Nutrients, Antinutrients, Phenolic Composition, and Antioxidant Activity of Common Bean Cultivars and their Potential for Food Applications

 ,
,  ,
,  ,
, 
Abstract
:
1. Introduction
2. Material and Methods
2.1. Sampling
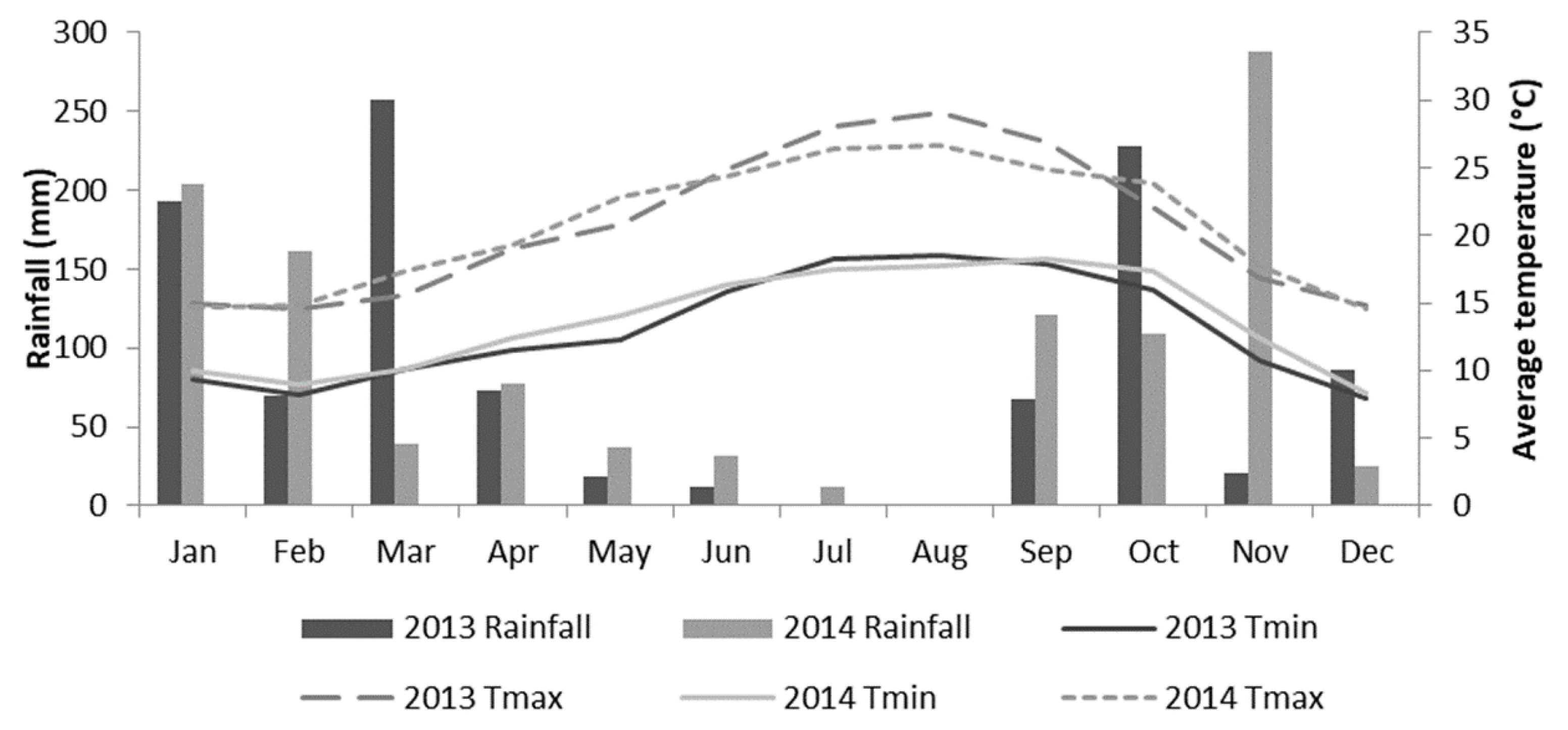
2.2. Climatic Conditions
2.3. Chemicals and Reagents
2.4. Nutritional Composition
2.4.1. Protein content
2.4.2. Amino Acids Composition
2.5. Antinutritional Composition
2.6. Phenolic Composition
2.6.1. Total Phenols
2.6.2. Ortho-Diphenols
2.6.3. Flavonoids
2.7. Individual Phenolic Compounds
2.8. In Vitro Antioxidant Activities
2.9. Statistical Analysis
3. Results and Discussion
3.1. Nutritional Composition
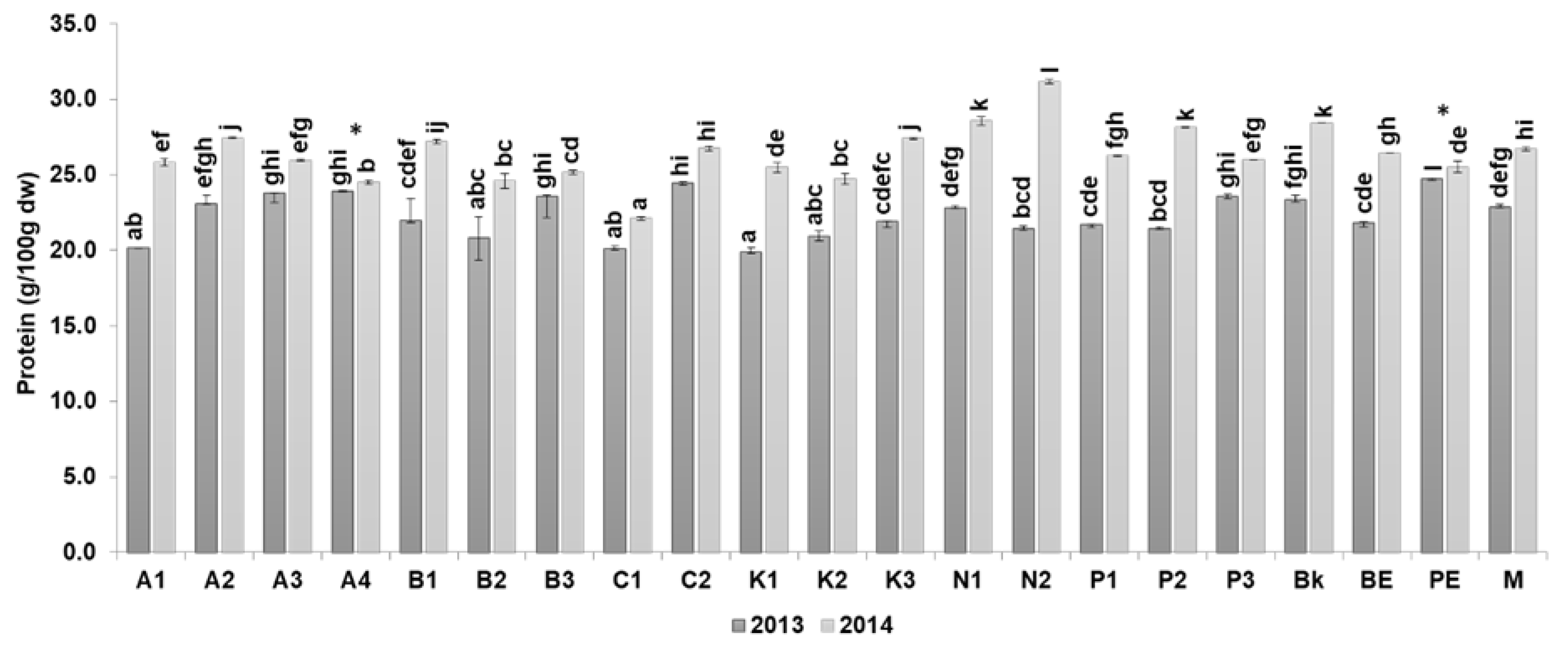
3.1.1. Protein Content
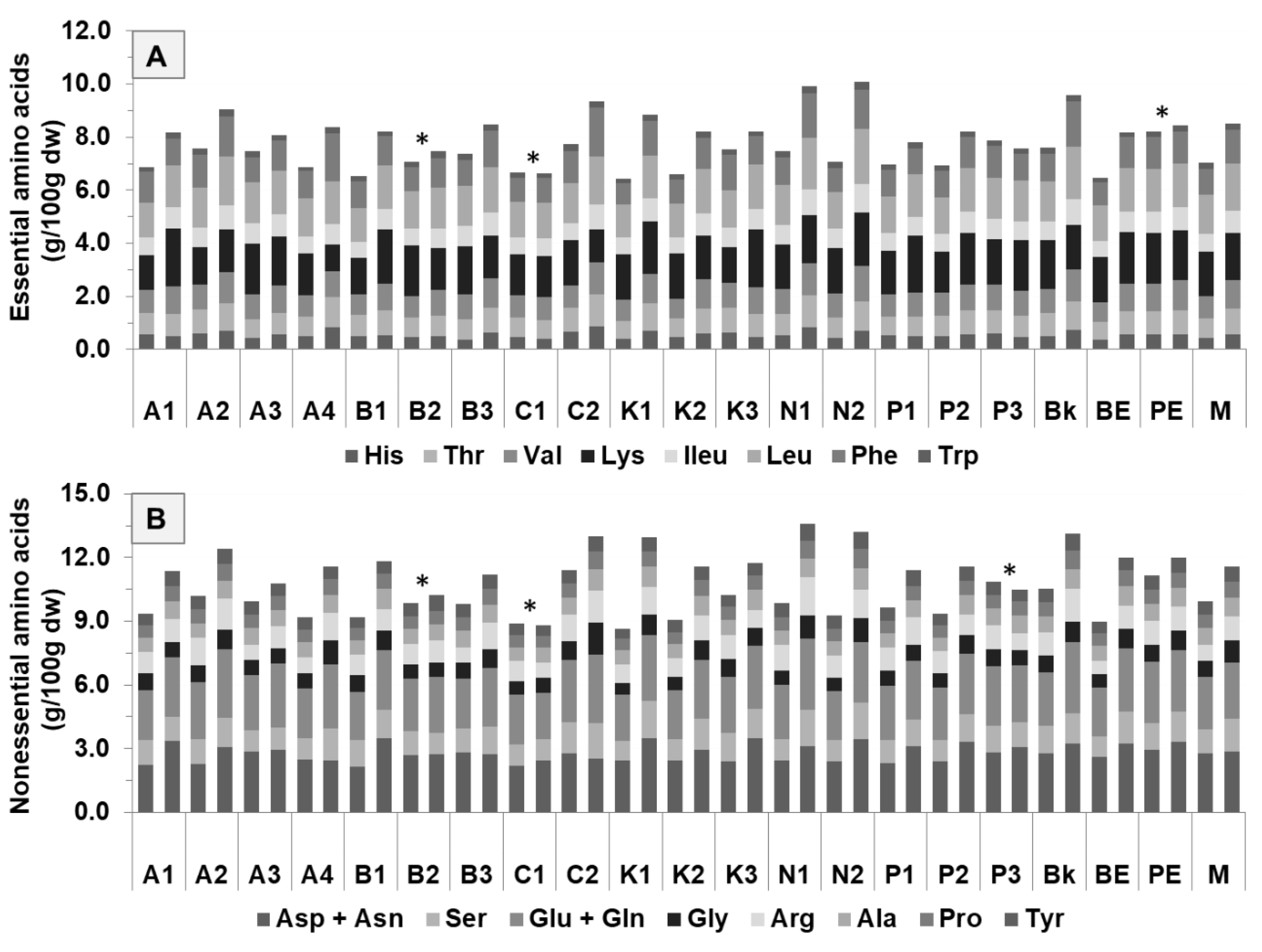
3.1.2. Amino Acids Composition
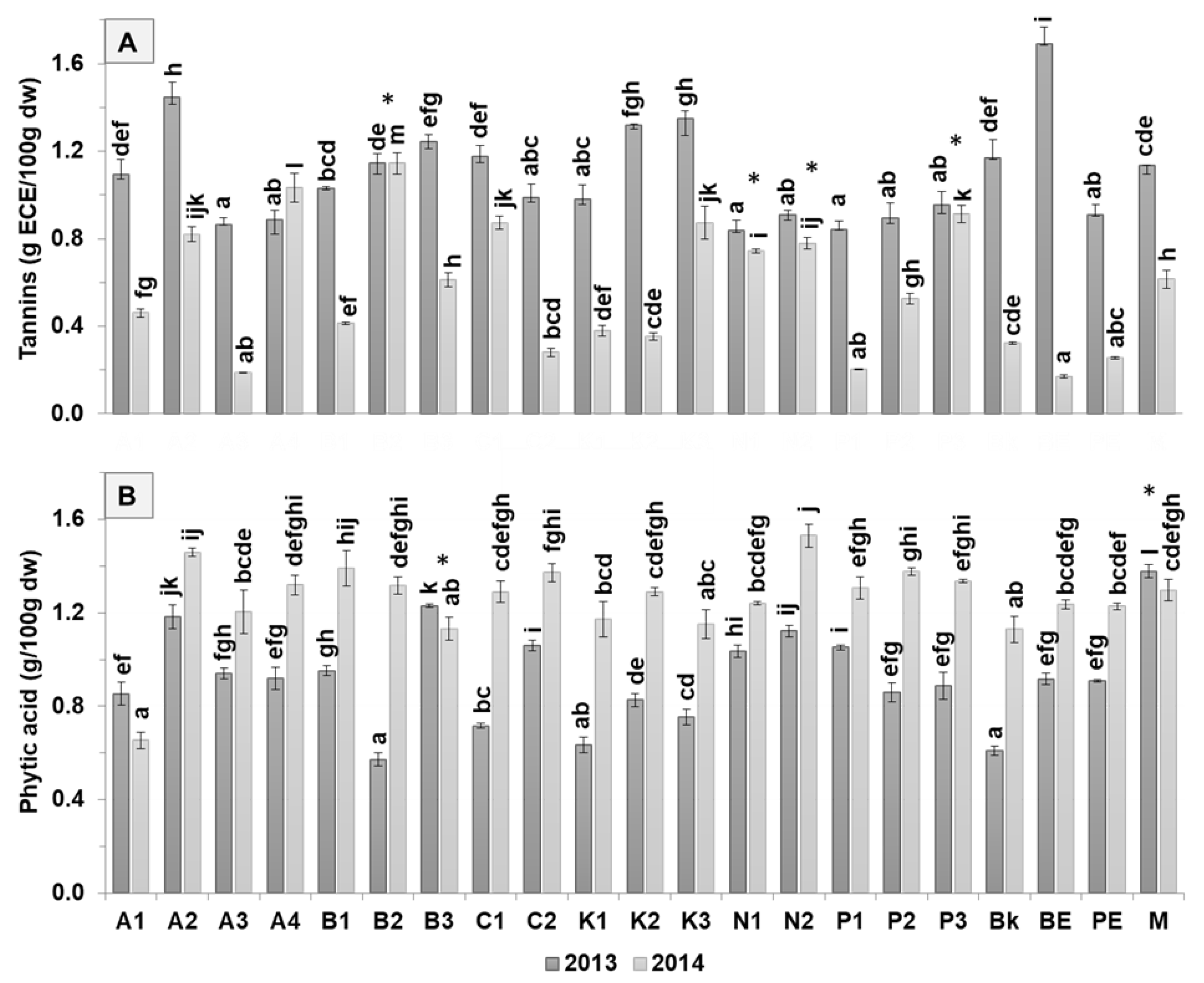
3.2. Antinutritional Composition
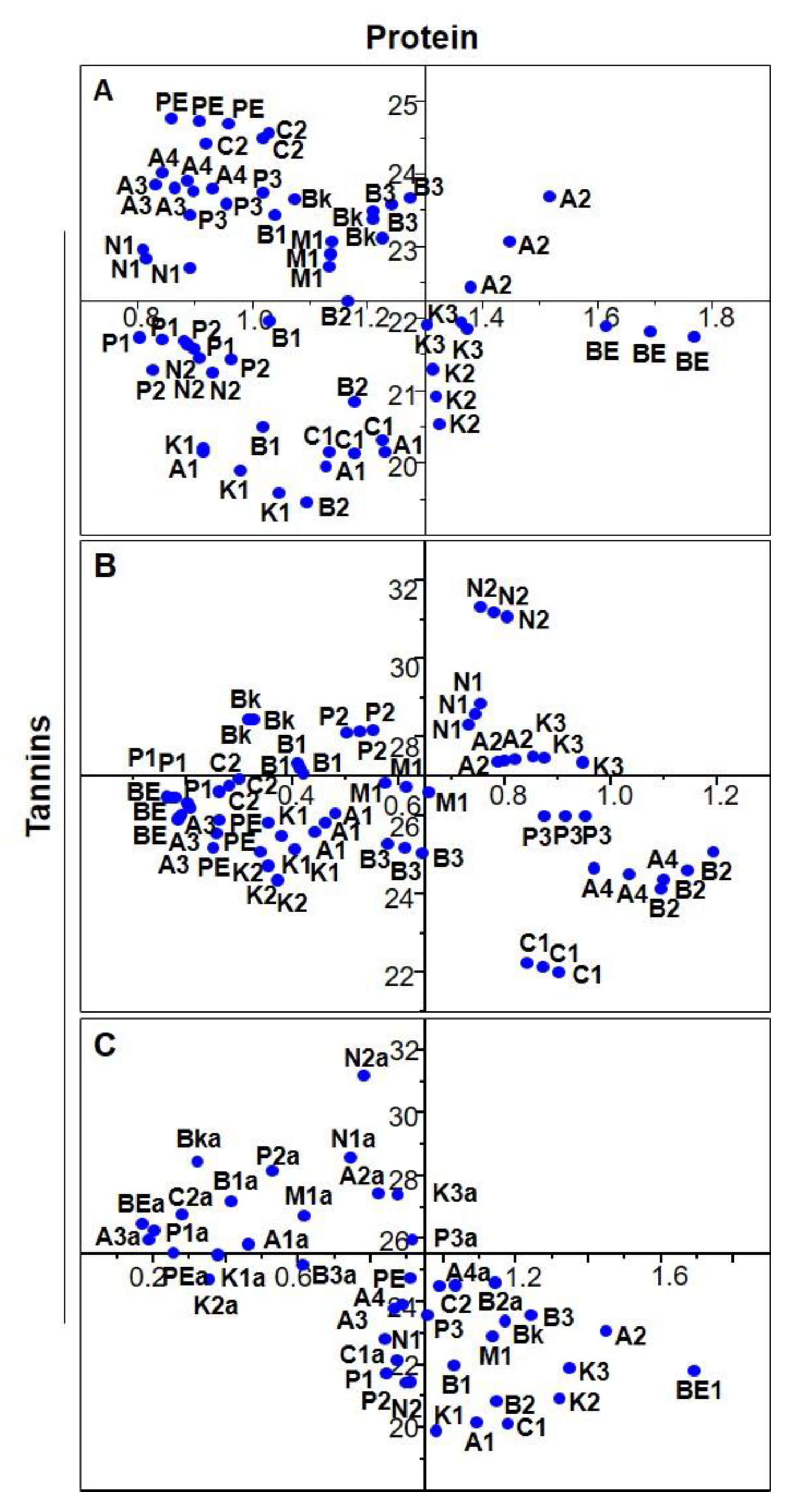
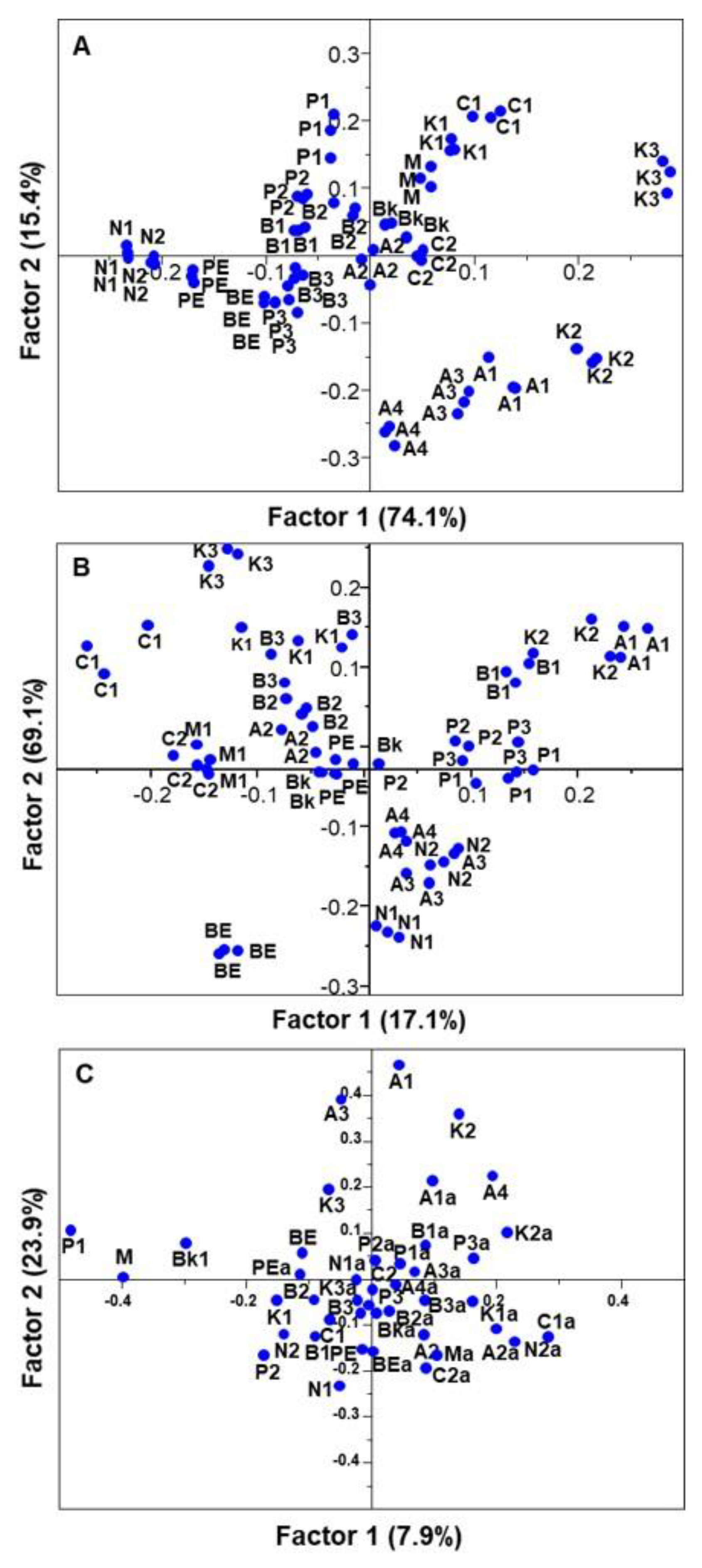
3.3. Phenolic Composition
3.4. Individual Phenolic Compounds
3.5. In Vitro Antioxidant Activities
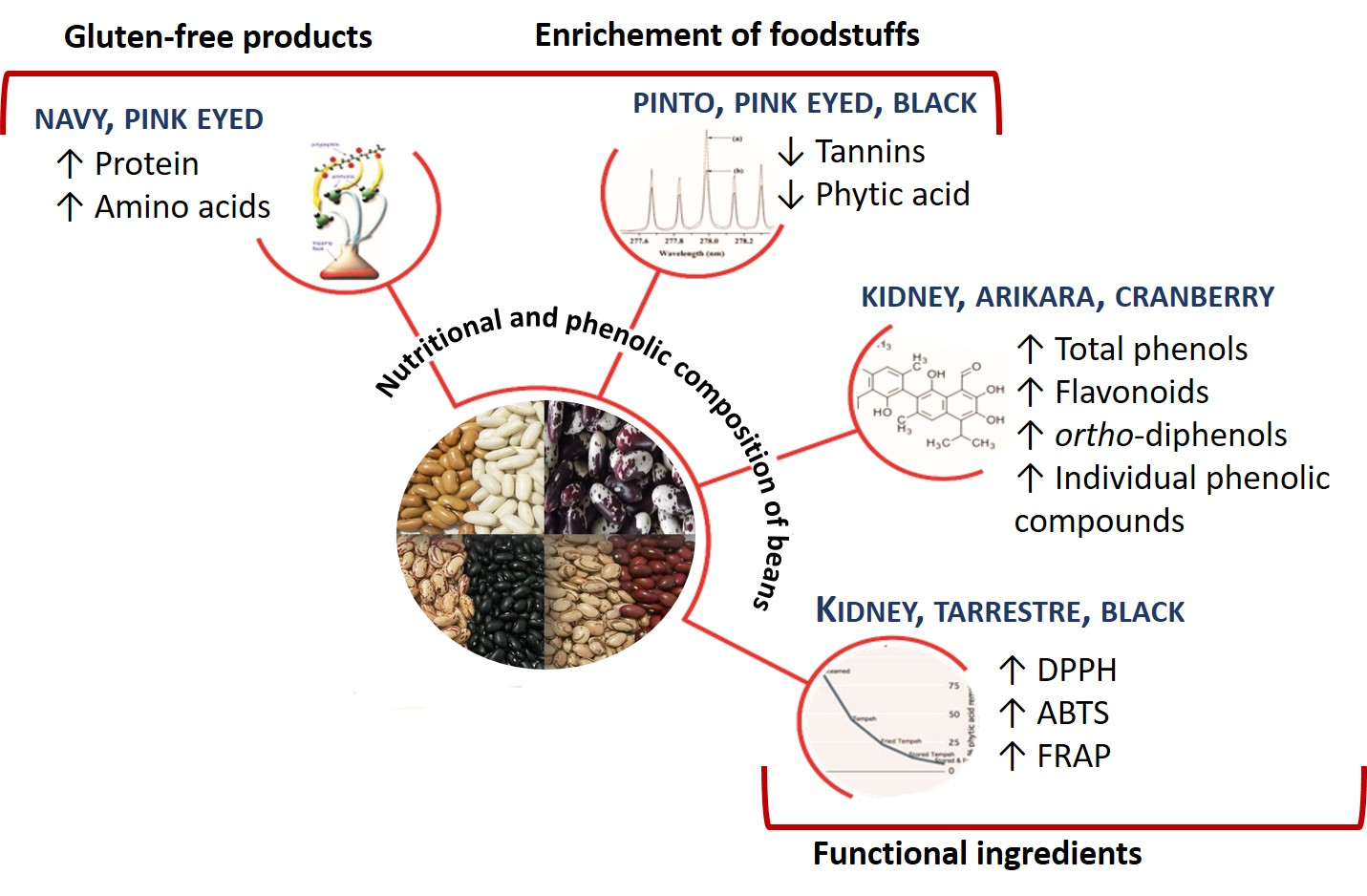
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ramírez-Jiménez, A.K.; Gaytán-Martínez, M.; Morales-Sánchez, E.; Loarca-Piña, G. Functional properties and sensory value of snack bars added with common bean flour as a source of bioactive compounds. LWT 2018, 89, 674–680. [Google Scholar] [CrossRef]
- Foyer, C.H.; Lam, H.M.; Nguyen, H.T.; Siddique, K.H.M.; Varshney, R.K.; Colmer, T.D.; Cowling, W.; Bramley, H.; Mori, T.A.; Hodgson, J.M.; et al. Neglecting legumes has compromised human health and sustainable food production. Nat. Plants 2016, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kan, L.; Nie, S.; Hu, J.; Wang, S.; Bai, Z.; Wang, J.; Zhou, Y.; Jiang, J.; Zeng, Q.; Song, K. Comparative study on the chemical composition, anthocyanins, tocopherols and carotenoids of selected legumes. Food Chem. 2018, 260, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Giusti, F.; Caprioli, G.; Ricciutelli, M.; Vittori, S.; Sagratini, G. Determination of fourteen polyphenols in pulses by high performance liquid chromatography-diode array detection (HPLC-DAD) and correlation study with antioxidant activity and colour. Food Chem. 2017, 221, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Kan, L.; Nie, S.; Hu, J.; Wang, S.; Cui, S.W.; Li, Y.; Xu, S.; Wu, Y.; Wang, J.; Bai, Z.; et al. Nutrients, phytochemicals and antioxidant activities of 26 kidney bean cultivars. Food Chem. Toxicol. 2017, 108, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Hatcher, D.W.; Tyler, R.T.; Toews, R.; Gawalko, E.J. Effect of cooking on the composition of beans (Phaseolus vulgaris L.) and chickpeas (Cicer arietinum L.). Food Res. Int. 2010, 43, 589–594. [Google Scholar] [CrossRef]
