Epp, E.R.; Astor, M.B.; Hall, E.J. Biochemistry of reduction of nitro heterocycles. Biochem. Pharmacol. 1986, 35, 77–90. [Google Scholar] [CrossRef]
- Jacobson, B.; Biaglow, J.E.; Fielden, E.M.; Adams, G.E. Respiratory effects and ascorbate reactions with misonidazole and other recently developed drugs. Cancer Clin. Trials 1980, 3, 47–53. [Google Scholar]
- Mengeaud, V.; Nano, J.L.; Fournel, S.; Rampal, P. Effects of eicosapentaenoic acid, gamma-linolenic acid and prostaglandin E1 on three human colon carcinoma cell lines. Prostaglandins Leukot Essent Fatty Acids 1992, 47, 313–319. [Google Scholar] [CrossRef]
- Ng, Y.; Barhoumi, R.; Tjalkens, R.B.; Fan, Y.Y.; Kolar, S.; Wang, N.; Lupton, J.R.; Chapkin, R.S. The role of docosahexaenoic acid in mediating mitochondrial membrane lipid oxidation and apoptosis in colonocytes. Carcinogenesis 2005, 26, 1914–1921. [Google Scholar] [CrossRef] [Green Version]
- Dommels, Y.E.; Haring, M.M.; Keestra, N.G.; Alink, G.M.; van Bladeren, P.J.; van Ommen, B. The role of cyclooxygenase in n-6 and n-3 polyunsaturated fatty acid mediated effects on cell proliferation, PGE synthesis and cytotoxicity in human colorectal carcinoma cell lines. Carcinogenesis 2003, 24, 385–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maziere, C.; Conte, M.A.; Degonville, J.; Ali, D.; Maziere, J.C. Cellular enrichment with polyunsaturated fatty acids induces an oxidative stress and activates the transcription factors AP1 and NFkappaB. Biochem. Biophys. Res. Commun. 1999, 265, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Bubici, C.; Papa, S.; Dean, K.; Franzoso, G. Mutual cross-talk between reactive oxygen species and nuclear factor-kappa B: Molecular basis and biological significance. Oncogene 2006, 25, 6731–6748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Hofmanova, J.; Vaculova, A.; Lojek, A.; Kozubık, A. Interaction of polyunsaturated fatty acids and sodium butyrate during apoptosis inHT-29 human colon adenocarcinoma cells. Eur. J. Nutr. 2005, 44, 40–51. [Google Scholar] [CrossRef]
- Sarsilmaz, M.; Songur, A.; Ozyurt, H.; Kus, I.; Ozen, O.A.; Ozyurt, B.; Söğüt, S.; Akyol, O. Potential role of dietary omega-3 essential fatty acids on some oxidant/antioxidant parameters in rats’ corpus striatum. Prostaglandins Leukot Essent Fatty Acids 2003, 69, 253–259. [Google Scholar] [CrossRef]
- Vartak, S.; Robbins, M.E.; Spector, A.A. Polyunsaturated fatty acids increase the sensitivity of 36B10 rat astrocytoma cells to radiation-induced cell kill. Lipids 1997, 32, 283–292. [Google Scholar] [CrossRef]
