Miracle Berry as a Potential Supplement in the Control of Metabolic Risk Factors in Cancer

 , ,
, ,
Abstract
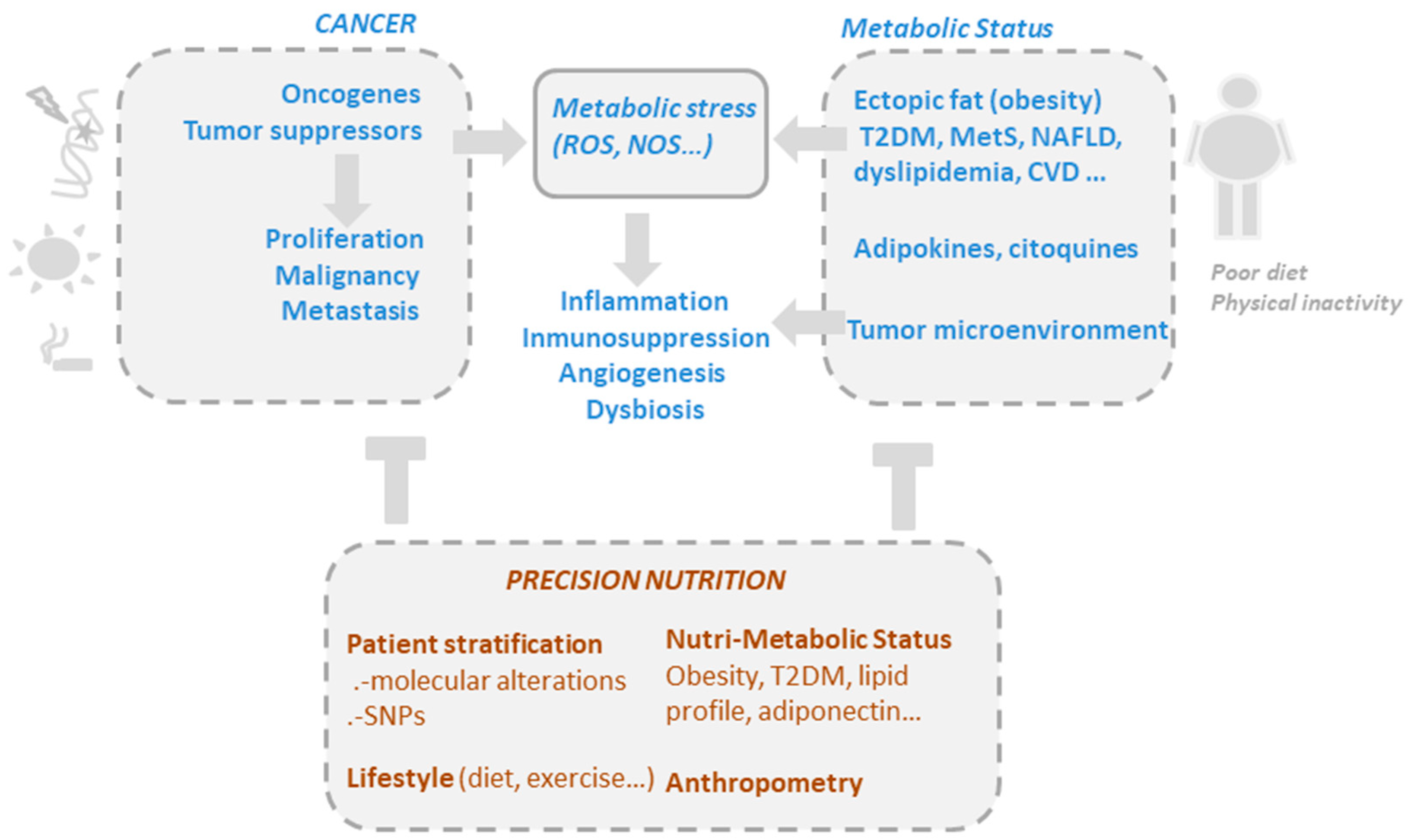
:1. Cancer as a Metabolic Disease
1.1. Obesity Is Associated with a Low Grade of Chronic Inflammation and Cancer Risk
1.2. Dual Role of Phytochemicals Modulating Oxidative Stress in Cancer
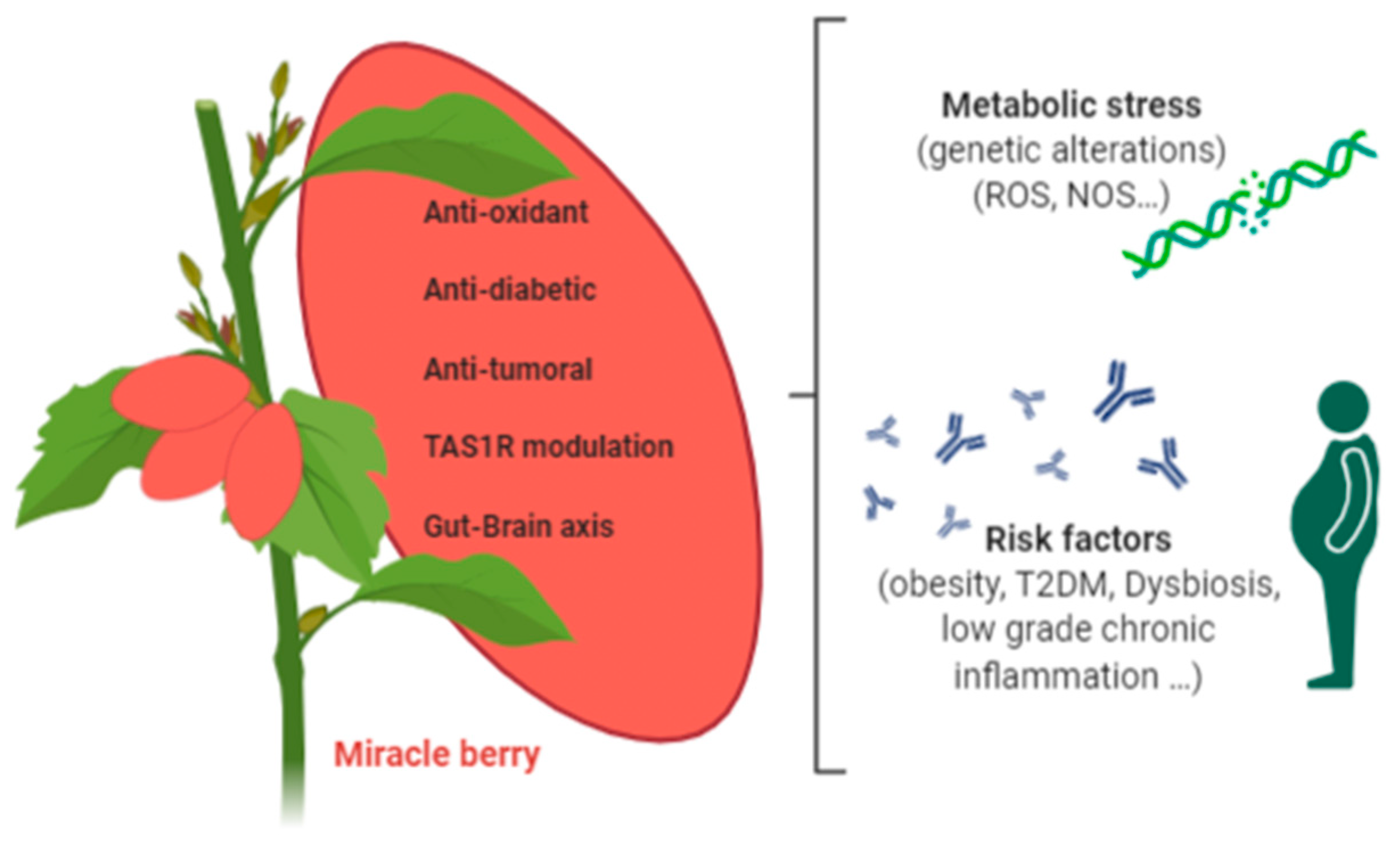
2. Miracle Berry
2.1. Antioxidant Activities of Miracle Berry Extracts
2.2. Miracle Berry Targeting Metabolic Risk Factors Associated with Cancer
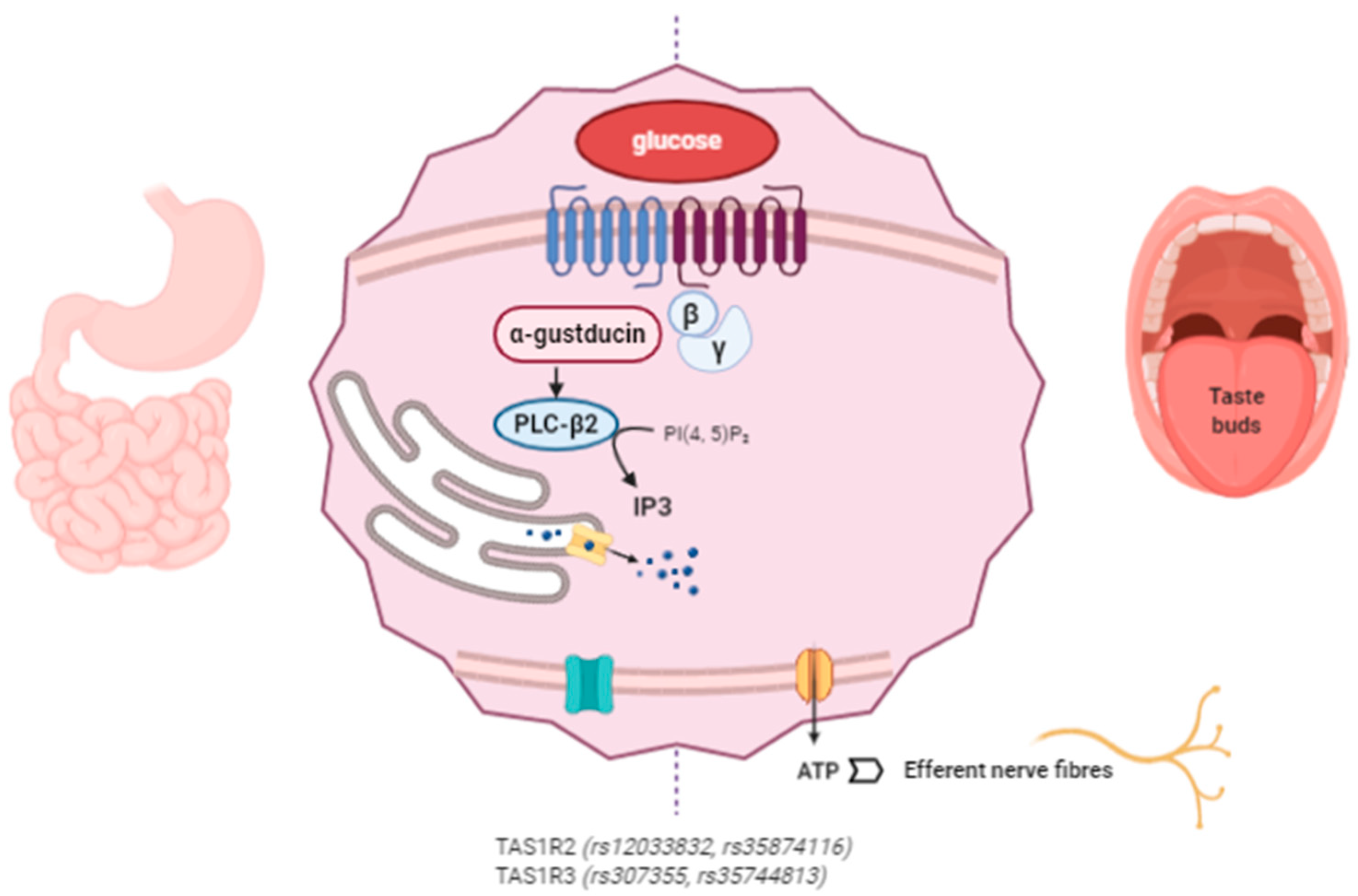
3. Miraculin
3.1. Mechanism of Action of Miraculin
3.2. Clinical Applications of Miraculin
3.3. Miraculin Modulating the Gut–Brain Axis
3.4. Nutrigenetics: Genetic Variability in the Perception of Sweet-Taste Affects Behavior and Nutrient Intake
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABTS | 2,20-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) |
| AICR | American Institute for Cancer Research |
| BC | Breast cancer |
| BMI | Body mass index |
| CVD | Cardiovascular disease |
| DPPH | (1,1-Diphenyl-2-picrylhydrazyl) |
| FRAP | Ferric reducing antioxidant power |
| HOMA-IR | Homeostatic model assessment for insulin resistance |
| hsCRP | High sensitivity C-reactive protein |
| MB | Miracle berry |
| MetS | Metabolic syndrome |
| mCRC | Metastatic colorectal cancer |
| NAFLD | Non-alcoholic fatty liver disease |
| NSCLC | Non-small cell lung cancer |
| ROS | Reactive oxygen species |
| T2DM | Type 2 diabetes mellitus |
| SNPs | Single nucleotide polymorphisms |
| WCRF | World Cancer Research Foundation |
References
- Arnold, M.; Leitzmann, M.; Freisling, H.; Bray, F.; Romieu, I.; Renehan, A.; Soerjomataram, I. Obesity and cancer: An update of the global impact. Cancer Epidemiol. 2016, 41, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, J.; Farr, O.; Perakakis, N.; Ghaly, W.; Mantzoros, C. Obesity as a Disease. Med. Clin. N. Am. 2018, 102, 13–33. [Google Scholar] [CrossRef]
- Bluher, M. Metabolically Healthy Obesity. Endocr. Rev. 2020, 41, bnaa004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyrgiou, M.; Kalliala, I.; Markozannes, G.; Gunter, M.J.; Paraskevaidis, E.; Gabra, H.; Martin-Hirsch, P.; Tsilidis, K.K. Adiposity and cancer at major anatomical sites: Umbrella review of the literature. BMJ 2017, 356, j477. [Google Scholar] [CrossRef] [Green Version]
- Trestini, I.; Carbognin, L.; Bonaiuto, C.; Tortora, G.; Bria, E. The obesity paradox in cancer: Clinical insights and perspectives. Eat. Weight Disord. 2018, 23, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Avgerinos, K.I.; Spyrou, N.; Mantzoros, C.S.; Dalamaga, M. Obesity and cancer risk: Emerging biological mechanisms and perspectives. Metabolism 2019, 92, 121–135. [Google Scholar] [CrossRef]
- Deng, T.; Lyon, C.J.; Bergin, S.; Caligiuri, M.A.; Hsueh, W.A. Obesity, Inflammation, and Cancer. Annu. Rev. Pathol. 2016, 11, 421–449. [Google Scholar] [CrossRef] [Green Version]
- Carbone, F.; la Rocca, C.; Matarese, G. Immunological functions of leptin and adiponectin. Biochimie 2012, 94, 2082–2088. [Google Scholar] [CrossRef]
