Allies or Enemies: The Role of Reactive Oxygen Species in Developmental Processes of Black Cottonwood (Populus trichocarpa)

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Microarray Analyses
2.3. Activity of Superoxide Dismutase (SOD) Assay
2.4. Localization of Superoxide
2.5. Localization of Hydrogen Peroxide
3. Results
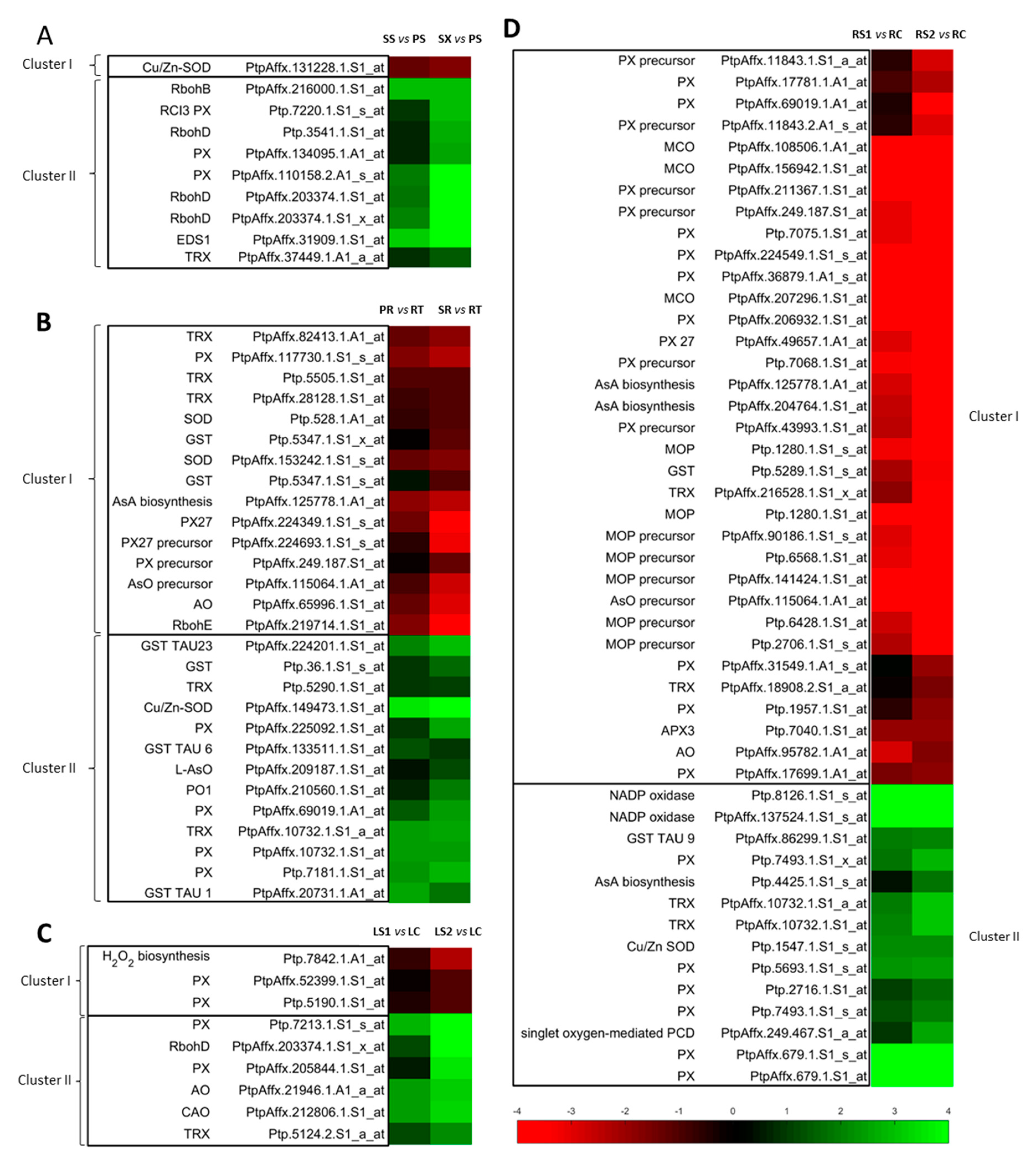
3.1. ROS and Oxidative Stress-Related Genes Are Overrepresented During the Primary/Secondary Growth of Stems and Pioneer Roots and Senescing Fine Roots and Leaves
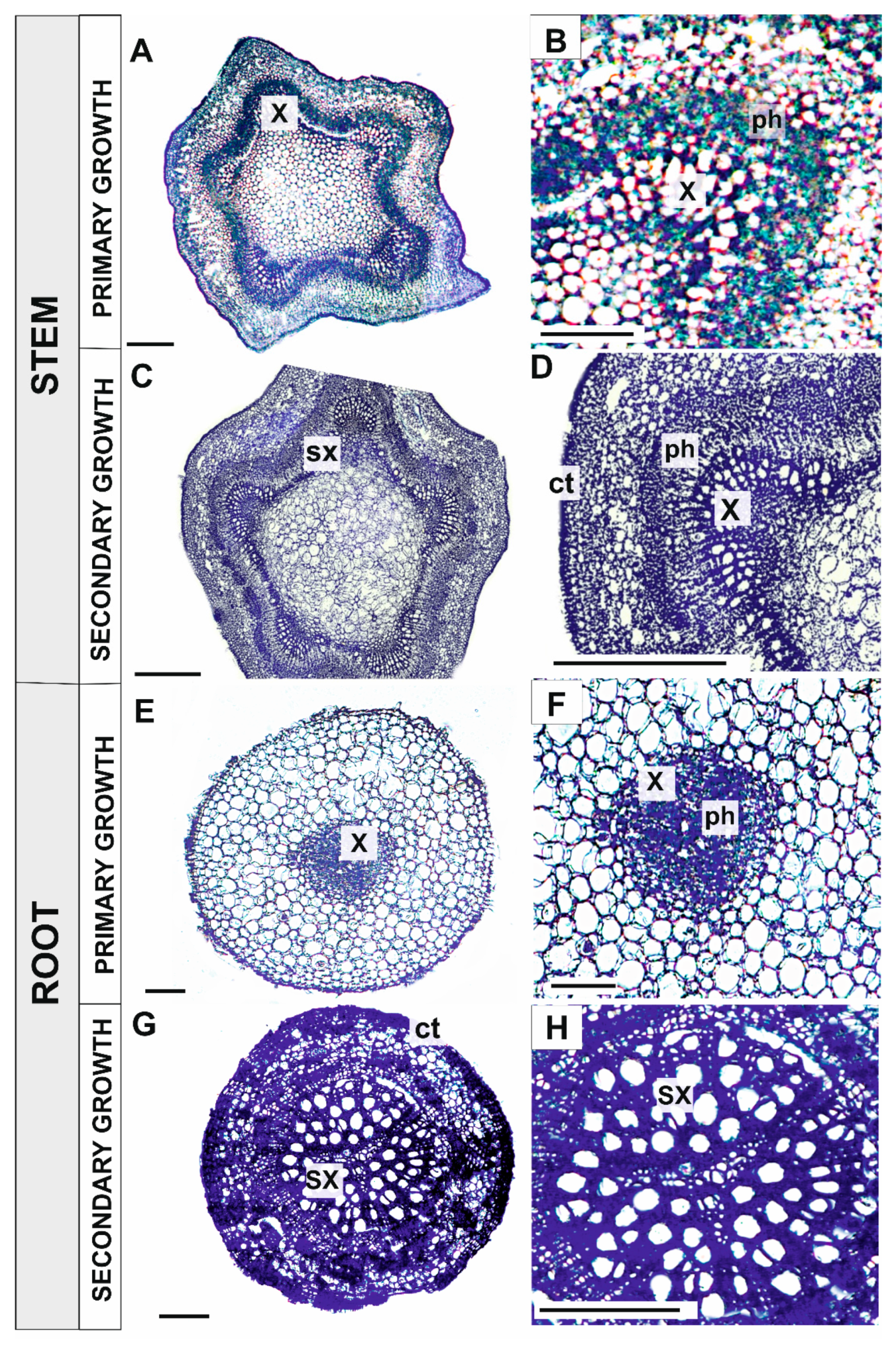
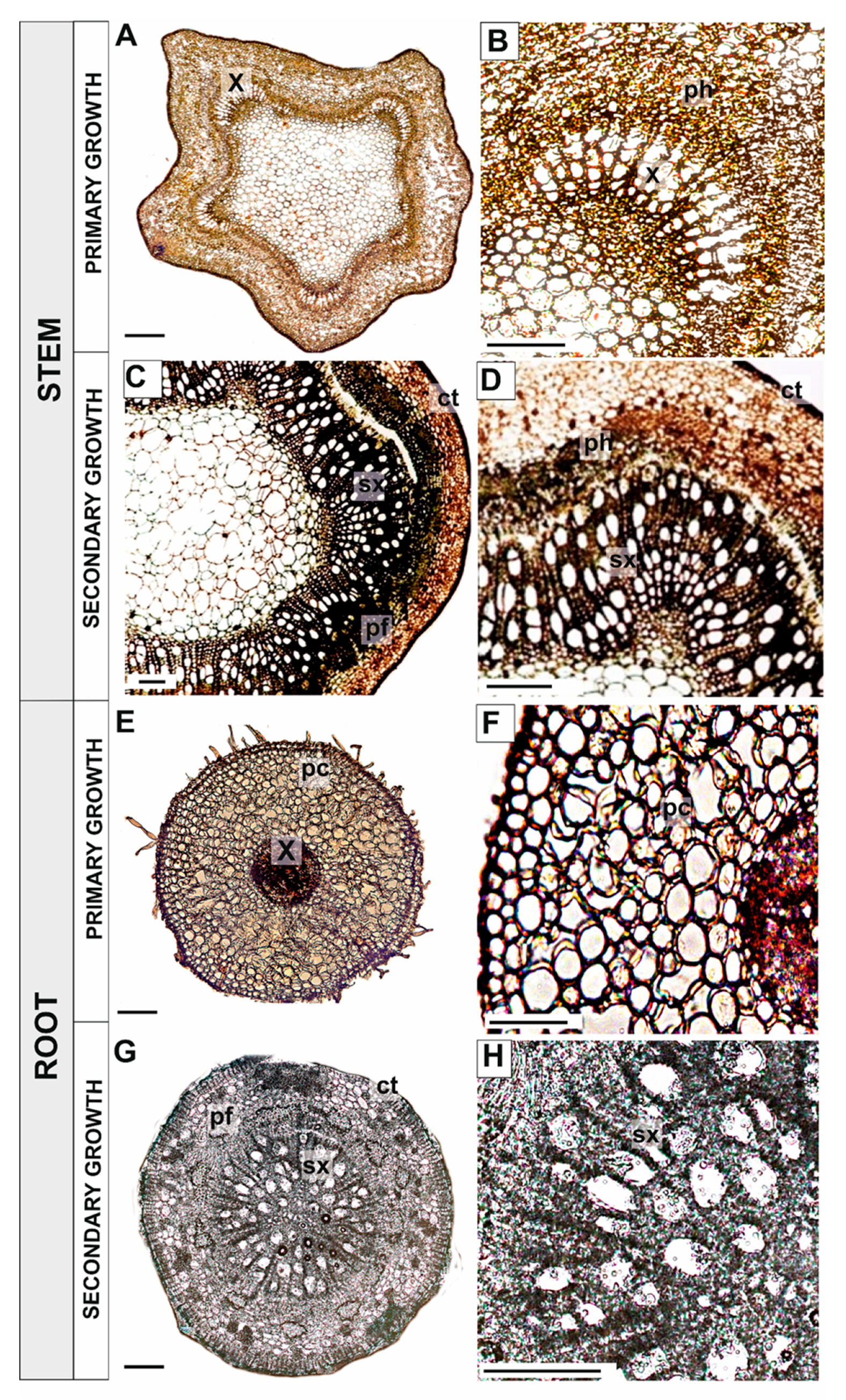
3.1.1. Primary and Secondary Growth of Plant Organs
Stems
Pioneer Roots
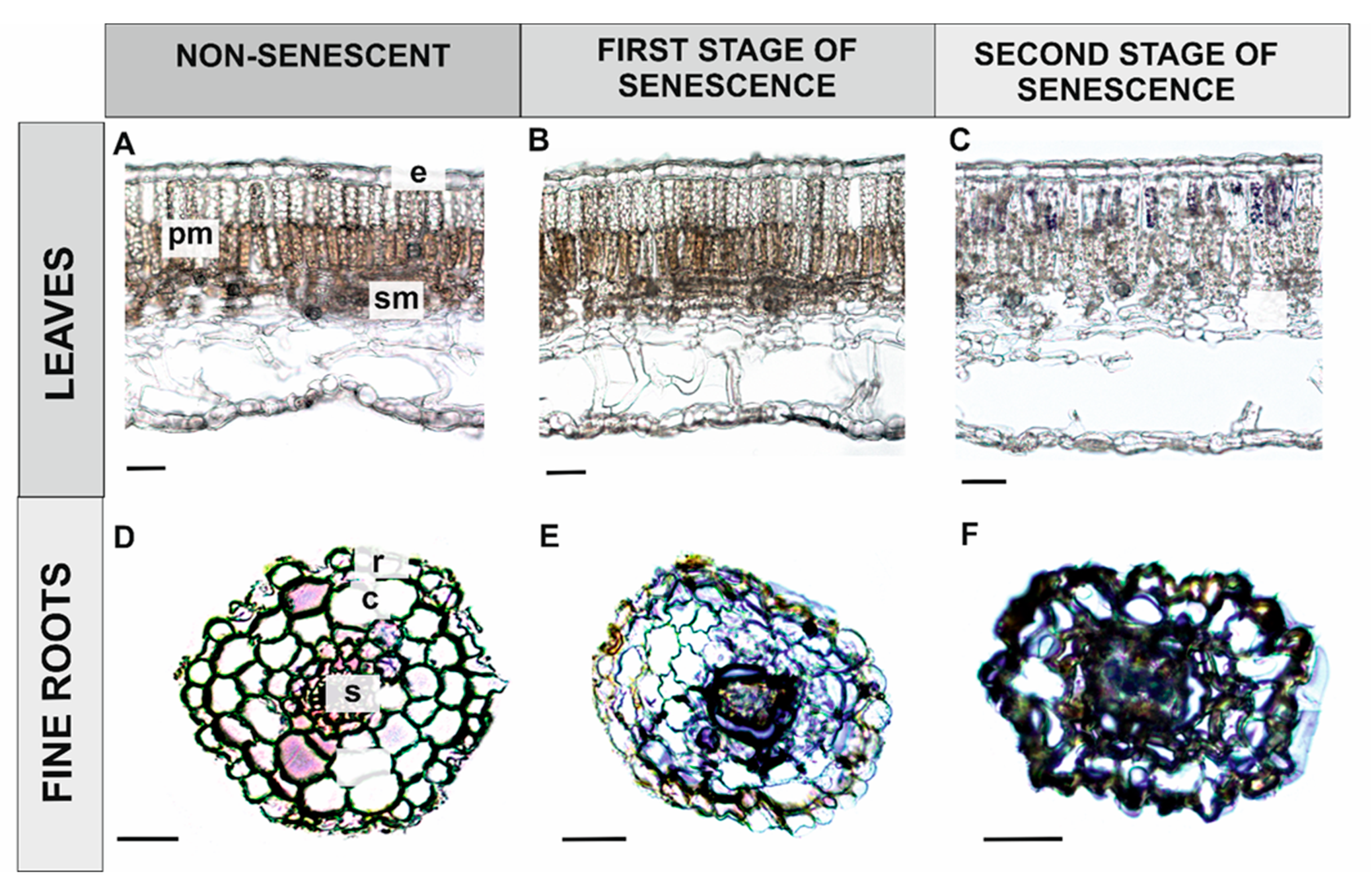
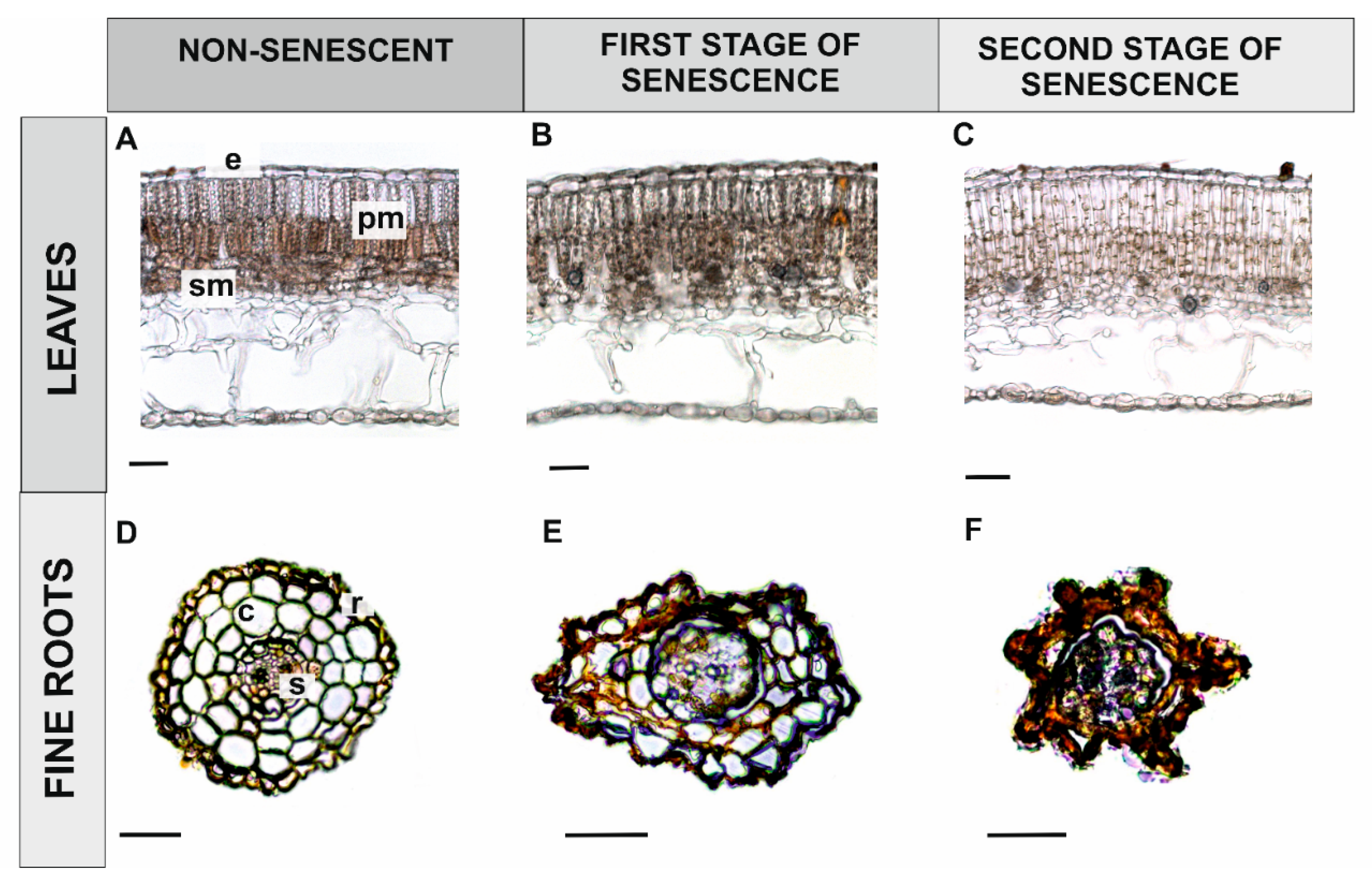
3.1.2. Senescing Plant Organs
Leaves
Fine Roots
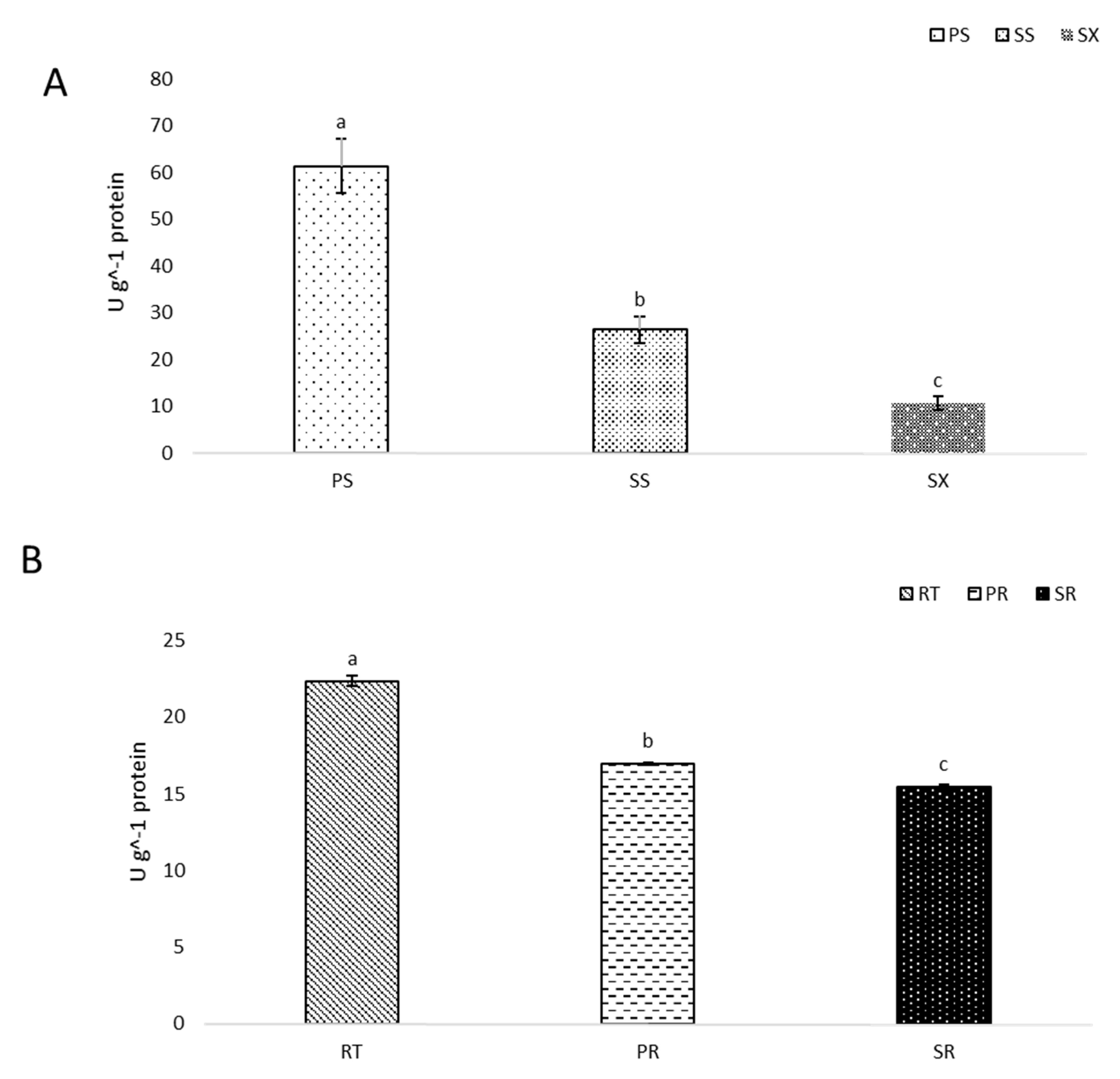
3.2. Superoxide Dismutase Activity Declines with Stem and Pioneer Root Development
3.3. Identification and Localization of Superoxide
3.4. Identification and Localization of Hydrogen Peroxide
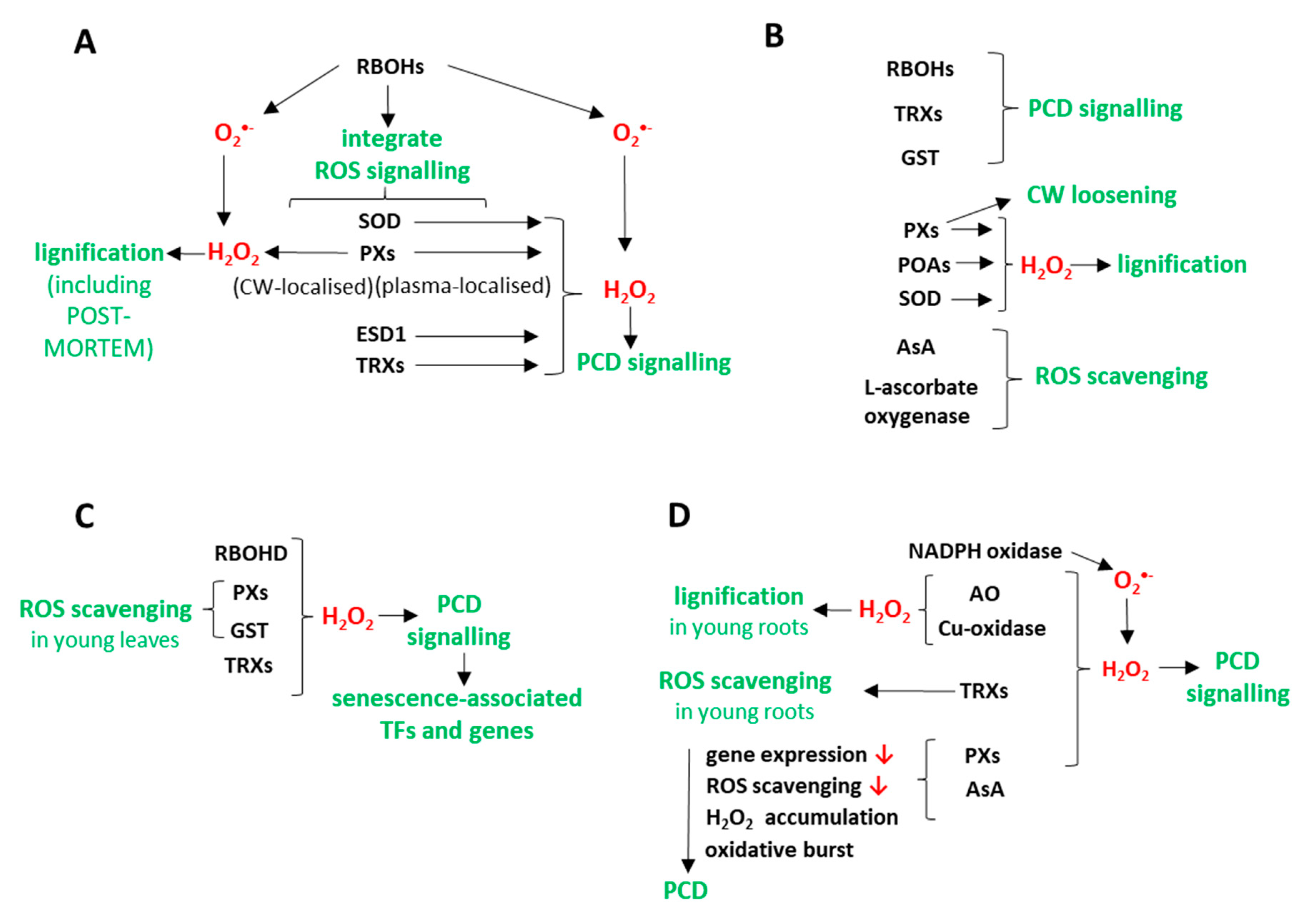
4. Discussion
4.1. ROS Are Involved in Xylem Development
4.2. ROS Play an Important Role in Developmental Process Involving PCD
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kangasjärvi, S.; Kangasjärvi, J. Towards understanding extracellular ROS sensory and signaling systems in plants. Adv. Bot. 2014. [Google Scholar] [CrossRef] [Green Version]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef] [PubMed]
- Mignolet-Spruyt, L.; Xu, E.; Idänheimo, N.; Hoeberichts, F.A.; Mühlenbock, P.; Brosché, M.; van Breusegem, F.; Kangasjärvi, J. Spreading the news: Subcellular and organellar reactive oxygen species production and signalling. J. Exp. Bot. 2016, 67, 3831–3844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Singh, S.; Parihar, P.; Mishra, R.K.; Tripathi, D.K.; Singh, V.P.; Chauhan, D.K.; Prasad, S.M. Reactive Oxygen Species (ROS): Beneficial companions of plants’ developmental processes. Front. Plant Sci. 2016, 7, 1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2016, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Serrano, I.; Romero-Puertas, M.C.; Sandalio, L.M.; Olmedilla, A. The role of reactive oxygen species and nitric oxide in programmed cell death associated with self-incompatibility. J. Exp. Bot. 2015, 66, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J. Interplay among nitric oxide and reactive oxygen species: A complex network determining cell survival or death. Plant Signal. Behav. 2007, 2, 544–547. [Google Scholar] [CrossRef] [Green Version]
- De Pinto, M.C.; Paradiso, A.; Leonetti, P.; De Gara, L. Hydrogen peroxide, nitric oxide and cytosolic ascorbate peroxidase at the crossroad between defence and cell death. Plant J. 2006, 48, 784–795. [Google Scholar] [CrossRef]