- Dueñas, M.; Martínez-Villaluenga, C.; Limón, R.I.; Peñas, E.; Frias, J. Effect of germination and elicitation on phenolic composition and bioactivity of kidney beans. Food Res. Int. 2015, 70, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Toews, R.; Wang, N. Physicochemical and functional properties of protein concentrates from pulses. Food Res. Int. 2013, 52, 445–451. [Google Scholar] [CrossRef]
- Cominelli, E.; Confalonieri, M.; Carlessi, M.; Cortinovis, G.; Daminati, M.G.; Porch, T.G.; Losa, A.; Sparvoli, F. Phytic acid transport in Phaseolus vulgaris: A new low phytic acid mutant in the PvMRP1 gene and study of the PvMRPs promoters in two different plant systems. Plant Sci. 2018, 270, 1–12. [Google Scholar] [CrossRef]
- Chen, P.X.; Zhang, H.; Marcone, M.F.; Pauls, K.P.; Liu, R.; Tang, Y.; Zhang, B.; Renaud, J.B. Anti-inflammatory effects of phenolic-rich cranberry bean (Phaseolus vulgaris L.) extracts and enhanced cellular antioxidant enzyme activities in Caco-2 cells. J. Funct. Foods 2017, 38, 675–685. [Google Scholar] [CrossRef]
- Moreno-Jiménez, M.R.; Cervantes-Cardoza, V.; Gallegos-Infante, J.A.; González-Laredo, R.F.; Estrella, I.; de J. García-Gasca, T.; Herrera-Carrera, E.; Díaz-Rivas, J.O.; Rocha-Guzmán, N.E. Phenolic composition changes of processed common beans: Their antioxidant and anti-inflammatory effects in intestinal cancer cells. Food Res. Int. 2015, 76, 79–85. [Google Scholar]
- Mojica, L.; Gonzalez de Mejia, E.; Granados-Silvestre, M.Á.; Menjivar, M. Evaluation of the hypoglycemic potential of a black bean hydrolyzed protein isolate and its pure peptides using in silico, in vitro and in vivo approaches. J. Funct. Foods 2017, 31, 274–286. [Google Scholar] [CrossRef]
- Reverri, E.J.; Randolph, J.M.; Kappagoda, C.T.; Park, E.; Edirisinghe, I.; Burton-Freeman, B.M. Assessing beans as a source of intrinsic fiber on satiety in men and women with metabolic syndrome. Appetite 2017, 118, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Giuberti, G.; Gallo, A.; Cerioli, C.; Fortunati, P.; Masoero, F. Cooking quality and starch digestibility of gluten free pasta using new bean flour. Food Chem. 2015, 175, 43–49. [Google Scholar] [CrossRef]
- Sparvoli, F.; Laureati, M.; Pilu, R.; Pagliarini, E.; Toschi, I.; Giuberti, G.; Fortunati, P.; Daminati, M.G.; Cominelli, E.; Bollini, R. Exploitation of Common Bean Flours with Low Antinutrient Content for Making Nutritionally Enhanced Biscuits. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Siddiq, M.; Kelkar, S.; Harte, J.B.; Dolan, K.D.; Nyombaire, G. Functional properties of flour from low-temperature extruded navy and pinto beans (Phaseolus vulgaris L.). LWT-Food Sci. Technol. 2013, 50, 215–219. [Google Scholar] [CrossRef]