- Lee, D.Y.; Rhee, E.J.; Chang, Y.; Sohn, C.I.; Shin, H.C.; Ryu, S.; Lee, W.Y. Impact of systemic inflammation on the relationship between insulin resistance and all-cause and cancer-related mortality. Metabolism 2018, 81, 52–62. [Google Scholar] [CrossRef]
- Sabharwal, S.S.; Schumacker, P.T. Mitochondrial ROS in cancer: Initiators, amplifiers or an Achilles’ heel? Nat. Rev. Cancer 2014, 14, 709–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.J.; Murphy, K.E.; Fernandez, M.L. Impact of Obesity and Metabolic Syndrome on Immunity. Adv. Nutr. 2016, 7, 66–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Cani, P.D. Diabetes obesity and gut microbiota. Best Pract. Res. Clin. Gastroenterol. 2013, 27, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Pisanu, S.; Palmas, V.; Madau, V.; Casula, E.; Deledda, A.; Cusano, R.; Uva, P.; Vascellari, S.; Boi, F.; Loviselli, A.; et al. Impact of a Moderately Hypocaloric Mediterranean Diet on the Gut Microbiota Composition of Italian Obese Patients. Nutrients 2020, 12, 2707. [Google Scholar] [CrossRef]
- Ortega, M.A.; Fraile-Martinez, O.; Naya, I.; Garcia-Honduvilla, N.; Alvarez-Mon, M.; Bujan, J.; Asunsolo, A.; de la Torre, B. Type 2 Diabetes Mellitus Associated with Obesity (Diabesity). The Central Role of Gut Microbiota and Its Translational Applications. Nutrients 2020, 12, 2749. [Google Scholar] [CrossRef]
- Yoo, J.Y.; Groer, M.; Dutra, S.V.O.; Sarkar, A.; McSkimming, D.I. Gut Microbiota and Immune System Interactions. Microorganisms 2020, 8, 1587. [Google Scholar] [CrossRef]
- Jie, Z.; Xia, H.; Zhong, S.L.; Feng, Q.; Li, S.; Liang, S.; Zhong, H.; Liu, Z.; Gao, Y.; Zhao, H.; et al. The gut microbiome in atherosclerotic cardiovascular disease. Nat. Commun. 2017, 8, 845. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 308–325. [Google Scholar] [CrossRef] [Green Version]
- Nagpal, R.; Newman, T.M.; Wang, S.; Jain, S.; Lovato, J.F.; Yadav, H. Obesity-Linked Gut Microbiome Dysbiosis Associated with Derangements in Gut Permeability and Intestinal Cellular Homeostasis Independent of Diet. J. Diabetes Res. 2018, 2018, 3462092. [Google Scholar] [CrossRef]
- Kim, B.; Choi, H.N.; Yim, J.E. Effect of Diet on the Gut Microbiota Associated with Obesity. J. Obes. Metab. Syndr. 2019, 28, 216–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damms-Machado, A.; Mitra, S.; Schollenberger, A.E.; Kramer, K.M.; Meile, T.; Konigsrainer, A.; Huson, D.H.; Bischoff, S.C. Effects of surgical and dietary weight loss therapy for obesity on gut microbiota composition and nutrient absorption. Biomed. Res. Int. 2015, 2015, 806248. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Ma, L.; Ma, Y.; Zhang, F.; Zhao, C.; Nie, Y. Insights into the role of gut microbiota in obesity: Pathogenesis, mechanisms, and therapeutic perspectives. Protein. Cell 2018, 9, 397–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sena, L.A.; Chandel, N.S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell 2012, 48, 158–167. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of Reactive Oxygen Species in Cancer Progression: Molecular Mechanisms and Recent Advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NavaneethaKrishnan, S.; Rosales, J.L.; Lee, K.Y. ROS-Mediated Cancer Cell Killing through Dietary Phytochemicals. Oxid. Med. Cell Longev. 2019, 2019, 9051542. [Google Scholar] [CrossRef]
