Effects of A2E-Induced Oxidative Stress on Retinal Epithelial Cells: New Insights on Differential Gene Response and Retinal Dystrophies

 ,
, 



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. MTT Assay
2.3. Total RNA Sequencing
2.4. Quality Validation and Read Mapping
2.5. Gene Expression and Statistical Analysis
2.6. DE, DAS and DTU Analysis
2.7. Gene-Enrichment and Functional Pathway Analysis
2.8. Selection of Single-Pathway “Master genes” and Selection of Retinitis Pigmentosa Candidate Genes by ToppGene Prioritization
2.9. Data Validation by qRT-PCR.
3. Results
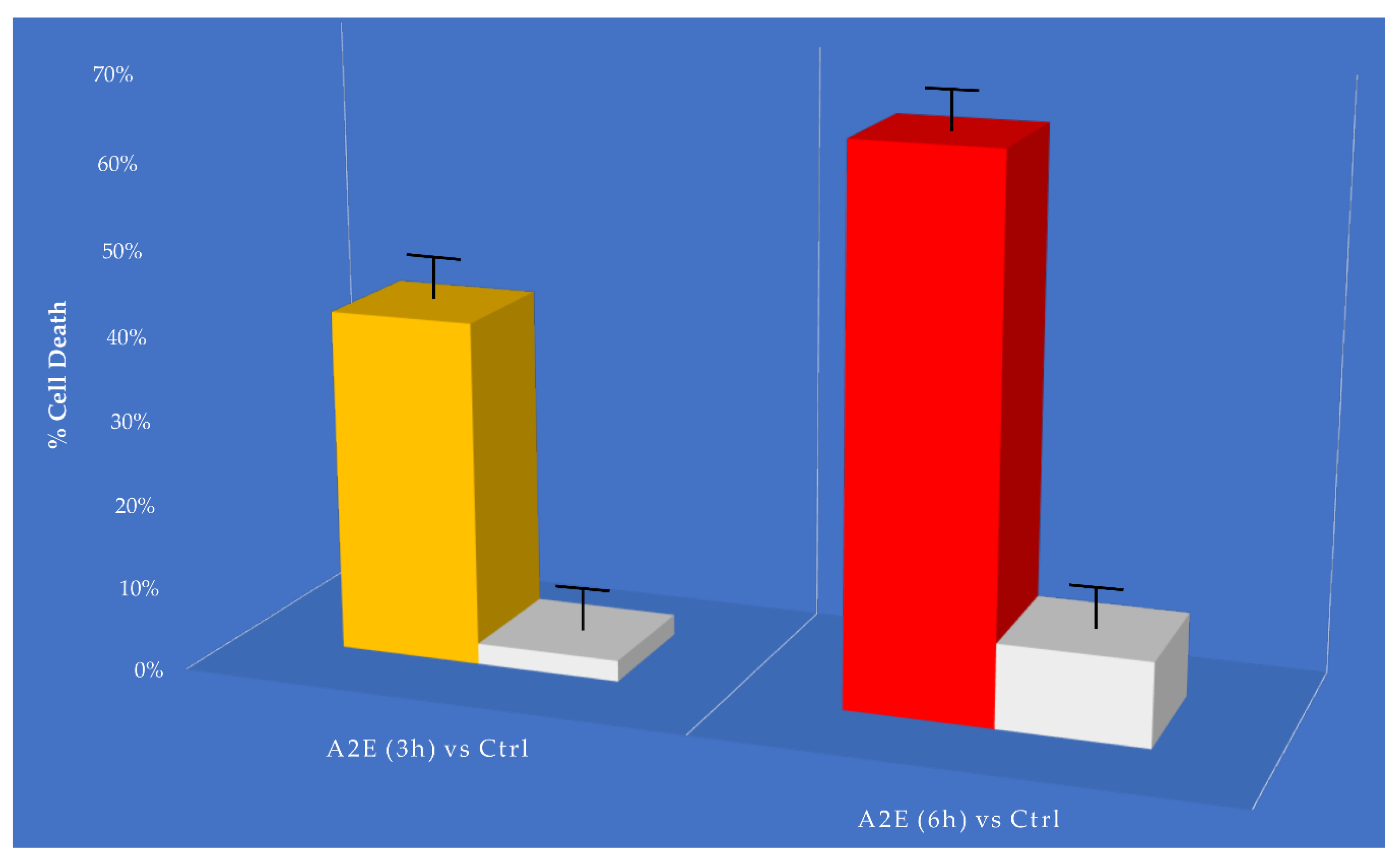
3.1. MTT Cell Viability Assay Results
3.2. Sequencing Analysis and Mapping Statistics
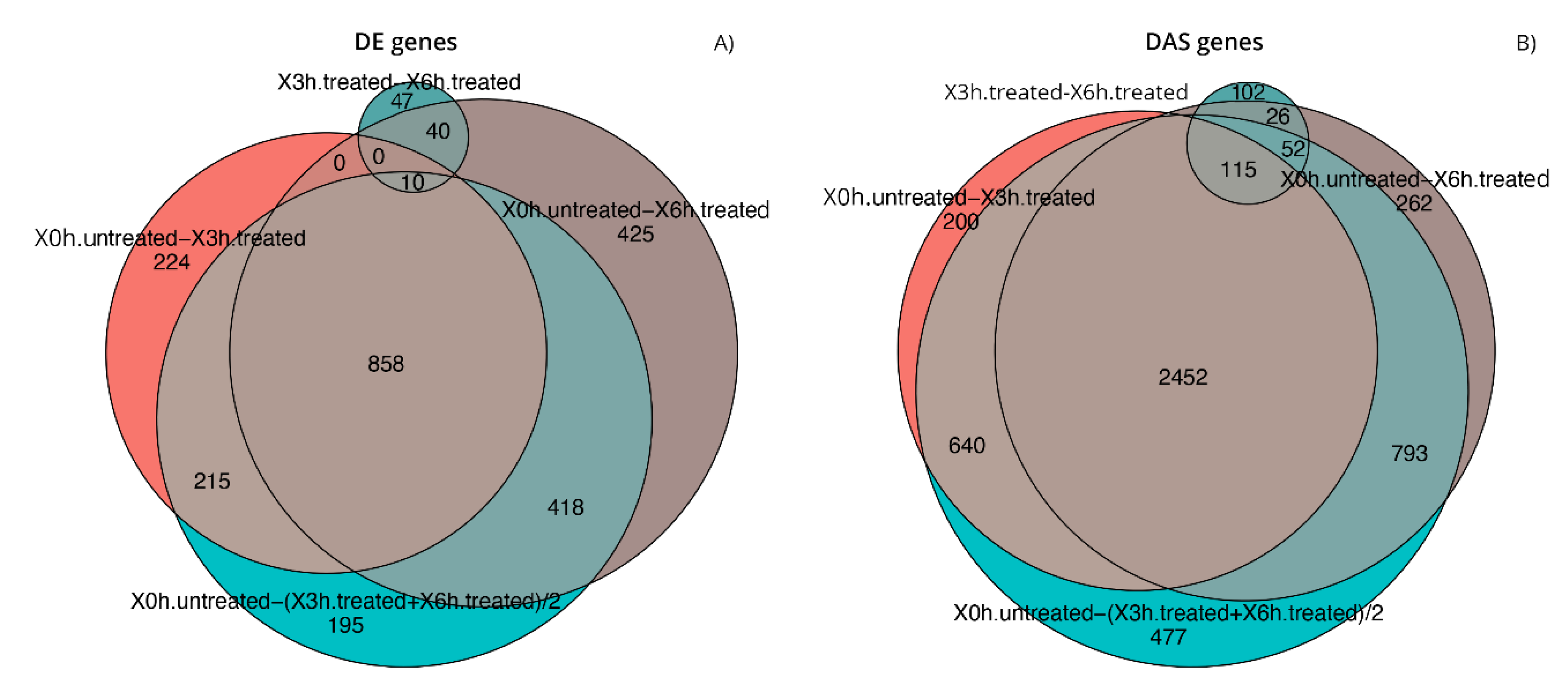
3.3. Analysis of Gene Expression Profile of RPE Cells
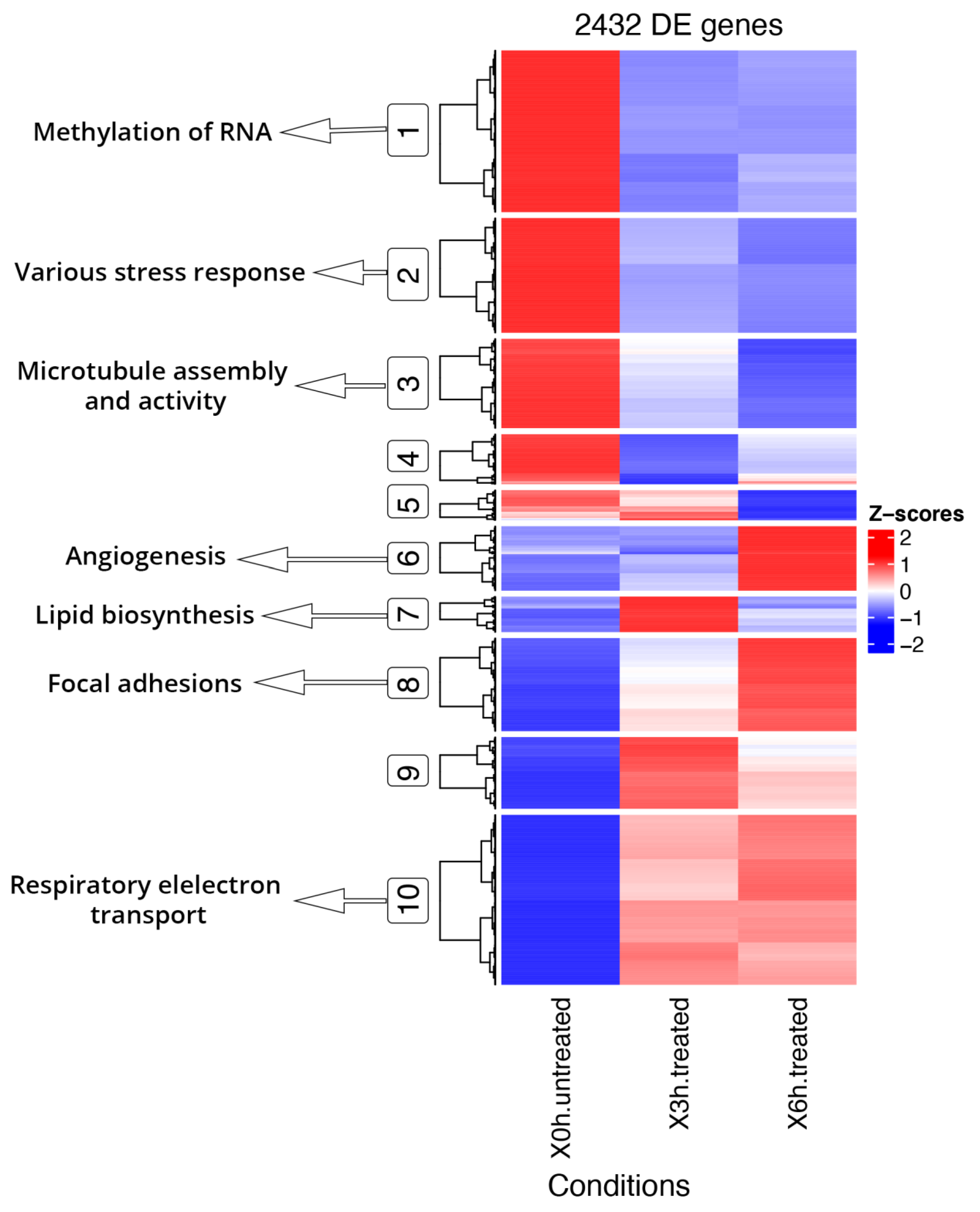
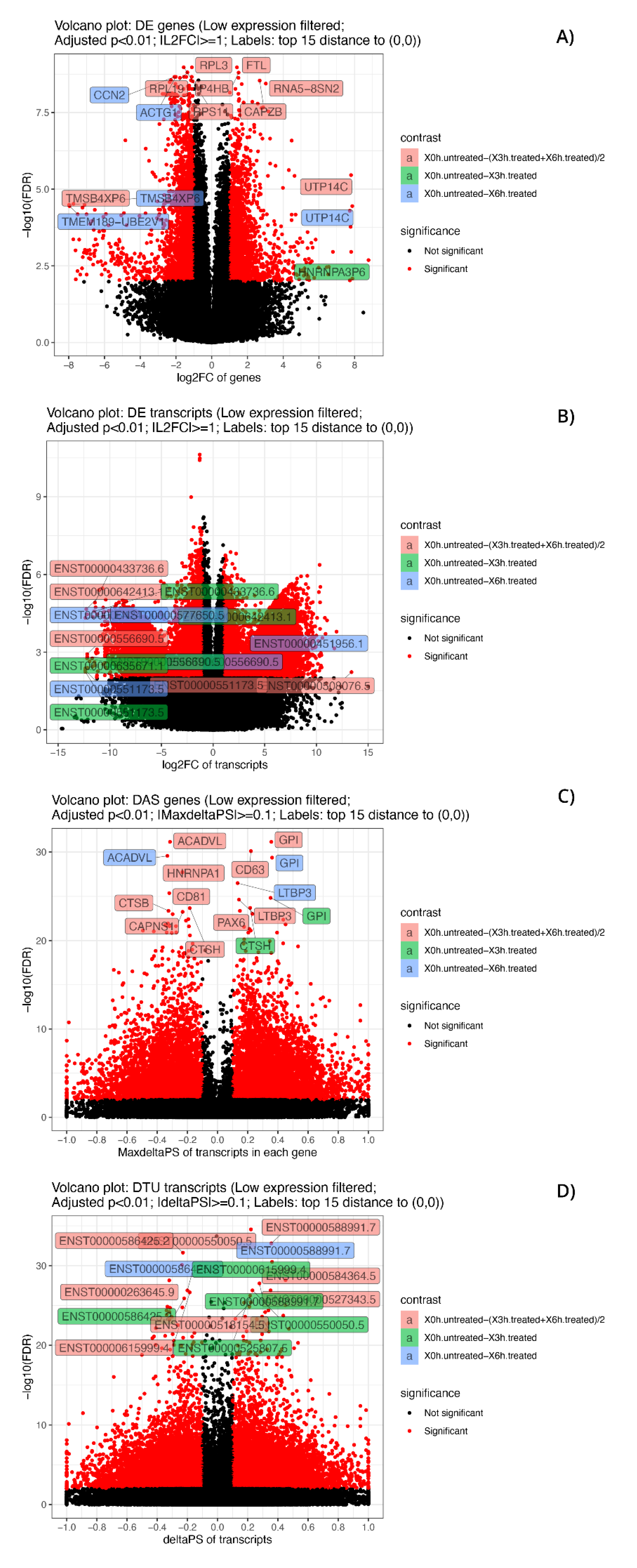
3.4. DE and DAS Genes Highlighted Different Functionality Patterns
3.5. Early Cellular Response to Induced Stress Mainly Involves Pre-mRNA Splicing and Glycolysis-Related DE and DAS Genes
3.6. Late RPE Cell Response to A2E Treatment Could Impair Bioenergetic Specific Reactions, Extracellular Matrix Integrity and Neurotransmission-Related DE and DAS Genes
3.7. The Transcriptome Comparison between Untreated (Time Zero) and Treated (3 h + 6 h) Rpe Cells Revealed the Possible Impairment of Retinal Cells Crosstalk and Synapses, Leading to Rescue or Cell Death
3.8. The Most Significant DAS Genes Represented the Main Retinal Dystrophy Candidate Genes
3.9. qRT-PCR Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [Green Version]