- Bhol, S.; John Don Bosco, S. Influence of malted finger millet and red kidney bean flour on quality characteristics of developed bread. LWT-Food Sci. Technol. 2014, 55, 294–300. [Google Scholar] [CrossRef]
- Gallegos-Infante, J.A.; Rocha-Guzman, N.E.; Gonzalez-Laredo, R.F.; Ochoa-Martínez, L.A.; Corzo, N.; Bello-Perez, L.A.; Medina-Torres, L.; Peralta-Alvarez, L.E. Quality of spaghetti pasta containing Mexican common bean flour (Phaseolus vulgaris L.). Food Chem. 2010, 119, 1544–1549. [Google Scholar] [CrossRef]
- Carbas, B.; Pathania, S.; Castanho, A.; Lourenço, D.; Veiga, I.M.; Patto, M.C.V.; Brites, C. Elucidating potential utilization of Portuguese common bean varieties in rice based processed foods. J. Food Sci. Technol. 2018, 55, 1056–1064. [Google Scholar] [CrossRef]
- Yang, Y.; He, Q.; Sun, H.; Cao, X.; Elfalleh, W.; Wu, Z.; Zhao, J.; Sun, X.; Zhang, Y.; He, S. PEGylation may reduce allergenicity and improve gelling properties of protein isolate from black kidney bean (Phaseolus vulgaris L.). Food Biosci. 2018, 25, 83–90. [Google Scholar] [CrossRef]
- Messina, V. Nutritional and health benefits of dried beans. Am. J. Clin. Nutr. 2014, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor-Cavada, E.; Juan, R.; Pastor, J.E.; Alaiz, M.; Vioque, J. Protein and amino acid composition of select wild legume species of tribe Fabeae. Food Chem. 2014, 163, 97–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haylock, M.R.; Hofstra, N.; Tank, A.M.G.K.; Klok, E.J.; Jones, P.D.; New, M. A European daily high-resolution gridded data set of surface temperature and precipitation for 1950–2006. J. Geophys. Res. Atmos. 2008, 113. [Google Scholar] [CrossRef] [Green Version]
- Machado, N.; Oppolzer, D.; Ramos, A.; Ferreira, L.; Rosa, E.A.S.; Rodrigues, M.; Domínguez-Perles, R.; Barros, A.I.R.N.A. Evaluating the freezing impact on the proximate composition of immature cowpea (Vigna unguiculata L.) pods: Classical versus spectroscopic approaches. J. Sci. Food Agric. 2017, 97, 4295–4305. [Google Scholar] [CrossRef]
- Dambergs, R.G.; Mercurio, M.D.; Kassara, S.; Cozzolino, D.; Smith, P.A. Rapid measurement of methyl cellulose precipitable tannins using ultraviolet spectroscopy with chemometrics: Application to red wine and inter-laboratory calibration transfer. Appl. Spectrosc. 2012, 66, 656–664. [Google Scholar] [CrossRef]
- Mena, P.; García-Viguera, C.; Navarro-Rico, J.; Moreno, D.A.; Bartual, J.; Saura, D.; Martí, N. Phytochemical characterisation for industrial use of pomegranate (Punica granatum L.) cultivars grown in Spain. J. Sci. Food Agric. 2011, 91, 1893–1906. [Google Scholar] [CrossRef]