- Zand, H.; Rahimipour, A.; Salimi, S.; Shafiee, S.M. Docosahexaenoic acid sensitizes Ramos cells to gamma-irradiation-induced apoptosis through involvement of PPAR-gamma activation and NF-kappaB suppression. Mol. Cell. Biochem. 2008, 317, 113–120. [Google Scholar] [CrossRef]
- Colas, S.; Paon, L.; Denis, F.; Prat, M.; Louisot, P.; Hoinard, C.; le Floch, O.; Ogilvie, G.; Bougnoux, P. Enhanced radiosensitivity of rat autochthonous mammary tumors by dietary docosahexaenoic acid. Int. J. Cancer 2004, 109, 449–454. [Google Scholar] [CrossRef]
- Calviello, G.; Serini, S.; Piccioni, E.; Pessina, G. Antineoplastic effects of n-3 polyunsaturated fatty acids in combination with drugs and radiotherapy: Preventive and therapeutic strategies. Nutr. Cancer 2009, 61, 287–301. [Google Scholar] [CrossRef]
- Mane, S.D.; Kamatham, A.N. Ascorbyl stearate and ionizing radiation potentiate apoptosis through intracellular thiols and oxidative stress in murine T lymphoma cells. Chem. Biol. Interact. 2018, 281, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Uckun, F.M.; Dibirdik, I.; Qazi, S. Augmentation of the Antileukemia Potency of Total-Body Irradiation (TBI) by a Novel P-site Inhibitor of Spleen Tyrosine Kinase (SYK). Radiat. Res. 2010, 174, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Allegra, A.; Innao, V.; Allegra, A.G.; Pugliese, M.; di Salvo, E.; Ventura-Spagnolo, V.; Musolino, C.; Gangemi, S. Lymphocyte Subsets and Inflammatory Cytokines of Monoclonal Gammopathy of Undetermined Significance and Multiple Myeloma. Int. J. Mol. Sci. 2019, 20, 2822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musolino, C.; Allegra, A.; Innao, V.; Allegra, A.G.; Pioggia, G.; Gangemi, S. Inflammatory and Anti-Inflammatory Equilibrium, Proliferative and Antiproliferative Balance: The Role of Cytokines in Multiple Myeloma. Mediators Inflamm. 2017, 2017, 1852517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharazmi, A.; Nielsen, H.; Rechnitzer, C.; Bendtzen, K. Interleukin 6 primes human neutrophil and monocyte oxidative burst response. Immunol. Lett. 1989, 21, 177–184. [Google Scholar] [CrossRef]
- Behrens, M.M.; Ali, S.S.; Dugan, L.L. Interleukin-6 mediates the increase in NADPH-oxidase in the ketamine model of schizophrenia. J. Neurosci. 2008, 28, 13957–13966. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Klein, A.S.; Radaeva, S.; Hong, F.; El-Assal, O.; Pan, H.N.; Jaruga, B.; Batkai, S.; Hoshino, S.; Tian, Z.; et al. In vitro interleukin-6 treatment prevents mortality associated with fatty liver transplants in rats. Gastroenterology 2003, 125, 202–215. [Google Scholar] [CrossRef]