- Rantong, G.; Gunawardena, A. Programmed cell death: Genes involved in signaling, regulation, and execution in plants and animals. Botany 2015, 93, 193–210. [Google Scholar] [CrossRef]
- Bieker, S.; Riester, L.; Stahl, M.; Franzaring, J.; Zentgraf, U. Senescence-specific alteration of hydrogen peroxide levels in Arabidopsis thaliana and oilseed rape spring variety Brassica napus L. Cv. Mozart. J. Integr. Plant Biol. 2012, 54, 540–554. [Google Scholar] [CrossRef]
- Gadjev, I.; Stone, J.M.; Gechev, T.S. Programmed cell death in plants: New insights into redox regulation and the role of hydrogen peroxide. Int. Rev. Cell Mol. Biol. 2008, 270, 87–144. [Google Scholar] [CrossRef] [PubMed]
- Bagniewska-Zadworna, A.; Arasimowicz-Jelonek, M.; Smoliński, D.; Stelmasik, A. New insights into pioneer root xylem development: Evidence obtained from Populus trichocarpa plants grown under field conditions. Ann. Bot. 2014, 113, 1235–1247. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Jiang, F.; Sodmergen; Cui, K. Time-course of programmed cell death during leaf senescence in Eucommia ulmoides. J. Plant Res. 2003, 116, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Bagniewska-Zadworna, A.; Byczyk, J.; Eissentstat, D.M.; Oleksyn, J.; Zadworny, M. Avoiding transport bottlenecks in an expanding root system: Xylem vessel development in fibrous and pioneer roots under field conditions. Am. J. Bot. 2012, 99, 1417–1426. [Google Scholar] [CrossRef] [Green Version]
- He, R.; Drury, G.E.; Rotari, V.; Gordon, A.; Willer, M.; Farzaneh, T.; Woltering, E.J.; Gallois, P. Metacaspase-8 modulates Programmed Cell Death induced by ultraviolet light and H2O2 in Arabidopsis. J. Biol. Chem. 2008, 283, 774–783. [Google Scholar] [CrossRef] [Green Version]
- Desikan, R.; Reynolds, A.; Hancock, T.J.; Neill, J.S. Hairpin and hydrogen peroxide both initiate programmed cell death but have differential effects on defence gene expression in Arabidopsis suspension cultures. Biochem. J. 1998, 330, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hurst, R.D.; Hancock, J.T. Hydrogen peroxide and nitric oxide as signalling molecules in plants. J. Exp. Bot. 2002, 53, 1237–1247. [Google Scholar] [CrossRef]
- Wojciechowska, N.; Marzec-Schmidt, K.; Kalemba, E.M.; Zarzyńska-Nowak, A.; Jagodziński, A.M.; Bagniewska-Zadworna, A. Autophagy counteracts instantaneous cell death during seasonal senescence of the fine roots and leaves in Populus trichocarpa. BMC Plant Biol. 2018, 18, 260. [Google Scholar] [CrossRef]
- Prochazkova, D.; Sairam, R.K.; Srivastava, G.C.; Singh, D.V. Oxidative stress and antioxidant activity as the basis of senescence in maize leaves. Plant Sci. 2001, 161, 765–771. [Google Scholar] [CrossRef]