- Mateos, R.; Espartero, J.L.; Trujillo, M.; Ríos, J.J.; León-Camacho, M.; Alcudia, F.; Cert, A. Determination of phenols, flavones, and lignans in virgin olive oils by solid-phase extraction and high-performance liquid chromatography with diode array ultraviolet detection. J. Agric. Food Chem. 2001, 49, 2185–2192. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Lin, L.-Z.; Harnly, J.M.; Pastor-Corrales, M.S.; Luthria, D.L. The polyphenolic profiles of common bean (Phaseolus vulgaris L.). Food Chem. 2008, 107, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Espín, J.C.; Soler-Rivas, C.; Wichers, H.J.; García-Viguera, C. Anthocyanin-based natural colorants: A new source of antiradical activity for foodstuff. J. Agric. Food Chem. 2000, 48, 1588–1592. [Google Scholar] [CrossRef]
- Heinemann, A.B.; Ramirez-Villegas, J.; Souza, T.L.P.O.; Didonet, A.D.; di Stefano, J.G.; Boote, K.J.; Jarvis, A. Drought impact on rainfed common bean production areas in Brazil. Agric. For. Meteorol. 2016, 225, 57–74. [Google Scholar] [CrossRef] [Green Version]
- Du, S.; Jiang, H.; Yu, X.; Jane, J. Physicochemical and functional properties of whole legume flour. LWT-Food Sci. Technol. 2014, 55, 308–313. [Google Scholar] [CrossRef]
- Borresen, E.; Gundlach, K.; Wdowik, M.; Rao, S.; Brown, R.; Ryan, E. Feasibility of Increased Navy Bean Powder Consumption for Primary and Secondary Colorectal Cancer Prevention. Curr. Nutr. Food Sci. 2014, 10, 112–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbonaro, M.; Maselli, P.; Nucara, A. Structural aspects of legume proteins and nutraceutical properties. Food Res. Int. 2015, 76, 19–30. [Google Scholar] [CrossRef]
- Grela, E.R.; Kiczorowska, B.; Samolińska, W.; Matras, J.; Kiczorowski, P.; Rybiński, W.; Hanczakowska, E. Chemical composition of leguminous seeds: Part I—Content of basic nutrients, amino acids, phytochemical compounds, and antioxidant activity. Eur. Food Res. Technol. 2017, 243, 1385–1395. [Google Scholar] [CrossRef]
- Segura-Campos, M.R.; García-Rodríguez, K.; Ruiz-Ruiz, J.C.; Chel-Guerrero, L.; Betancur-Ancona, D. In vitro bioactivity, nutritional and sensory properties of semolina pasta added with hard-to-cook bean (Phaseolus vulgaris L.) protein hydrolysate. J. Funct. Foods 2014, 8, 1–8. [Google Scholar] [CrossRef]
- Campos-Vega, R.; Loarca-Piña, G.; Oomah, B.D. Minor components of pulses and their potential impact on human health. Food Res. Int. 2010, 43, 461–482. [Google Scholar] [CrossRef]