- Ward, N.S.; Waxman, A.B.; Homer, R.J.; Mantell, L.L.; Einarsson, O.; Du, Y.; Elias, J.A. Interleukin-6-induced protection in hyperoxic acute lung injury. Am. J. Respir. Cell. Mol. Biol. 2000, 22, 535–542. [Google Scholar] [CrossRef]
- Gangemi, S.; Allegra, A.; Alonci, A.; Cristani, M.; Russo, S.; Speciale, A.; Penna, G.; Spatari, G.; Cannavò, A.; Bellomo, G.; et al. Increase of novel biomarkers for oxidative stress in patients with plasma cell disorders and in multiple myeloma patients with bone lesions. Inflamm. Res. 2012, 61, 1063–1067. [Google Scholar] [CrossRef]
- Allegra, A.; Pace, E.; Tartarisco, G.; Innao, V.; DI Salvo, E.; Allegra, A.G.; Ferraro, M.; Musolino, C.; Gangemi, S. Changes in Serum Interleukin-8 and sRAGE Levels in Multiple Myeloma Patients. Anticancer Res. 2020, 40, 1443–1449. [Google Scholar] [CrossRef]
- Ettari, R.; Zappalà, M.; Grasso, S.; Musolino, C.; Innao, V.; Allegra, A. Immunoproteasome-selective and non-selective inhibitors: A promising approach for the treatment of multiple myeloma. Pharmacol. Ther. 2018, 182, 176–192. [Google Scholar] [CrossRef]
- Kuku, I.; Aydogdu, I.; Bayraktar, N.; Kaya, E.; Akyol, O.; Erkurt, M.A. Oxidant/antioxidant parameters and their relationship with medical treatment in multiple myeloma. Cell. Biochem. Funct. 2005, 23, 47–50. [Google Scholar] [CrossRef]
- Sharma, A.; Tripathi, M.; Satyam, A.; Kumar, L. Study of antioxidant levels in patients with multiple myeloma. Leuk. Lymphoma 2009, 50, 809–815. [Google Scholar] [CrossRef]
- Zima, T.; Spicka, I.; Stipek, S.; Crkovska, J.; Platenik, J.; Merta, M.; Nemecek, K.; Tesar, V. Lipid peroxidation and activity of antioxidative enzymes in patients with multiple myeloma. Cas. Lek. Cesk. 1996, 135, 14–17. [Google Scholar]
- Goel, A.; Spitz, D.R.; Weiner, G.J. Manipulation of cellular redox metabolism for improving therapeutic responses in B-cell lymphoma and multiple myeloma. J. Cell. Biochem. 2011, 113, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.O.; Salem, K.; Wagner, B.A.; Soumen Bera, S.; Singh, N.; Tiwari, A.; Choudhury, A.; Buettner, G.R.; Goel, A. Interleukin-6 counteracts therapy-induced cellular oxidative stress in multiple myeloma by up-regulating manganese superoxide dismutase. Biochem. J. 2012, 444, 515–527. [Google Scholar] [CrossRef] [Green Version]
- Gougelet, A.; Mansuy, A.; Blay, J.Y.; Alberti, L.; Vermot-Desroches, C. Lymphoma and myeloma cell resistance to cytotoxic agents and ionizing radiations is not affected by exposure to anti-IL-6 antibody. PLoS ONE 2009, 4, e8026. [Google Scholar] [CrossRef]