- Khanna-Chopra, R.; Nutan, K.K.; Pareek, A. Regulation of leaf senescence: Role of reactive oxygen species. In Plastid Development in Leaves during Growth and Senescence; Biswal, B., Krupinska, K., Biswal, U.C., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 393–416. [Google Scholar]
- Jajic, I.; Sarna, T.; Strzałka, K. Senescence, stress and reactive oxygen species. Plants 2015, 4, 393–411. [Google Scholar] [CrossRef] [Green Version]
- Khanna-Chopra, R. Leaf senescence and abiotic stresses share reactive oxygen species-mediated chloroplast degradation. Protoplasma 2012, 249, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Kurepa, J.; Smalle, J.; van Montagu, M.; Inzé, D. Oxidative stress tolerance and longevity in Arabidopsis: The late-flowering mutant gigantea is tolerant to paraquat. Plant J. 1998, 14, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G. Metabolic signalling in defence and stress: The central roles of soluble redox couples. Plant Cell Environ. 2006, 29, 409–425. [Google Scholar] [CrossRef] [PubMed]
- Woo, H.R.; Kim, J.H.; Nam, H.G.; Lim, P.O. The delayed leaf senescence mutants of Arabidopsis, ore1, ore3, and ore9 are tolerant to oxidative stress. Plant Cell Physiol. 2004, 45, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Van Doorn, W.G.; Woltering, E.J. Physiology and molecular biology of petal senescence. J. Exp. Bot. 2008, 59, 453–480. [Google Scholar] [CrossRef] [Green Version]
- Rogers, H.J. Is there an important role for reactive oxygen species and redox during floral senescence? Plant Cell Environ. 2012, 35, 217–233. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Guo, W.M.; Chen, S.M.; Han, L.; Li, Z.M. The role of N-lauroylethanolamine in the regulation of senescence of cut carnations (Dianthus caryophyllus). J. Plant Physiol. 2007, 164, 993–1001. [Google Scholar] [CrossRef]
- Panavas, T.; Rubinstein, B. Oxidative events during programmed cell death of daylily (Hemerocallis hybrid) petals. Plant Sci. 1998, 133, 125–138. [Google Scholar] [CrossRef]
- Rogers, H.; Munné-Bosch, S. Production and scavenging of reactive oxygen species and redox signaling during leaf and flower senescence: Similar but different. Plant Physiol. 2016, 171, 1560–1568. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowska, N.; Smugarzewska, I.; Marzec-Schmidt, K.; Zarzyńska-Nowak, A.; Bagniewska-Zadworna, A. Occurrence of autophagy during pioneer root and stem development in Populus trichocarpa. Planta 2019, 250, 1789–1801. [Google Scholar] [CrossRef] [Green Version]
- Marzec-Schmidt, K.; Ludwików, A.; Wojciechowska, N.; Kasprowicz-Maluśki, A.; Mucha, J.; Bagniewska-Zadworna, A. Xylem cell wall formation in pioneer roots and stems of Populus trichocarpa (Torr. & Gray). Front. Plant Sci. 2019, 10, 1419. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Doke, N. Generation of superoxide anion by potato tuber protoplasts during the hypersensitive response to hyphal wall components of Phytophthora infestans and specific inhibition of the reaction by suppressors of hypersensitivity. Physiol. Plant Pathol. 1983, 23, 359–367. [Google Scholar] [CrossRef]