- Schumacker, P.T. Reactive oxygen species in cancer cells: Live by the sword, die by the sword. Cancer Cell 2006, 10, 175–176. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Huang, J.; Zuo, Y.; Li, B.; Guo, Q.; Cui, B.; Shao, W.; Du, J.; Bu, X. 2a, a novel curcumin analog, sensitizes cisplatin-resistant A549 cells to cisplatin by inhibiting thioredoxin reductase concomitant oxidative stress damage. Eur. J. Pharmacol. 2013, 707, 130–139. [Google Scholar] [CrossRef]
- Li, Q.; Zhan, M.; Chen, W.; Zhao, B.; Yang, K.; Yang, J.; Yi, J.; Huang, Q.; Mohan, M.; Hou, Z.; et al. Phenylethyl isothiocyanate reverses cisplatin resistance in biliary tract cancer cells via glutathionylation-dependent degradation of Mcl-1. Oncotarget 2016, 7, 10271–10282. [Google Scholar] [CrossRef] [Green Version]
- Chikara, S.; Nagaprashantha, L.D.; Singhal, J.; Horne, D.; Awasthi, S.; Singhal, S.S. Oxidative stress and dietary phytochemicals: Role in cancer chemoprevention and treatment. Cancer Lett. 2018, 413, 122–134. [Google Scholar] [CrossRef]
- Chirumbolo, S.; Bjorklund, G.; Lysiuk, R.; Vella, A.; Lenchyk, L.; Upyr, T. Targeting Cancer with Phytochemicals via Their Fine Tuning of the Cell Survival Signaling Pathways. Int. J. Mol. Sci. 2018, 19, 3568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Chen, Y.; St Clair, D.K. ROS and p53: A versatile partnership. Free Radic. Biol. Med. 2008, 44, 1529–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckmire, R.E.; Francis, F.J. Anthocyanins and Flavonols of miracle fruit, Synsepalum dulcificum, Schum. J. Food Sci. 1976, 41, 1363–1365. [Google Scholar] [CrossRef]
- Ntie-Kang, F.; Lifongo, L.L.; Mbaze, L.M.; Ekwelle, N.; Owono Owono, L.C.; Megnassan, E.; Judson, P.N.; Sippl, W.; Efange, S.M. Cameroonian medicinal plants: A bioactivity versus ethnobotanical survey and chemotaxonomic classification. BMC Complement. Altern. Med. 2013, 13, 147. [Google Scholar] [CrossRef] [Green Version]
- Inglett, G.E.; Chen, D. Contents of phenolics and flavonoids and antioxidant activities in skin, pulp, and seeds of miracle fruit. J. Food Sci. 2011, 76, C479–C482. [Google Scholar] [CrossRef]
- Wang, H.M.; Chou, Y.T.; Hong, Z.L.; Chen, H.A.; Chang, Y.C.; Yang, W.L.; Chen, C.Y.; Mai, C.T.; Chen, C.Y. Bioconstituents from stems of Synsepalum dulcificum Daniell (Sapotaceae) inhibit human melanoma proliferation, reduce mushroom tyrosinase activity and have antioxidant properties. J. Taiwan Inst. Chem. Eng. 2011, 42, 204–211. [Google Scholar] [CrossRef]
- Brouwer, J.N.; Glaser, D.; Hard Af Segerstad, C.; Hellekant, G.; Ninomiya, Y.; Van der Wel, H. The sweetness-inducing effect of miraculin; behavioural and neurophysiological experiments in the rhesus monkey Macaca mulatta. J. Physiol. 1983, 337, 221–240. [Google Scholar] [CrossRef]
- Bartoshuk, L.M.; Gentile, R.L.; Moskowitz, H.R.; Meiselman, H.L. Sweet taste induced by miracle fruit (Synsepalum dulcificum). Physiol. Behav. 1974, 12, 449–456. [Google Scholar] [CrossRef]
- Theerasilp, S.; Hitotsuya, H.; Nakajo, S.; Nakaya, K.; Nakamura, Y.; Kurihara, Y. Complete amino acid sequence and structure characterization of the taste-modifying protein, miraculin. J. Biol. Chem. 1989, 264, 6655–6659. [Google Scholar]