- Donato, L.; Scimone, C.; Nicocia, G.; D’Angelo, R.; Sidoti, A. Role of oxidative stress in Retinitis pigmentosa: New involved pathways by an RNA-Seq analysis. Cell Cycle 2019, 18, 84–104. [Google Scholar] [CrossRef] [Green Version]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The impact of oxidative stress and inflammation on RPE degeneration in non-neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.A. Age-related macular degeneration. Workplace Health Saf. 2014, 62, 352. [Google Scholar] [CrossRef] [PubMed]
- Gheorghe, A.; Mahdi, L.; Musat, O. Age-Related Macular Degeneration. Rom. J. Ophthalmol. 2015, 59, 74–77. [Google Scholar] [PubMed]
- Lim, L.S.; Mitchell, P.; Seddon, J.M.; Holz, F.G.; Wong, T.Y. Age-related macular degeneration. Lancet 2012, 379, 1728–1738. [Google Scholar] [CrossRef]
- Kivinen, N.; Koskela, A.; Kauppinen, A.; Kaarniranta, K. Pathogenesis of age-related macular degeneration—dialogue between autophagy and inflammasomes. Duodecim 2017, 133, 641–646. [Google Scholar] [PubMed]
- Xu, B.; Zhang, H.; Zhu, M.; Le, Y.Z. Critical Role of Trophic Factors in Protecting Muller Glia: Implications to Neuroprotection in Age-Related Macular Degeneration, Diabetic Retinopathy, and Anti-VEGF Therapies. Adv. Exp. Med. Biol. 2019, 1185, 469–473. [Google Scholar] [CrossRef]
- Natarajan, S. Decoding retinitis pigmentosa. Indian J. Ophthalmol. 2013, 61, 91–94. [Google Scholar] [CrossRef]
- Verbakel, S.K.; van Huet, R.A.C.; Boon, C.J.F.; den Hollander, A.I.; Collin, R.W.J.; Klaver, C.C.W.; Hoyng, C.B.; Roepman, R.; Klevering, B.J. Non-syndromic retinitis pigmentosa. Prog. Retin. Eye Res. 2018, 66, 157–186. [Google Scholar] [CrossRef]