- Ogawa, K.; Kanematsu, S.; Asada, K. Intra- and extra-cellular localization of “cytosolic” CuZn-superoxide dismutase in spinach leaf and hypocotyl. Plant Cell Physiol. 1996, 37, 790–799. [Google Scholar] [CrossRef] [Green Version]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley—Powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37. [Google Scholar] [CrossRef] [Green Version]
- Ros Barceló, A. Xylem parenchyma cells deliver the H2O2 necessary for lignification in differentiating xylem vessels. Planta 2005, 220, 747–756. [Google Scholar] [CrossRef]
- Podgórska, A.; Burian, M.; Szal, B. Extra-cellular but extra-ordinarily important for cells: Apoplastic reactive oxygen species metabolism. Front. Plant Sci. 2017, 8, 1353. [Google Scholar] [CrossRef] [Green Version]
- Rustérucci, C.; Aviv, D.H.; Holt, B.F.; Dangl, J.L.; Parker, J.E. The disease resistance signalling components EDS1 and PAD4 are essential regulators of the cell death pathway controlled by LSD1 in Arabidopsis. Plant Cell 2001, 13, 2211–2224. [Google Scholar] [CrossRef] [Green Version]
- Marjamaa, K.; Kukkola, E.M.; Fagerstedt, K.V. The role of xylem class III peroxidases in lignification. J. Exp. Bot. 2009, 60, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Francoz, E.; Ranocha, P.; Nguyen-Kim, H.; Jamet, E.; Burlat, V.; Dunand, C. Roles of cel wall peroxidases in plant development. Phytochemistry 2015, 112, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Yoda, H.; Yamaguchi, Y.; Sano, H. Induction of hypersensitive cell death by hydrogen peroxide produced through polyamine degradation in tobacco plants. Plant Physiol. 2003, 132, 1973–1981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnoff, N. The function and metabolism of ascorbic acid in plants. Ann. Bot. 1996, 78, 661–669. [Google Scholar] [CrossRef] [Green Version]
- Meyer, R.C.; Goldsbrough, P.B.; Woodson, W.R. An ethylene-responsive flower senescence-related gene from carnation encodes a protein homologous to glutathione S-transferase. Plant Mol. Biol. 1991, 17, 277–281. [Google Scholar] [CrossRef]
- Dauphinee, A.N.; Fletcher, J.I.; Denbigh, G.L.; Lacroix, C.R.; Gunawardena, A.H.L.A.N. Remodelling of lace plant leaves: Antioxidants and ROS are key regulators of programmed cell death. Planta 2017, 246, 133–147. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Wojciechowska, N.; Marzec-Schmidt, K.; Ludwików, A.; Kalemba, E.; Bagniewska-Zadworna, A. Seasonal senescence of leaves and roots of Populus trichocarpa—Is the scenario the same or different? Tree Physiol. 2020, in press. [Google Scholar] [CrossRef]