- Wong, J.M.; Kern, M. Miracle fruit improves sweetness of a low-calorie dessert without promoting subsequent energy compensation. Appetite 2011, 56, 163–166. [Google Scholar] [CrossRef]
- Chen, C.C.; Liu, I.M.; Cheng, J.T. Improvement of insulin resistance by miracle fruit (Synsepalum dulcificum) in fructose-rich chow-fed rats. Phytother. Res. 2006, 20, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Shen, Y.; Zhang, X.; Prinyawiwatkul, W.; Xu, Z. Antioxidant-rich phytochemicals in miracle berry (Synsepalum dulcificum) and antioxidant activity of its extracts. Food Chem. 2014, 153, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.C.; Wu, J.Y.; Wang, C.K. Modulatory effects of miracle fruit ethanolic extracts on glucose uptake through the insulin signaling pathway in C2C12 mouse myotubes cells. Food Sci. Nutr. 2019, 7, 1035–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.Y.; Wu, P.Y.; Huang, T.S.; Lin, C.W.; Li, Y.C.; Chou, R.H.; Wang, H.M.; Chang, H.W. The Sour Taste-Modifying Protein (Miraculin), Tyrosinase Inhibitors and Antioxidants from Synsepalum dulcificum. Curr. Nutr. Food Sci. 2009, 5, 172–179. [Google Scholar] [CrossRef]
- Obafemi, T.O.; Akinmoladun, A.C.; Olaleye, M.T.; Agboade, S.O.; Onasanya, A.A. Antidiabetic potential of methanolic and flavonoid-rich leaf extracts of Synsepalum dulcificum in type 2 diabetic rats. J. Ayurveda Integr. Med. 2017, 8, 238–246. [Google Scholar] [CrossRef]
- Chen, T.Y.; Kang, Z.C.; Yen, M.T.; Huang, M.H.; Wang, B.S. Inhibitory effect of aqueous extracts from Miracle Fruit leaves on mutation and oxidative damage. Food Chem. 2015, 169, 411–416. [Google Scholar] [CrossRef]
- Salehi, B.; Ata, A.; V Anil Kumar, N.; Sharopov, F.; Ramirez-Alarcon, K.; Ruiz-Ortega, A.; Abdulmajid Ayatollahi, S.; Tsouh Fokou, P.V.; Kobarfard, F.; Amiruddin Zakaria, Z.; et al. Antidiabetic Potential of Medicinal Plants and Their Active Components. Biomolecules 2019, 9, 551. [Google Scholar] [CrossRef] [Green Version]
- Obafemi, T.O.; Olaleye, M.T.; Akinmoladun, A.C. Antidiabetic property of miracle fruit plant (Synsepalum dulcificum Shumach. & Thonn. Daniell) leaf extracts in fructose-fed streptozotocin-injected rats via anti-inflammatory activity and inhibition of carbohydrate metabolizing enzymes. J. Ethnopharmacol. 2019, 244, 112124. [Google Scholar]
- Huang, W.; Chung, H.Y.; Xuan, W.; Wang, G.; Li, Y. The cholesterol-lowering activity of miracle fruit (Synsepalum dulcificum). J. Food Biochem. 2020, 44, e13185. [Google Scholar] [CrossRef]
- Shi, Y.C.; Lin, K.S.; Jhai, Y.F.; Lee, B.H.; Han, Y.; Cui, Z.; Hsu, W.H.; Wu, S.C. Miracle Fruit (Synsepalum dulcificum) Exhibits as a Novel Anti-Hyperuricaemia Agent. Molecules 2016, 21, 140. [Google Scholar] [CrossRef] [Green Version]
- Seong, J.; Oyong, G.; Cabrera, E. Synsepalum dulcificum extracts exhibit cytotoxic activity on human colorectal cancer cells and upregulate c-fos and c-jun early apoptotic gene expression. Asian Pac. J. Trop. Biomed. 2018, 8, 173–178. [Google Scholar]
- Kant, R. Sweet proteins--potential replacement for artificial low calorie sweeteners. Nutr. J. 2005, 4, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurihara, K.; Beidler, L.M. Taste-modifying protein from miracle fruit. Science 1968, 161, 1241–1243. [Google Scholar] [CrossRef] [PubMed]