- Chávez-Mendoza, C.; Hernández-Figueroa, K.I.; Sánchez, E. Antioxidant capacity and phytonutrient content in the seed coat and cotyledon of common beans (Phaseolus vulgaris L.) from various regions in Mexico. Antioxidants 2019, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Rocchetti, G.; Lucini, L.; Rodriguez, J.M.L.; Barba, F.J.; Giuberti, G. Gluten-free flours from cereals, pseudocereals and legumes: Phenolic fingerprints and in vitro antioxidant properties. Food Chem. 2019, 271, 157–164. [Google Scholar] [CrossRef]
- Zhao, Y.; Du, S.; Wang, H.; Cai, M. In vitro antioxidant activity of extracts from common legumes. Food Chem. 2014, 152, 462–466. [Google Scholar] [CrossRef]
- Mecha, E.; Leitão, S.T.; Carbas, B.; Serra, A.T.; Moreira, P.M.; Veloso, M.M.; Gomes, R.; Figueira, M.E.; Brites, C.; Vaz Patto, M.C.; et al. Characterization of Soaking Process’ Impact in Common Beans Phenolic Composition: Contribute from the Unexplored Portuguese Germplasm. Foods 2019, 8, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilera, Y.; Estrella, I.; Benitez, V.; Esteban, R.M.; Martín-Cabrejas, M.A. Bioactive phenolic compounds and functional properties of dehydrated bean flours. Food Res. Int. 2011, 44, 774–780. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivars | Total Phenols | Ortho-Diphenols | Flavonoids | |||
|---|---|---|---|---|---|---|
| 2013 | 2014 | 2013 | 2014 | 2013 | 2014 | |
| A1 | 2.97 ± 0.24 j,k,* | 2.98 ± 0.08 i,j,k | 6.46 ± 0.34 i,j,* | 5.07 ± 0.17 k | 2.63 ± 0.12 I,* | 2.47 ± 0.22 h,i |
| A2 | 2.32 ± 0.22 h,I,* | 2.79 ± 0.22 h,i | 2.54 ± 0.11 d,e,f,* | 2.78 ± 0.07 d,e,f,g | 2.18 ± 0.08 g,* | 2.30 ± 0.12 g,h,i |
| A3 | 2.27 ± 0.05 g,h | 1.16 ± 0.11 b | 6.16 ± 0.07 i | 3.26 ± 0.17 g,h,i | 1.62 ± 0.04 d,e,f,* | 1.36 ± 0.12 b,c |
| A4 | 2.30 ± 0.05 h | 1.36 ± 0.07 b,c | 5.53 ± 0.08 h | 3.00 ± 0.06 e,f,g,h | 1.33 ± 0.04 c,d,* | 1.19 ± 0.00 a,b |
| B1 | 1.38 ± 0.06 d,e | 2.87 ± 0.21 i,j | 1.93 ± 0.03 b,c | 3.97 ± 0.05 j | 1.76 ± 0.05 e,f,* | 1.93 ± 0.01 d,e,f |
| B2 | 1.89 ± 0.18 f,g | 2.67 ± 0.26 g,h,i | 2.42 ± 0.11 d,e,* | 2.84 ± 0.06 d,e,f,g | 2.28 ± 0.14 g,h,* | 2.17 ± 0.14 f,g,h |
| B3 | 1.41 ± 0.10 d,e | 3.03 ± 0.14 i,j,k | 2.39 ± 0.07 d,e | 3.00 ± 0.20 e,f,g,h | 1.70 ± 0.09 e,f | 2.53 ± 0.22 i |
| C1 | 3.17 ± 0.18 k,* | 3.39 ± 0.30 k,l | 2.23 ± 0.03 c,d | 2.73 ± 0.24 c,d,e,f | 3.90 ± 0.13 k,* | 4.02 ± 0.04 l |
| C2 | 2.75 ± 0.05 j,* | 2.61 ± 0.17 f,g,h,i | 2.87 ± 0.16 f | 2.06 ± 0.03 a,b | 2.53 ± 0.15 h,i | 2.14 ± 0.01 e,f,g |
| K1 | 2.70 ± 0.16 i,j | 3.37 ± 0.23 j,k,l | 2.50 ± 0.06 d,e,f | 3.42 ± 0.27 h,i | 3.10 ± 0.05 j,* | 3.00 ± 0.16 j |
| K2 | 4.03 ± 0.19 l | 3.31 ± 0.11 j,k | 6.69 ± 0.27 j | 4.86 ± 0.38 k | 2.81 ± 0.02 i,j | 2.19 ± 0.04 f,g,h |
| K3 | 4.59 ± 0.24 m | 3.84 ± 0.13 l | 4.70 ± 0.15 g | 3.40 ± 0.10 h,i | 4.33 ± 0.25 l | 3.35 ± 0.08 k |
| N1 | 0.11 ± 0.00 a,* | 0.43 ± 0.01 a | 0.89 ± 0.06 a | 2.37 ± 0.07 a,b,c,d | 0.80 ± 0.03 a,* | 0.97 ± 0.04 a |