- Karlsson, M.; Melzer, M.; Prokhorenko, I.; Johansson, T.; Wingsle, G. Hydrogen peroxide and expression of hipI-superoxide dismutase are associated with the development of secondary cell walls in Zinnia elegans. J. Exp. Bot. 2005, 56, 2085–2093. [Google Scholar] [CrossRef] [Green Version]
- Kärkönen, A.; Kuchitsu, K. Reactive oxygen species in cell wall metabolism and development. Phytochemistry 2015, 112, 22–32. [Google Scholar] [CrossRef]
- Coll, N.S.; Epple, P.; Dangl, J.L. Programmed cell death in the plant immune system. Cell Death Differ. 2011, 18, 1247–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willems, P.; Mhamdi, A.; Stael, S.; Storme, V.; Kerchev, P.; Noctor, G.; Gevaert, K.; van Breusegem, F. The ROS wheel: Refining ROS transcriptional footprints in Arabidopsis. Plant Physiol. 2016, 171, 1720–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, M.J.; Choi, W.G.; Gilroy, S.; Morris, R. A ROS-assisted calcium wave dependent on AtRBOHD and TPC1 propagates the systemic response to salt stress in Arabidopsis roots. Plant Physiol. 2016, 17, 1771–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilroy, S.; Białasek, M.; Suzuki, N.; Górecka, M.; Devireddy, A.R.; Karpinski, S.; Mittler, R. ROS, calcium, and electric signals: Key mediators of rapid systemic signalling in plants. Plant Physiol. 2016, 171, 1606–1615. [Google Scholar] [CrossRef]
- Ogawa, K.; Kanematsu, S.; Asada, K. Generation of superoxide anion and localization of CuZn-superoxide dismutase in the vascular tissue of spinach hypocotyls: Their association with lignification. Plant Cell Physiol. 1997, 38, 1118–1126. [Google Scholar] [CrossRef] [Green Version]
- Schweikert, C.; Liszkay, A.; Schopfer, P. Scission of polysaccharides by peroxidase-generated hydroxyl radicals. Phytochemistry 2000, 53, 565–570. [Google Scholar] [CrossRef]
- Sakamoto, M.; Munemura, I.; Tomita, R.; Kobayashi, K. Reactive oxygen species in leaf abscission signalling. Plant Signal. Behav. 2008, 3, 1014–1015. [Google Scholar] [CrossRef] [Green Version]
- Schopfer, P. Hydroxyl radical-induced cell-wall loosening in vitro and in vivo: Implications for the control of elongation growth. Plant J. 2001, 28, 679–688. [Google Scholar] [CrossRef] [Green Version]
- Hohl, M.; Greiner, H.; Schopfer, P. The cryptic growth response of maize coleoptiles and its relationship to H2O2 dependent cell wall stiffening. Physiol. Plant. 1995, 94, 491–498. [Google Scholar] [CrossRef]
- Schopfer, P. Hydrogen peroxide-mediated cell-wall stiffening in vitro in maize coleoptiles. Planta 1996, 199, 43–49. [Google Scholar] [CrossRef]