- Wilken, M.K.; Satiroff, B.A. Pilot study of “miracle fruit” to improve food palatability for patients receiving chemotherapy. Clin. J. Oncol. Nurs. 2012, 16, E173–E177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, H.P.; Cusnir, M.; Schwartz, M.A.; Pizzolato, J.F.; Lutzky, J.; Campbell, R.J.; Beaumont, J.L.; Eton, D.; Stonick, S.; Lilenbaum, R. Treatment of taste alterations in chemotherapy patients using the “miracle fruit”: Preliminary analysis of a pilot study. J. Clin. Oncol. 2010, 28 (Suppl. 15), e19523. [Google Scholar] [CrossRef]
- Hiwasa-Tanase, K.; Hirai, T.; Kato, K.; Duhita, N.; Ezura, H. From miracle fruit to transgenic tomato: Mass production of the taste-modifying protein miraculin in transgenic plants. Plant Cell Rep. 2012, 31, 513–525. [Google Scholar] [CrossRef]
- Temussi, P.A. Natural sweet macromolecules: How sweet proteins work. Cell Mol. Life Sci. 2006, 63, 1876–1888. [Google Scholar] [CrossRef]
- Kurihara, Y. Characteristics of antisweet substances, sweet proteins, and sweetness-inducing proteins. Crit. Rev. Food Sci. Nutr. 1992, 32, 231–252. [Google Scholar] [CrossRef]
- Misaka, T. Molecular mechanisms of the action of miraculin, a taste-modifying protein. Semin Cell Dev. Biol. 2013, 24, 222–225. [Google Scholar] [CrossRef]
- Picone, D.; Temussi, P.A. Dissimilar sweet proteins from plants: Oddities or normal components? Plant Sci. 2012, 195, 135–142. [Google Scholar] [CrossRef]
- Koizumi, A.; Tsuchiya, A.; Nakajima, K.; Ito, K.; Terada, T.; Shimizu-Ibuka, A.; Briand, L.; Asakura, T.; Misaka, T.; Abe, K. Human sweet taste receptor mediates acid-induced sweetness of miraculin. Proc. Natl. Acad. Sci. USA 2011, 108, 16819–16824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.E.; Garza, J. Effects of different miracle fruit products on the sensory characteristics of different types of sour foods by descriptive analysis. J. Food Sci. 2020, 85, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.F.; Andrade, R.D.S.; Bastos, S.C.; Coelho, S.B.; Pinheiro, A.C.M. Miracle fruit: An alternative sugar substitute in sour beverages. Appetite 2016, 107, 645–653. [Google Scholar] [CrossRef] [PubMed]
- van der Wel, H.; Loeve, K. Isolation and characterization of thaumatin I and II, the sweet-tasting proteins from Thaumatococcus daniellii Benth. Eur. J. Biochem. 1972, 31, 221–225. [Google Scholar]
- Yamashita, H.; Theerasilp, S.; Aiuchi, T.; Nakaya, K.; Nakamura, Y.; Kurihara, Y. Purification and complete amino acid sequence of a new type of sweet protein taste-modifying activity, curculin. J. Biol. Chem. 1990, 265, 15770–15775. [Google Scholar] [PubMed]
- Ming, D.; Hellekant, G. Brazzein, a new high-potency thermostable sweet protein from Pentadiplandra brazzeana B. FEBS Lett. 1994, 355, 106–108. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Maeda, S.; Hu, Z.; Aiuchi, T.; Nakaya, K.; Kurihara, Y. Purification, complete amino acid sequence and structural characterization of the heat-stable sweet protein, mabinlin II. Eur. J. Biochem. 1993, 211, 281–287. [Google Scholar] [CrossRef]
- Brouwer, J.N.; van der Wel, H.; Francke, A.; Henning, G.J. Mieraculin, the sweetness-inducing protein from miracle fruit. Nature 1968, 220, 373–374. [Google Scholar] [CrossRef]