- Fahim, A. Retinitis pigmentosa: Recent advances and future directions in diagnosis and management. Curr. Opin. Pediatr. 2018, 30, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Agbaga, M.P.; Merriman, D.K.; Brush, R.S.; Lydic, T.A.; Conley, S.M.; Naash, M.I.; Jackson, S.; Woods, A.S.; Reid, G.E.; Busik, J.V.; et al. Differential composition of DHA and very-long-chain PUFAs in rod and cone photoreceptors. J. Lipid Res. 2018, 59, 1586–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birtel, J.; Gliem, M.; Oishi, A.; Muller, P.L.; Herrmann, P.; Holz, F.G.; Mangold, E.; Knapp, M.; Bolz, H.J.; Charbel Issa, P. Genetic testing in patients with retinitis pigmentosa: Features of unsolved cases. Clin. Exp. Ophthalmol. 2019, 47, 779–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campochiaro, P.A.; Mir, T.A. The mechanism of cone cell death in Retinitis Pigmentosa. Prog. Retin. Eye Res. 2018, 62, 24–37. [Google Scholar] [CrossRef]
- Audo, I.; Mohand-Said, S.; Boulanger-Scemama, E.; Zanlonghi, X.; Condroyer, C.; Demontant, V.; Boyard, F.; Antonio, A.; Mejecase, C.; El Shamieh, S.; et al. MERTK mutation update in inherited retinal diseases. Hum. Mutat. 2018, 39, 887–913. [Google Scholar] [CrossRef]
- Scimone, C.; Donato, L.; Esposito, T.; Rinaldi, C.; D’Angelo, R.; Sidoti, A. A novel RLBP1 gene geographical area-related mutation present in a young patient with retinitis punctata albescens. Hum. Genom. 2017, 11, 18. [Google Scholar] [CrossRef]
- Miraldi Utz, V.; Coussa, R.G.; Antaki, F.; Traboulsi, E.I. Gene therapy for RPE65-related retinal disease. Ophthalmic Genet. 2018, 39, 671–677. [Google Scholar] [CrossRef]