- Novo-Uzal, E.; Fernández-Pérez, F.; Herrero, J.; Gutiérrez, J.; Gómez-Ros, L.V.; Ángeles Bernal, M.; Díaz, J.; Cuello, J.; Pomar, F.; Pedreño, M.A. From Zinnia to Arabidopsis: Approaching the involvement of peroxidases in lignification. J. Exp. Bot. 2013, 64, 3499–3518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Mansouri, I.; Mercado, J.A.; Santiago-Domenech, N.; Pliego-Alfaro, F.; Valpuesta, V.; Quesada, M.A. Biochemical and phenotypical characterization of transgenic tomato plants overexpressing a basic peroxidase. Physiol. Plant. 1999, 106, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Kajita, S.; Kawai, S.; Katayama, Y.; Morohoshi, N. Down-regulation of an anionic peroxidase in transgenic aspen and its effect on lignin characteristics. J. Plant Res. 2003, 116, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Herrero, J.; Carrasco, A.E.; Zapata, J.M. Arabidopsis thaliana peroxidases in lignin biosynthesis: In silico promoter analysis and hormonal regulation. Plant Physiol. Biochem. 2014, 80, 192–202. [Google Scholar] [CrossRef]
- Llorente, F.; Lopez-Cobollo, R.M.; Catala, R.; Martinez-Zapater, J.M.; Salinas, J. A novel cold-inducible gene from Arabidopsis, RCI3, encodes a peroxidase that constitutes a component for stress tolerance. Plant J. 2002, 32, 13–24. [Google Scholar] [CrossRef]
- Tisi, A.; Federico, R.; Moreno, S.; Lucretti, S.; Moschou, P.N.; Roubelakis-Angelakis, K.A.; Angelini, R.; Cona, A. Perturbation of polyamine catabolism can strongly affect root development and xylem differentiation. Plant Physiol. 2011, 157, 200–215. [Google Scholar] [CrossRef] [Green Version]
- Møller, S.G.; McPherson, M.J. Developmental expression and biochemical analysis of the Arabidopsis atao1 gene encoding an H2O2-generating diamine oxidase. Plant J. 1998, 13, 781–791. [Google Scholar] [CrossRef]
- Pignocchi, C.; Foyer, C.H. Apoplastic ascorbate metabolism and its role in the regulation of cell signalling. Curr. Opin. Plant Biol. 2003, 6, 379–389. [Google Scholar] [CrossRef]
- Nühse, T.S.; Bottrill, A.R.; Jones, A.M.; Peck, S.C. Quantitative phosphoproteomic analysis of plasma membrane proteins reveals regulatory mechanisms of plant innate immune responses. Plant J. 2007, 51, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Van Aken, O.; van Breuseyem, F. Licensed to kill: Mitochondria, chloroplasts, and cell death. Trends Plant Sci. 2015, 20, 754–766. [Google Scholar] [CrossRef]
- Sakharov, I.Y.; Vesga, M.K.; Galaev, I.Y.; Sakharova, I.V.; Pletjushkina, O.Y. Peroxidase from leaves of royal palm tree Roystonea regia: Purification and some properties. Plant Sci. 2001, 161, 853–860. [Google Scholar] [CrossRef]