- Sun, H.J.; Cui, M.L.; Ma, B.; Ezura, H. Functional expression of the taste-modifying protein, miraculin, in transgenic lettuce. FEBS Lett. 2006, 580, 620–626. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.J.; Kataoka, H.; Yano, M.; Ezura, H. Genetically stable expression of functional miraculin, a new type of alternative sweetener, in transgenic tomato plants. Plant Biotechnol. J. 2007, 5, 768–777. [Google Scholar] [CrossRef]
- Kim, S.K.; Chen, Y.; Abrol, R.; Goddard, W.A., 3rd; Guthrie, B. Activation mechanism of the G protein-coupled sweet receptor heterodimer with sweeteners and allosteric agonists. Proc. Natl. Acad. Sci. USA 2017, 114, 2568–2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Staszewski, L.; Xu, H.; Durick, K.; Zoller, M.; Adler, E. Human receptors for sweet and umami taste. Proc. Natl. Acad. Sci. USA 2002, 99, 4692–4696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashani-Amin, E.; Sakhteman, A.; Larijani, B.; Ebrahim-Habibi, A. Introducing a New Model of Sweet Taste Receptor, a Class C G-protein Coupled Receptor (C GPCR). Cell Biochem. Biophys. 2019, 77, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Sanematsu, K.; Kitagawa, M.; Yoshida, R.; Nirasawa, S.; Shigemura, N.; Ninomiya, Y. Intracellular acidification is required for full activation of the sweet taste receptor by miraculin. Sci. Rep. 2016, 6, 22807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Hoon, M.A.; Chandrashekar, J.; Mueller, K.L.; Cook, B.; Wu, D.; Zuker, C.S.; Ryba, N.J. Coding of sweet, bitter, and umami tastes: Different receptor cells sharing similar signaling pathways. Cell 2003, 112, 293–301. [Google Scholar] [CrossRef] [Green Version]
- Rossler, P.; Kroner, C.; Freitag, J.; Noe, J.; Breer, H. Identification of a phospholipase C beta subtype in rat taste cells. Eur. J. Cell Biol. 1998, 77, 253–261. [Google Scholar] [CrossRef]
- Perez, C.A.; Huang, L.; Rong, M.; Kozak, J.A.; Preuss, A.K.; Zhang, H.; Max, M.; Margolskee, R.F. A transient receptor potential channel expressed in taste receptor cells. Nat. Neurosci. 2002, 5, 1169–1176. [Google Scholar] [CrossRef]
- Pineli, L.L.O.; Aguiar, L.A.; Fiusa, A.; Botelho, R.B.A.; Zandonadi, R.P.; Melo, L. Sensory impact of lowering sugar content in orange nectars to design healthier, low-sugar industrialized beverages. Appetite 2016, 96, 239–244. [Google Scholar] [CrossRef]
- Malik, V.S.; Hu, F.B. Fructose and Cardiometabolic Health: What the Evidence from Sugar-Sweetened Beverages Tells Us. J. Am. Coll. Cardiol. 2015, 66, 1615–1624. [Google Scholar] [CrossRef] [Green Version]
- Malik, V.S.; Pan, A.; Willett, W.C.; Hu, F.B. Sugar-sweetened beverages and weight gain in children and adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2013, 98, 1084–1102. [Google Scholar] [CrossRef] [Green Version]