- Jaramillo, M.C.; Frye, J.B.; Crapo, J.D.; Briehl, M.M.; Tome, M.E. Increased manganese superoxide dismutase expression or treatment with manganese porphyrin potentiates dexamethasone-induced apoptosis in lymphoma cells. Cancer Res. 2009, 69, 5450–5457. [Google Scholar] [CrossRef] [Green Version]
- Yi, J.; Gao, F.; Shi, G.; Li, H.; Wang, Z.; Shi, X.; Tang, X. The inherent cellular level of reactive oxygen species: One of the mechanisms determining apoptotic susceptibility of leukemic cells to arsenic trioxide. Apoptosis 2002, 7, 209–215. [Google Scholar] [CrossRef]
- Bellosillo, B.; Villamor, N.; López-Guillermo, A.; Marcé, S.; Esteve, J.; Campo, E.; Colomer, D.; Montserrat, E. Complement-mediated cell death induced by rituximab in B-cell lymphoproliferative disorders is mediated in vitro by a caspase-independent mechanism involving the generation of reactive oxygen species. Blood 2001, 98, 2771–2777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Lim, M.-J.; Kim, M.-H.; Yu, C.H.; Yun, Y.S.; Ahn, J.; Song, J.Y. An effective strategy for increasing the radiosensitivity of Human lung Cancer cells by blocking Nrf2-dependent antioxidant responses. Free Radic. Biol. Med. 2012, 53, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Allegra, A.; Penna, G.; Alonci, A.; Rizzo, V.; Russo, S.; Musolino, C. Nanoparticles in oncology: The new theragnostic molecules. Anticancer Agents Med. Chem. 2011, 11, 669–686. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Liu, P.; He, N.; Gu, N.; Wu, F.G.; Chen, Z. Action of Gold Nanospikes-Based Nanoradiosensitizers: Cellular Internalization, Radiotherapy, and Autophagy. ACS Appl. Mater. Interfaces 2017, 9, 31526–31542. [Google Scholar] [CrossRef] [PubMed]
- Plunkett, W.; Gandhi, V.; Huang, P.; Robertson, L.E.; Yang, L.Y.; Gregoire, V.; Estey, E.; Keating, M.J. Fludarabine: Pharmacokinetics, mechanisms of action, and rationales for combination therapies. Semin. Oncol. 1993, 20, 2–12. [Google Scholar]
- Laurent, D.; Pradier, O.; Schmidberger, H.; Rave-Fränk, M.; Frankenberg, D.; Hess, C.F. Radiation rendered more cytotoxic by fludarabine monophosphate in a human oropharynx carcinoma cell-line than in fetal lung fibroblasts. J. Cancer Res. Clin. Oncol. 1998, 124, 485–492. [Google Scholar] [CrossRef]
- Donehower, R.C. An overview of the clinical experience with hydroxyurea. Semin. Oncol. 1992, 19, 11–19. [Google Scholar]
- Vokes, E.; Panje, W.; Schilsky, R.; Mick, R.; Awan, A.; Moran, W.; Goldman, M.; Tybor, A.; Weichselbaum, R. Hydroxyurea, fluorouracil, and concomitant radiotherapy in poor prognosis head and neck cancer: A phase I-II study. J. Clin. Oncol. 1989, 7, 761–768. [Google Scholar] [CrossRef]