- Sparrow, J.R.; Hicks, D.; Hamel, C.P. The retinal pigment epithelium in health and disease. Curr. Mol. Med. 2010, 10, 802–823. [Google Scholar] [CrossRef]
- Nowak, J.Z. Oxidative stress, polyunsaturated fatty acids-derived oxidation products and bisretinoids as potential inducers of CNS diseases: Focus on age-related macular degeneration. Pharmacol. Rep. 2013, 65, 288–304. [Google Scholar] [CrossRef]
- Donato, L.; Scimone, C.; Rinaldi, C.; D’Angelo, R.; Sidoti, A. Non-coding RNAome of RPE cells under oxidative stress suggests unknown regulative aspects of Retinitis pigmentosa etiopathogenesis. Sci. Rep. 2018, 8, 16638. [Google Scholar] [CrossRef]
- Crouch, R.K.; Koutalos, Y.; Kono, M.; Schey, K.; Ablonczy, Z. A2E and Lipofuscin. Prog. Mol. Biol. Transl. Sci. 2015, 134, 449–463. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, J.R.; Fishkin, N.; Zhou, J.; Cai, B.; Jang, Y.P.; Krane, S.; Itagaki, Y.; Nakanishi, K. A2E, a byproduct of the visual cycle. Vis. Res. 2003, 43, 2983–2990. [Google Scholar] [CrossRef] [Green Version]
- Parmar, V.M.; Parmar, T.; Arai, E.; Perusek, L.; Maeda, A. A2E-associated cell death and inflammation in retinal pigmented epithelial cells from human induced pluripotent stem cells. Stem Cell Res. 2018, 27, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Dontsov, A.; Koromyslova, A.; Ostrovsky, M.; Sakina, N. Lipofuscins prepared by modification of photoreceptor cells via glycation or lipid peroxidation show the similar phototoxicity. World J. Exp. Med. 2016, 6, 63–71. [Google Scholar] [CrossRef]