- Pepino, M.Y. Metabolic effects of non-nutritive sweeteners. Physiol. Behav. 2015, 152, 450–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, M.V.; Small, D.M. Physiological mechanisms by which non-nutritive sweeteners may impact body weight and metabolism. Physiol. Behav. 2015, 152, 381–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benton, D. Can artificial sweeteners help control body weight and prevent obesity? Nutr. Res. Rev. 2005, 18, 63–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raben, A.; Vasilaras, T.H.; Moller, A.C.; Astrup, A. Sucrose compared with artificial sweeteners: Different effects on ad libitum food intake and body weight after 10 wk of supplementation in overweight subjects. Am. J. Clin. Nutr. 2002, 76, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Sarnelli, G.; Annunziata, G.; Magno, S.; Oriolo, C.; Savastano, S.; Colao, A. Taste and the Gastrointestinal tract: From physiology to potential therapeutic target for obesity. Int. J. Obes. Suppl. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.T.; Gannon, K.S.; Margolskee, R.F. Transduction of bitter and sweet taste by gustducin. Nature 1996, 381, 796–800. [Google Scholar] [CrossRef]
- Halyard, M.Y. Taste and smell alterations in cancer patients—Real problems with few solutions. J. Support. Oncol. 2009, 7, 68–69. [Google Scholar]
- Sherry, V.W. Taste alterations among patients with cancer. Clin. J. Oncol. Nurs. 2002, 6, 73–77. [Google Scholar] [CrossRef] [Green Version]
- Berteretche, M.V.; Dalix, A.M.; d’Ornano, A.M.; Bellisle, F.; Khayat, D.; Faurion, A. Decreased taste sensitivity in cancer patients under chemotherapy. Support. Care Cancer 2004, 12, 571–576. [Google Scholar] [CrossRef]
- Ravasco, P.; Monteiro-Grillo, I.; Camilo, M. Individualized nutrition intervention is of major benefit to colorectal cancer patients: Long-term follow-up of a randomized controlled trial of nutritional therapy. Am. J. Clin. Nutr. 2012, 96, 1346–1353. [Google Scholar] [CrossRef] [Green Version]
- Laffitte, A.; Neiers, F.; Briand, L. Functional roles of the sweet taste receptor in oral and extraoral tissues. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Folgueira, C.; Seoane, L.M.; Casanueva, F.F. The brain-stomach connection. Front. Horm. Res. 2014, 42, 83–92. [Google Scholar] [PubMed]
- Stimac, D.; Majanovic, S.K.; Franjic, N. Stomach--key player in the regulation of metabolism. Dig. Dis. 2014, 32, 192–201. [Google Scholar] [PubMed]
- Depoortere, I. Taste receptors of the gut: Emerging roles in health and disease. Gut 2014, 63, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Shirazi-Beechey, S.P.; Daly, K.; Al-Rammahi, M.; Moran, A.W.; Bravo, D. Role of nutrient-sensing taste 1 receptor (T1R) family members in gastrointestinal chemosensing. Br. J. Nutr. 2014, 111 (Suppl. 1), S8–S15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baggio, L.L.; Drucker, D.J. Biology of incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef] [PubMed]
- Widmayer, P.; Kuper, M.; Kramer, M.; Konigsrainer, A.; Breer, H. Altered expression of gustatory-signaling elements in gastric tissue of morbidly obese patients. Int. J. Obes. 2012, 36, 1353–1359. [Google Scholar] [CrossRef] [Green Version]
- Young, R.L.; Sutherland, K.; Pezos, N.; Brierley, S.M.; Horowitz, M.; Rayner, C.K.; Blackshaw, L.A. Expression of taste molecules in the upper gastrointestinal tract in humans with and without type 2 diabetes. Gut 2009, 58, 337–346. [Google Scholar] [CrossRef]
- Nicklaus, S.; Boggio, V.; Chabanet, C.; Issanchou, S. A prospective study of food variety seeking in childhood, adolescence and early adult life. Appetite 2005, 44, 289–297. [Google Scholar] [CrossRef]