- Allegra, A.; Musolino, C.; Tonacci, A.; Pioggia, G.; Gangemi, S. Interactions between the MicroRNAs and Microbiota in Cancer Development: Roles and Therapeutic opportunities. Cancers 2020, 12, 805. [Google Scholar] [CrossRef] [Green Version]
- Pajic, M.; Froio, D.; Daly, S.; Doculara, L.; Millar, E.; Graham, P.H.; Drury, A.; Steinmann, A.; de Bock, C.E.; Boulghourjian, A.; et al. miR-139-5p Modulates Radiotherapy Resistance in Breast Cancer by Repressing Multiple Gene Networks of DNA Repair and ROS Defense. Cancer Res. 2018, 78, 501–515. [Google Scholar] [CrossRef] [Green Version]
- Mao, Z.; Liu, S.; Cai, J.; Huang, Z.Z.; Lu, S.C. Cloning and functional characterization of the 5’-flanking region of human methionine adenosyltransferase 2A gene. Biochem. Biophys. Res. Commun. 1998, 248, 479–484. [Google Scholar] [CrossRef] [Green Version]
- Innao, V.; Allegra, A.; Pulvirenti, N.; Allegra, A.G.; Musolino, C. Therapeutic potential of antagomiRs in haematological and oncological neoplasms. Eur. J. Cancer Care 2020, 29, e13208. [Google Scholar] [CrossRef] [PubMed]
- Allegra, A.; Innao, V.; Basile, G.; Pugliese, M.; Allegra, A.G.; Pulvirenti, N.; Musolino, C. Post-chemotherapy cognitive impairment in hematological patients: Current understanding of chemobrain in hematology. Expert Rev. Hematol. 2020, 13, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Omuro, A.M.; Ben-Porat, L.S.; Panageas, K.S.; Kim, A.K.; Correa, D.D.; Yahalom, J.; Deangelis, L.M.; Abrey, L.E. Delayed neurotoxicity in primary central nervous system lymphoma. Arch. Neurol. 2005, 62, 1595–1600. [Google Scholar] [CrossRef] [Green Version]
- Weibrich, G.; Kleis, W.K.; Hafner, G.; Hitzler, W.E. Growth factor levels in platelet rich plasma and correlations with donor age, sex, and platelet count. J. Craniomaxillofac. Surg. 2002, 30, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Soliman, A.F.; Saif-Elnasr, M.; Abdel Fattah, S.M. Platelet-rich plasma ameliorates gamma radiation-induced nephrotoxicity via modulating oxidative stress and apoptosis. Life Sci. 2019, 219, 238–247. [Google Scholar] [CrossRef]
- Szumiel, I. From radioresistance to radiosensitivity: In vitro evolution of L5178Y lymphoma. Int. J. Radiat. Biol. 2015, 91, 465–471. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
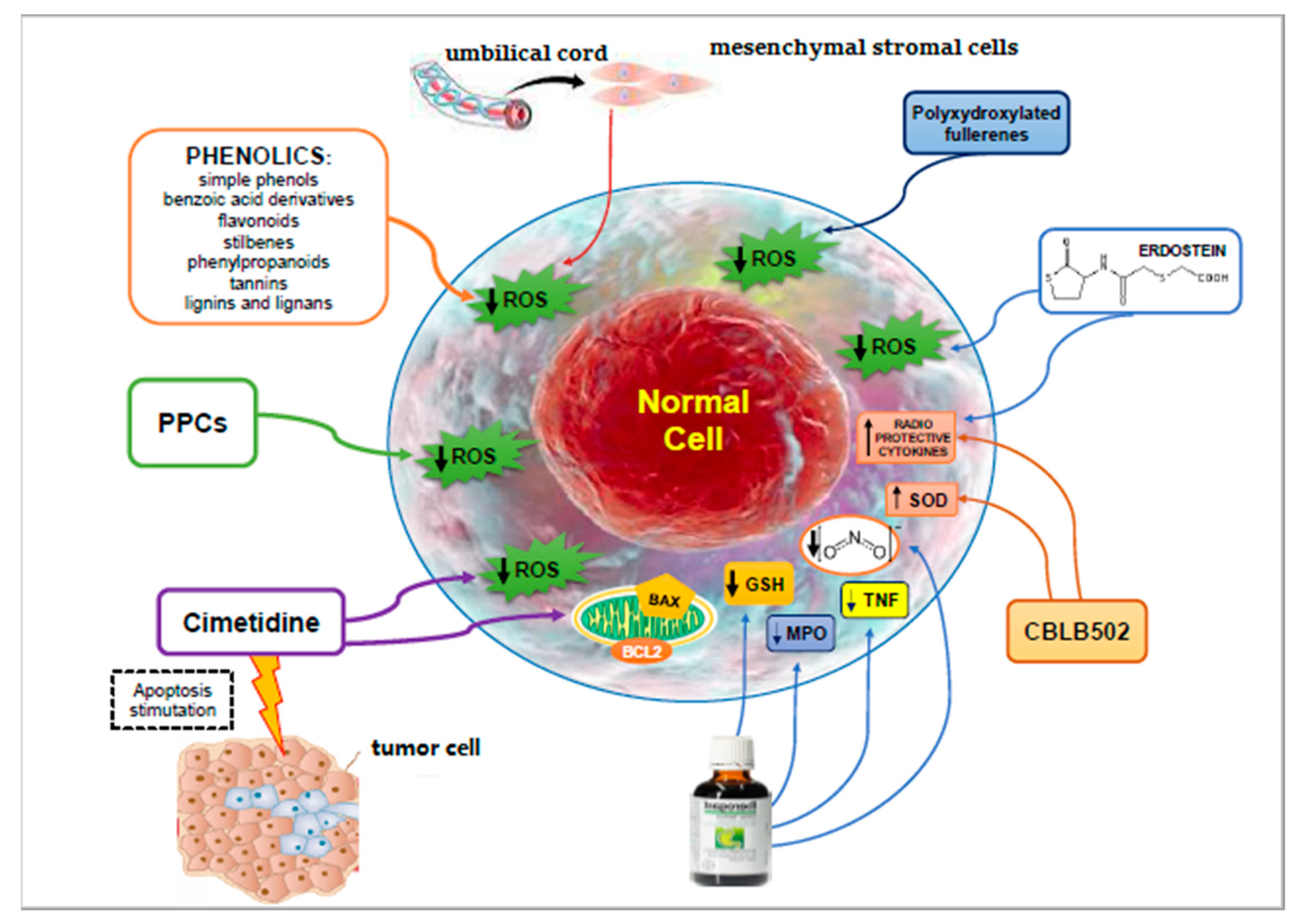
| Agent | Mechanisms of Action | Limits | Advantages | Ref. |
|---|---|---|---|---|
| Phenolic (flavonoids, stilbenes, tannins, lignans, lignins, quercetin, genistein) | Inhibition of apoptosis | Limited water solubility and poor availability | [49,50,51] | |