| N2 | 0.35 ± 0.01 a,b | 2.13 ± 0.13 d,e,f | 1.07 ± 0.07 a | 2.23 ± 0.09 a,b,c | 0.98 ± 0.03 a,b,* | 0.91 ± 0.02 a |
| P1 | 1.39 ± 0.02 d,e | 2.06 ± 0.04 d,e | 2.57 ± 0.04 d,e,f | 3.53 ± 0.17 i,j | 1.78 ± 0.11 e,f | 1.29 ± 0.03 b |
| P2 | 1.30 ± 0.02 d,e | 2.04 ± 0.18 d | 1.72 ± 0.06 b | 3.25 ± 0.29 f,g,h,i | 1.59 ± 0.08 d,e,f,* | 1.71 ± 0.07 d |
| P3 | 1.53 ± 0.02 e,f | 2.30 ± 0.20 d,e,f,g,h | 2.28 ± 0.13 c,d,e | 3.58 ± 0.21 i,j | 1.65 ± 0.01 e,f,* | 1.83 ± 0.15 d,e |
| Bk | 2.25 ± 0.06 g,h | 2.21 ± 0.12 d,e,f,g | 2.91 ± 0.08 f | 2.87 ± 0.07 d,e,f,g | 1.78 ± 0.01 e,f | 1.67 ± 0.06 c,d |
| BE | 1.03 ± 0.03 c,d,* | 0.65 ± 0.02 a | 2.58 ± 0.20 d,e,f | 1.85 ± 0.07 a | 1.54 ± 0.06 d,e | 1.09 ± 0.08 a,b |
| PE | 0.67 ± 0.06 b,c,* | 1.84 ± 0.12 c,d | 1.53 ± 0.04 b | 2.52 ± 0.04 b,c,d,e | 1.23 ± 0.01 b,c,* | 2.04 ± 0.09 e,f,g |
| M | 1.96 ± 0.18 g,h | 2.56 ± 0.10 e,f,g,h,i | 2.68 ± 0.11 e,f | 2.40 ± 0.01 b,c,d | 1.84 ± 0.10 f | 2.23 ± 0.03 f,g,h,i |
| Cultivars | Myricetin 3-O-glucoside | Quercetin 3-O-glucoside | Quercetin 3-O-(6´´-O-manolyl) glucoside | Kaempferol 3-O-glucoside | Myricetin | Kaempferol 3-O-(6´´-O-manolyl)glucoside | Kaempferol 3-O-(malonyl)glucoside | Kaempferol | Protocatechuicacid | Gallic acid | Catechin | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2013 | 2014 | 2013 | 2014 | 2013 | 2014 | 2013 | 2014 | 2013 | 2014 | 2013 | 2014 | 2013 | 2014 | 2013 | 2014 | 2013 | 2014 | 2013 | 2014 | 2013 | 2014 | |
| A1 | nd | nd | 126.9 | 41.7 | nd | nd | 1767.1 | 974.6 | nd | nd | 89.8 | 119.0 | 5.2 * | 5.7 | 10.3 | 5.6 | nd | nd | nd | nd | 324.7 | 370.5 |
| A2 | nd | 10.4 | 6.0 * | 5.7 | nd | nd | 184.1 * | 114.0 | nd | nd | 43.7 * | 45.4 | nd | nd | 48.3 | 31.3 | nd | nd | nd | nd | 625.0 | 554.9 |
| A3 | nd | nd | 104.1 | 12.0 | nd | nd | 1322.6 | 570.2 | nd | nd | 62.3 | 40.0 | nd | nd | 16.8 | 8.9 | 15.4 | 17.7 | nd | nd | 1562.8 | 381.4 |
| A4 | nd | nd | 78.2 | 11.2 | nd | nd | 1171.4 | 504.4 | nd | nd | 79.4 | 61.1 | nd | 4.7 | 10.1 | nd | nd | nd | nd | nd | 765.1 | 550.6 |
| B1 | nd | nd | 37.1 * | 36.0 | 5.3 | 8.1 | 49.0 * | 37.4 | nd | nd | 20.6 * | 24.0 | nd | nd | nd | nd | nd | nd | nd | nd | 1004.4 | 1412.3 |
| B2 | nd | nd | 61.3 | 30.3 | 9.0 | 6.5 | 73.5 | 46.0 | nd | nd | 22.6 | 33.3 | nd | nd | nd | nd | nd | nd | nd | nd | 856.8 | 433.3 |
| B3 | nd | nd | 1.6 | nd | nd | nd | 74.8 | 41.3 | nd | nd | 37.8 * | 39.2 | 8.6 | nd | 163.4 | 93.3 | nd | nd | nd | nd | 253.5 * | 209.8 |
| C1 | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | 594.4 | 740.0 |
| C2 | nd | nd | nd | nd | nd | nd | 16.8 | 8.6 | nd | nd | 16.1 | nd | nd | nd | nd | nd | nd | nd | nd | nd | 1569.9 | 543.0 |