- Garcia-Bailo, B.; Toguri, C.; Eny, K.M.; El-Sohemy, A. Genetic variation in taste and its influence on food selection. OMICS 2009, 13, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Salbe, A.D.; DelParigi, A.; Pratley, R.E.; Drewnowski, A.; Tataranni, P.A. Taste preferences and body weight changes in an obesity-prone population. Am. J. Clin. Nutr. 2004, 79, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Eny, K.M.; Wolever, T.M.; Corey, P.N.; El-Sohemy, A. Genetic variation in TAS1R2 (Ile191Val) is associated with consumption of sugars in overweight and obese individuals in 2 distinct populations. Am. J. Clin. Nutr. 2010, 92, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Bartoshuk, L.M.; Duffy, V.B.; Hayes, J.E.; Moskowitz, H.R.; Snyder, D.J. Psychophysics of sweet and fat perception in obesity: Problems, solutions and new perspectives. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1137–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, A.G.; Eny, K.M.; Cockburn, M.; Chiu, W.; Nielsen, D.E.; Duizer, L.; El-Sohemy, A. Variation in the TAS1R2 Gene, Sweet Taste Perception and Intake of Sugars. J. Nutrigenet. Nutr. 2015, 8, 81–90. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Start Material | Solvent Extract | Effects | Type of Study | Ref. |
|---|---|---|---|---|
| Powder seeds | 95% Ethanol | Cholesterol-lowering agent | Preclinical | [49] |
| Air-dried leaves | 80% Methanol | Antidiabetic | Preclinical | [45,48] |
| Air-dried skin+pulp | 95% EtOH | Uptake of glucose | In vitro (C2C12) | [43] |
| Miracle fruit powder | Water/butanol | Antioxidant; Anti-hyperuricemia | In vitro, Preclinical | [50] |
| Leaves, stems, and berries dried | 80% MeOH, 10% EtOH | Reduce proliferation of colorectal cancer cell lines | In vitro | [51] |
| Stems | MeOH | Reduce proliferation in melanoma cell lines | In vitro | [36] |
| Pulp and fruit | MeOH-fruit | Anti-tyrosinase and antioxidation effects | In vitro | [44] |
| CHCl3-pulp | ||||
| Miracle fruit powder | No extraction | Improve insulin resistance | Preclinical | [41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez de Cedrón, M.; Wagner, S.; Reguero, M.; Menéndez-Rey, A.; Ramírez de Molina, A. Miracle Berry as a Potential Supplement in the Control of Metabolic Risk Factors in Cancer. Antioxidants 2020, 9, 1282. https://doi.org/10.3390/antiox9121282
Gómez de Cedrón M, Wagner S, Reguero M, Menéndez-Rey A, Ramírez de Molina A. Miracle Berry as a Potential Supplement in the Control of Metabolic Risk Factors in Cancer. Antioxidants. 2020; 9(12):1282. https://doi.org/10.3390/antiox9121282
Chicago/Turabian StyleGómez de Cedrón, Marta, Sonia Wagner, Marina Reguero, Adrián Menéndez-Rey, and Ana Ramírez de Molina. 2020. "Miracle Berry as a Potential Supplement in the Control of Metabolic Risk Factors in Cancer" Antioxidants 9, no. 12: 1282. https://doi.org/10.3390/antiox9121282
APA StyleGómez de Cedrón, M., Wagner, S., Reguero, M., Menéndez-Rey, A., & Ramírez de Molina, A. (2020). Miracle Berry as a Potential Supplement in the Control of Metabolic Risk Factors in Cancer. Antioxidants, 9(12), 1282. https://doi.org/10.3390/antiox9121282