- Damdimopoulos, A.E.; Miranda-Vizuete, A.; Pelto-Huikko, M.; Gustafsson, J.A.; Spyrou, G. Human mitochondrial thioredoxin. Involvement in mitochondrial membrane potential and cell death. J. Biol. Chem. 2002, 277, 33249–33257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, J.M.; Zhang, H.; Jones, D.P. Mitochondrial thioredoxin-2 has a key role in determining tumour necrosis factor-{alpha}-induced reactive oxygen species generation, NF-{kappa}B activation, and apoptosis. Toxicol. Sci. 2006, 91, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Smart, C.M.; Hosken, S.E.; Thomas, H.; Greaves, J.A.; Blair, B.G.; Schuch, W. The timing of maize leaf senescence and characterization of senescence-related cDNAs. Physiol. Plant. 1995, 93, 673–682. [Google Scholar] [CrossRef]
- Op den Camp, R.G.L.; Przybyla, D.; Ochsenbein, C.; Laloi, C.; Kim, C.; Danon, A.; Wagner, D.; Hideg, É.; Göbel, C.; Feussner, I.; et al. Rapid induction of distinct stress responses after the release of singlet oxygen in arabidopsis. Plant Cell 2003, 15, 2320–2332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laloi, C.; Stachowiak, M.; Pers-Kamczyc, E.; Warzych, E.; Murgia, I.; Apel, K. Cross-talk between singlet oxygen- and hydrogen peroxide-dependent signaling of stress responses in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2007, 104, 672–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.; Meskauskiene, R.; Apel, K.; Laloi, C. No single way to understand singlet oxygen signalling in plants. EMBO Rep. 2008, 9, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Munné-Bosch, S.; Jubany-Marí, T.; Alegre, L. Drought-induced senescence is characterized by a loss of antioxidant defences in chloroplasts. Plant Cell Environ. 2001, 24, 1319–1327. [Google Scholar] [CrossRef]
- Berger, S.; Weichert, H.; Porzel, A.; Wasternack, C.; Kühn, H.; Feussner, I. Enzymatic and non-enzymatic lipid peroxidation in leaf development. Biochim. Biophys. Acta 2001, 1533, 266–276. [Google Scholar] [CrossRef]
- Zhang, M.P.; Zhang, C.J.; Yu, G.H.; Jiang, Y.Z.; Strasser, R.J.; Yuan, Z.Y.; Yang, X.S.; Chen, G.X. Changes in chloroplast ultrastructure, fatty acid components of thylakoid membrane and chlorophyll a fluorescence transient in flag leaves of a super-high-yield hybrid rice and its parents during the reproductive stage. J. Plant Physiol. 2010, 167, 277–285. [Google Scholar] [CrossRef]
- Zimmermann, P.; Heinlein, C.; Orendi, G.; Zentgraf, U. Senescence-specific regulation of catalases in Arabidopsis thaliana (L.) heynh. Plant Cell Environ. 2006, 29, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Dat, J.; Vandenabeele, S.; Vranová, E.; van Montagu, M.; Inzé, D.; van Breusegem, F. Dual action of the active oxygen species during plant stress responses. Cell. Mol. Life Sci. 2000, 57, 779–795. [Google Scholar] [CrossRef] [PubMed]