| Polyphenolic—polysaccharide conjugates | Inhibition of apoptosis | Non-toxic, water-soluble polymeric compounds. They did not protect the leukemic cells against radiation-induced apoptotic death. | [52,53] | |
| Cimetidine | Hydroxyl radical scavenging mechanism. Decrease of inflammation, and Bax/Bcl2 ratio. | Reduction of loss of bone marrow cell count, intestinal lining destruction, and fibrosis. Differential effect on both cancer cells and adjacent healthy cells. | [54,55] | |
| Amifostine (WR2721) | Organ repair via bone marrow recruitment or dedifferentiation. | Intolerance and significant accumulative toxicity. | [56] | |
| STW5 (Iberogast) | Anti-apoptotic effects. | Antioxidant activity and anti-inflammatory properties | Action on radiation enteritis. Preservation of the mucosal integrity of the small intestine. | [57,58,59,60] |
| CBLB502 | Increased expression of the strong natural antioxidant superoxide dismutase and induction of radioprotective cytokines (G-CSF, IL-6, and TNF-α). Suppression of p53-dependent apoptosis. Reduction of DNA damage and chromosomal aberrations. Action on the TLR5 signaling pathway. | It protects mammals from gastrointestinal and hematopoietic acute radiation syndrome. Reduction of IR-induced oxidative stress, reduction of decline of sperm quantity and quality. | [61,62,63,64,65] [66,67] | |
| Erdosteine | Protective role on the release of free oxygen radicals. Action on TNF alpha, interleukin 1, and IL-6. | Protection against radiation induced inflammatory kidney damage. | [68] | |
| Human umbilical cord-derived mesenchymal stromal cells | Prevention of oxidative stress and increased antioxidant status. Reduction of pro-fibrotic TGF-β1, IL-6, and IL-8 levels. | Protective effects on irradiation myocardial fibrosis with increased cell viability, reduction of collagen deposition. | [69] | |
| Calf spleen extractive injection | Regenerating action on damaged cells. | Reduction of thrombocytopenia and leucopenia. | [70,71,72] | |
| Polyxydroxylated fullerenes | Anti-oxidative effects. | Prevention of radiation-induced reduction in the white cell count. | [73,74] | |
| Platelet-rich plasma | Administration of growth factors. Reduction of oxidative stress and inhibition of the induced apoptosis. | Neuroprotection. | [75,76] |
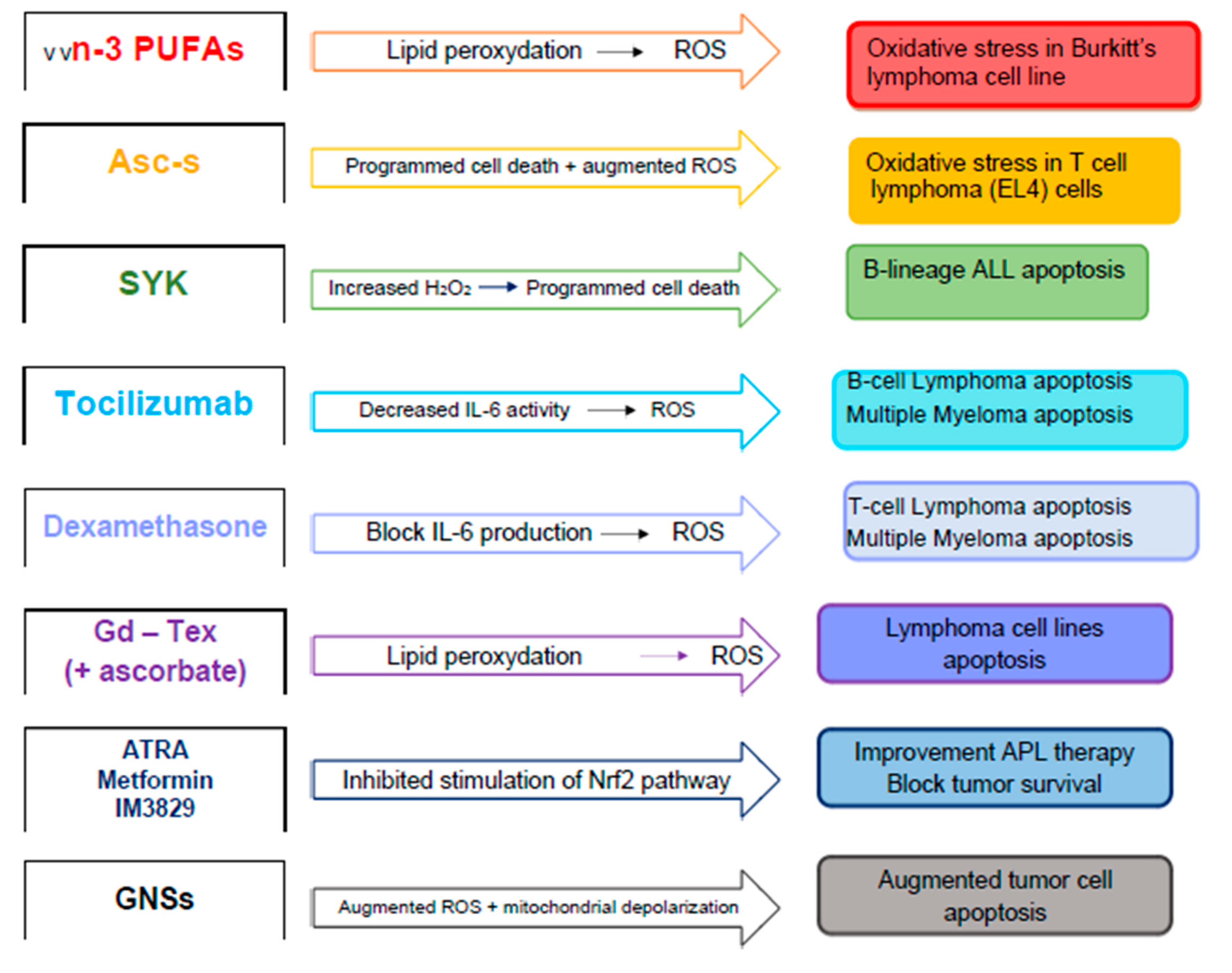
| Agent | Mechanisms of Action | Effects | Ref. |
|---|---|---|---|
| Natural phytochemicals (curcumin, demethoxycurcumin, quercetin, genistein) | Alteration of levels of radioprotective metabolites. Electron transfer to a radiation sensitizer. | Reduction of radio-resistance. | [100,101,102] |