- Alaimo, A.; Linares, G.G.; Bujjamer, J.M.; Gorojod, R.M.; Alcon, S.P.; Martinez, J.H.; Baldessari, A.; Grecco, H.E.; Kotler, M.L. Toxicity of blue led light and A2E is associated to mitochondrial dynamics impairment in ARPE-19 cells: Implications for age-related macular degeneration. Arch. Toxicol. 2019, 93, 1401–1415. [Google Scholar] [CrossRef] [PubMed]
- Alaimo, A.; Di Santo, M.C.; Dominguez Rubio, A.P.; Chaufan, G.; Garcia Linares, G.; Perez, O.E. Toxic effects of A2E in human ARPE-19 cells were prevented by resveratrol: A potential nutritional bioactive for age-related macular degeneration treatment. Arch. Toxicol. 2019, 94, 553–572. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Conesa, A.; Garcia-Alcalde, F. Qualimap 2: Advanced multi-sample quality control for high-throughput sequencing data. Bioinformatics 2016, 32, 292–294. [Google Scholar] [CrossRef]
- Li, B.; Ruotti, V.; Stewart, R.M.; Thomson, J.A.; Dewey, C.N. RNA-Seq gene expression estimation with read mapping uncertainty. Bioinformatics 2010, 26, 493–500. [Google Scholar] [CrossRef] [Green Version]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Soneson, C.; Matthes, K.L.; Nowicka, M.; Law, C.W.; Robinson, M.D. Isoform prefiltering improves performance of count-based methods for analysis of differential transcript usage. Genome Biol. 2016, 17, 12. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.B.; Wallroth, M.; Jonsson, V.; Kristiansson, E. Comparison of normalization methods for the analysis of metagenomic gene abundance data. BMC Genom. 2018, 19, 274. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.T.N.; Stephen, G.; Milne, I.; Calixto, C.; Waugh, R.; Brown, J.W.; Zhang, R. 3D RNA-seq—A powerful and flexible tool for rapid and accurate differential expression and alternative splicing analysis of RNA-seq data for biologists. bioRxiv 2019, 656686. [Google Scholar] [CrossRef]