- Wright, H.; van Doorn, W.G.; Gunawardena, A.H.L.A.N. In vivo study of developmental cell death using the lace plant (Aponogeton madagascariensis; Aponogetonaceae) leaf model system. Am. J. Bot. 2009, 96, 856–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, H.; Yoshimoto, K.; Izumi, M.; Reisen, D.; Yano, Y.; Makino, A.; Ohsumi, Y.; Hanson, M.R.; Mae, T. Mobilization of rubisco and stroma-localized fluorescent proteins of chloroplasts to the vacuole by an ATG gene-dependent autophagic process. Plant Physiol. 2008, 148, 142–155. [Google Scholar] [CrossRef] [Green Version]
- Wada, S.; Ishida, H.; Izumi, M.; Yoshimoto, K.; Ohsumi, Y.; Mae, T.; Makino, A. Autophagy plays a role in chloroplast degradation during senescence in individually darkened leaves. Plant Physiol. 2009, 149, 885–893. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Abbreviation | Characteristic Features |
|---|---|---|
| Stem | PS | apical meristem with primary growth |
| SS | secondary growth | |
| SX | isolated secondary xylem | |
| Pioneer Roots | RT | root tip with apical meristem |
| PR | primary growth | |
| SR | secondary growth | |
| Leaves | LC | control green leaves without senescence symptoms |
| LS1 | first stage of senescence: yellowing leaves in which chlorophyll level had decreased by approximately 40% | |
| LS2 | second stage of senescence: yellow leaves in which chlorophyll level had decreased by approximately 60% | |
| Fine Roots | RC | control white roots without senescence symptoms |
| RS1 | first stage of senescence: roots which had changed in colour from white to brown | |
| RS2 | second stage of senescence: roots which had changed in colour from brown to dark brown or almost black, shrinkage was also visible in most fine roots |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marzec-Schmidt, K.; Wojciechowska, N.; Nemeczek, K.; Ludwików, A.; Mucha, J.; Bagniewska-Zadworna, A. Allies or Enemies: The Role of Reactive Oxygen Species in Developmental Processes of Black Cottonwood (Populus trichocarpa). Antioxidants 2020, 9, 199. https://doi.org/10.3390/antiox9030199
Marzec-Schmidt K, Wojciechowska N, Nemeczek K, Ludwików A, Mucha J, Bagniewska-Zadworna A. Allies or Enemies: The Role of Reactive Oxygen Species in Developmental Processes of Black Cottonwood (Populus trichocarpa). Antioxidants. 2020; 9(3):199. https://doi.org/10.3390/antiox9030199
Chicago/Turabian StyleMarzec-Schmidt, Katarzyna, Natalia Wojciechowska, Klaudia Nemeczek, Agnieszka Ludwików, Joanna Mucha, and Agnieszka Bagniewska-Zadworna. 2020. "Allies or Enemies: The Role of Reactive Oxygen Species in Developmental Processes of Black Cottonwood (Populus trichocarpa)" Antioxidants 9, no. 3: 199. https://doi.org/10.3390/antiox9030199
APA StyleMarzec-Schmidt, K., Wojciechowska, N., Nemeczek, K., Ludwików, A., Mucha, J., & Bagniewska-Zadworna, A. (2020). Allies or Enemies: The Role of Reactive Oxygen Species in Developmental Processes of Black Cottonwood (Populus trichocarpa). Antioxidants, 9(3), 199. https://doi.org/10.3390/antiox9030199