| K1 | nd | nd | 13.8 | 44.7 | nd | 3.3 | 56.2 * | 56.4 | 29.7 | 19.3 | 5.6 * | 12.2 | nd | nd | nd | nd | nd | nd | nd | nd | 380.6 * | 373.6 |
| K2 | nd | nd | 90.4 | 35.8 | 2.7 | nd | 1740.6 | 761.2 | 11.6 * | 12.4 | 199.0 | 132.5 | 10.2 | 6.8 | 19.5 | 9.3 | nd | nd | nd | nd | 668.1 | 455.3 |
| K3 | nd | nd | 53.2 | 47.3 | nd | 3.3 | 96.3 * | 75.9 | 25.4 | 18.9 | 9.5 * | 12.8 | nd | nd | nd | nd | nd | nd | nd | nd | 724.6 | 380.1 |
| N1 | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | 54.0 | 63.2 | 763.4 * | 761.2 |
| N2 | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | 67.9 | 63.7 | 658.4 * | 555.0 |
| P1 | nd | nd | 8.2 | 3.0 | nd | nd | 158.5 | 74 | nd | nd | 18.5* | 20.9 | nd | nd | 14.3 * | 14.1 | nd | nd | nd | nd | 1380.7 | 1027.7 |
| P2 | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | 563.1 * | 559.0 |
| P3 | nd | nd | 33.8 | nd | nd | nd | 245.2 | 516.9 | nd | nd | 19.1 | 75.9 | nd | 3.6 | 5.3 | nd | nd | nd | nd | nd | 1057.0 | 202.0 |
| Bk | 150.2 | 79.7 | 22.2 | 14.7 | nd | nd | 17.5 * | 12.4 | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | 1003.2 | 206.5 |
| BE | nd | nd | nd | nd | nd | nd | nd | 15.6 | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | 666.3 | 268.4 |
| PE | 11.7 | 22.6 | 31.2 | 87.3 | nd | 3.0 | 18.6 | 75.0 | nd | 10.2 | nd | 12.9 | nd | nd | nd | nd | nd | nd | nd | nd | 759.0 | 119.1 |
| M | nd | nd | 4.8 | nd | nd | nd | 140.6 | 40.1 | nd | nd | 32.7 | 21.0 | nd | nd | 44.9 | 12.1 | nd | nd | nd | nd | 737.0 | 309.3 |
| Cultivars | DPPH• | ABTS• | FRAP | |||
|---|---|---|---|---|---|---|
| 2013 | 2014 | 2013 | 2014 | 2013 | 2014 | |
| A1 | 8.93 ± 0.20 d,e,* | 10.39 ± 0.61 c,d,e | 19.94 ± 0.58 f | 14.93 ± 0.70 e,f | 18.10 ± 0.54 g,h,* | 16.61 ± 0.69 i |
| A2 | 11.39 ± 0.80 g,h | 9.87 ± 0.23 c,d | 16.39 ± 0.35 e,* | 15.16 ± 0.90 e,f,g | 11.14 ± 0.65 c,d,* | 10.15 ± 0.06 c,d |
| A3 | 10.01 ± 0.24 e,f,g | 7.15 ± 0.17 b | 20.36 ± 0.75 f,g | 9.10 ± 0.44 b,c | 16.48 ± 0.86 f,g | 7.50 ± 0.62 b |
| A4 | 8.22 ± 0.27 d,* | 7.19 ± 0.05 b | 13.25 ± 0.56 c,d,* | 13.72 ± 0.69 d,e | 9.96 ± 0.15 c,d | 7.68 ± 0.13 b |
| B1 | 10.13 ± 0.49 e,f,g,* | 9.16 ± 0.59 c | 14.21 ± 0.40 c,d,e | 16.96 ± 0.66 f,g | 11.64 ± 0.33 d | 14.13 ± 0.36 g |
| B2 | 10.43 ± 0.36 f,g,* | 11.22 ± 0.44 e,f | 15.39 ± 1.06 d,e,* | 17.69 ± 1.06 g,h | 14.73 ± 0.23 e,f,* | 14.33 ± 0.34 g,h |
| B3 | 8.88 ± 0.50 d,e | 11.34 ± 0.21 e,f | 13.26 ± 0.70 c,d | 21.00 ± 1.14 i | 9.96 ± 0.36 c,d | 13.84 ± 0.73 g |
| C1 | 15.82 ± 0.06 j | 14.81 ± 0.70 g | 23.87 ± 1.11 h,i | 20.98 ± 1.57 i | 19.68 ± 0.65 h,i | 11.14 ± 0.40 d,e |
| C2 | 10.98 ± 0.63 g,h,* | 10.95 ± 0.80 d,e,f | 20.25 ± 1.11 f,g | 14.04 ± 1.11 d,e | 5.15 ± 0.48 e,f | 12.44 ± 0.63 e,f |