| Long-chain n-3 polyunsaturated fatty acids | Their peroxidation may sensitize cells to ROS, inducing an oxidative stress. Modulation of ROS-sensitive mitogen-activated protein kinases and phosphatases, and transcription factors. | Cytotoxicity. Increased radiation-induced apoptosis. | [103,104] |
| Ascorbyl stearate | Augmented levels of ROS, drop in mitochondrial membrane potential and increased caspase-3 activity. | Reduction of cell proliferation, induction of apoptosis by arresting the cells at S/G2-M phase of cell cycle. | [105] |
| Spleen tyrosine kinase (SYK) P-site inhibitor | Increased H2O2-induced apoptosis. | Action on radio-chemotherapy resistance. | [106] |
| Dexamethasone | Increased superoxide and hydrogen peroxide production and augmented radiation-induced oxidative stress. | Clonogenic cell killing and apoptosis of myeloma cells. | [107] |
| Rituximab | Elevation in ROS generation | Increase of cell growth inhibition. Augmented apoptosis. | [108,109,110,111,112,113] |
| All-trans-retinoic acid, Metformin, IM3829 | Inhibition of nuclear factor erythroid 2-related factor 2 | Inhibition of cancer cell survival. | [114,115,116] |
| Gold nanoparticle-based compounds | Increased ROS levels, mitochondrial depolarization, and cell cycle redistribution. | Inhibition of protective autophagy. | [117,118] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allegra, A.G.; Mannino, F.; Innao, V.; Musolino, C.; Allegra, A. Radioprotective Agents and Enhancers Factors. Preventive and Therapeutic Strategies for Oxidative Induced Radiotherapy Damages in Hematological Malignancies. Antioxidants 2020, 9, 1116. https://doi.org/10.3390/antiox9111116
Allegra AG, Mannino F, Innao V, Musolino C, Allegra A. Radioprotective Agents and Enhancers Factors. Preventive and Therapeutic Strategies for Oxidative Induced Radiotherapy Damages in Hematological Malignancies. Antioxidants. 2020; 9(11):1116. https://doi.org/10.3390/antiox9111116
Chicago/Turabian StyleAllegra, Andrea Gaetano, Federica Mannino, Vanessa Innao, Caterina Musolino, and Alessandro Allegra. 2020. "Radioprotective Agents and Enhancers Factors. Preventive and Therapeutic Strategies for Oxidative Induced Radiotherapy Damages in Hematological Malignancies" Antioxidants 9, no. 11: 1116. https://doi.org/10.3390/antiox9111116
APA StyleAllegra, A. G., Mannino, F., Innao, V., Musolino, C., & Allegra, A. (2020). Radioprotective Agents and Enhancers Factors. Preventive and Therapeutic Strategies for Oxidative Induced Radiotherapy Damages in Hematological Malignancies. Antioxidants, 9(11), 1116. https://doi.org/10.3390/antiox9111116