| K1 | 14.65 ± 0.18 i,j,* | 13.89 ± 0.41 g | 22.38 ± 0.18 g,h,* | 20.18 ± 1.17 h,i | 19.84 ± 0.39 h,i | 14.47 ± 0.10 g,h |
| K2 | 13.82 ± 0.59 i | 11.75 ± 0.60 f | 23.12 ± 1.27 h,i | 17.51 ± 1.57 g | 20.20 ± 0.65 i | 13.45 ± 0.71 f,g |
| K3 | 18.31 ± 1.05 k,* | 17.83 ± 0.45 h | 32.58 ± 0.16 j | 23.93 ± 0.09 j | 26.92 ± 1.08 l | 22.24 ± 0.32 j |
| N1 | 5.06 ± 0.19 b | 3.23 ± 0.06 a | 3.39 ± 0.19 a | 7.45 ± 0.48 a,b | 4.90 ± 0.29 a,* | 5.71 ± 0.40 a |
| N2 | 2.73 ± 0.08 a,* | 2.14 ± 0.07 a | 6.30 ± 0.31 b,* | 6.28 ± 0.23 a | 7.97 ± 0.14 b | 5.58 ± 0.18 a |
| P1 | 9.24 ± 0.61 d,e,f | 7.28 ± 0.06 b | 15.58 ± 0.58 d,e | 11.77 ± 0.07 d | 23.85 ± 1.46 k | 11.78 ± 0.35 e |
| P2 | 11.83 ± 0.39 h | 7.67 ± 0.14 b | 15.00 ± 0.71 d,e,* | 14.87 ± 0.14 e,f | 13.48 ± 0.25 e,* | 12.30 ± 0.42 e,f |
| P3 | 6.80 ± 0.34 c,* | 6.57 ± 0.26 b | 13.67 ± 0.86 c,d,* | 12.00 ± 0.57 d | 9.55 ± 0.35 b,c,* | 9.71 ± 0.42 c |
| Bk | 9.37 ± 0.21 d,e,f | 10.47 ± 0.38 d,e | 20.72 ± 1.63 f,g | 13.99 ± 0.50 d,e | 20.97 ± 0.33 i,j | 13.33 ± 0.33 f,g |
| BE | 4.94 ± 0.03 b | 7.49 ± 0.43 b | 12.60 ± 0.47 c | 7.18 ± 0.20 a,b | 10.71 ± 0.40 c,d | 6.81 ± 0.47 a,b |
| PE | 5.77 ± 0.46 b,c,* | 7.36 ± 0.10 b | 6.96 ± 0.12 b | 11.53 ± 0.07 c,d | 5.96 ± 0.25 a | 15.62 ± 0.42 h,i |
| M | 14.56 ± 0.16 i,j | 12.09 ± 0.36 f | 24.90 ± 0.50 i | 15.58 ± 0.94 e,f,g | 22.24 ± 0.15 j,k | 11.80 ± 0.32 e |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carbas, B.; Machado, N.; Oppolzer, D.; Ferreira, L.; Queiroz, M.; Brites, C.; Rosa, E.A.; Barros, A.I. Nutrients, Antinutrients, Phenolic Composition, and Antioxidant Activity of Common Bean Cultivars and their Potential for Food Applications. Antioxidants 2020, 9, 186. https://doi.org/10.3390/antiox9020186
Carbas B, Machado N, Oppolzer D, Ferreira L, Queiroz M, Brites C, Rosa EA, Barros AI. Nutrients, Antinutrients, Phenolic Composition, and Antioxidant Activity of Common Bean Cultivars and their Potential for Food Applications. Antioxidants. 2020; 9(2):186. https://doi.org/10.3390/antiox9020186
Chicago/Turabian StyleCarbas, Bruna, Nelson Machado, David Oppolzer, Luís Ferreira, Marcelo Queiroz, Carla Brites, Eduardo AS Rosa, and Ana IRNA Barros. 2020. "Nutrients, Antinutrients, Phenolic Composition, and Antioxidant Activity of Common Bean Cultivars and their Potential for Food Applications" Antioxidants 9, no. 2: 186. https://doi.org/10.3390/antiox9020186
APA StyleCarbas, B., Machado, N., Oppolzer, D., Ferreira, L., Queiroz, M., Brites, C., Rosa, E. A., & Barros, A. I. (2020). Nutrients, Antinutrients, Phenolic Composition, and Antioxidant Activity of Common Bean Cultivars and their Potential for Food Applications. Antioxidants, 9(2), 186. https://doi.org/10.3390/antiox9020186