- Ge, Y.; Sealfon, S.C.; Speed, T.P. Some Step-down Procedures Controlling the False Discovery Rate under Dependence. Stat. Sin. 2008, 18, 881–904. [Google Scholar] [PubMed]
- Jiao, X.; Sherman, B.T.; Huang da, W.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. DAVID-WS: A stateful web service to facilitate gene/protein list analysis. Bioinformatics 2012, 28, 1805–1806. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Nivison-Smith, L.; Milston, R.; Madigan, M.; Kalloniatis, M. Age-related macular degeneration: Linking clinical presentation to pathology. Optom. Vis. Sci. 2014, 91, 832–848. [Google Scholar] [CrossRef] [PubMed]
- Vancura, P.; Csicsely, E.; Leiser, A.; Iuvone, P.M.; Spessert, R. Rhythmic Regulation of Photoreceptor and RPE Genes Important for Vision and Genetically Associated With Severe Retinal Diseases. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3789–3799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuhrmann, S.; Zou, C.; Levine, E.M. Retinal pigment epithelium development, plasticity, and tissue homeostasis. Exp. Eye Res. 2014, 123, 141–150. [Google Scholar] [CrossRef] [Green Version]
- D’Angelo, R.; Donato, L.; Venza, I.; Scimone, C.; Aragona, P.; Sidoti, A. Possible protective role of the ABCA4 gene c.1268A>G missense variant in Stargardt disease and syndromic retinitis pigmentosa in a Sicilian family: Preliminary data. Int. J. Mol. Med. 2017, 39, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Donato, L.; Scimone, C.; Rinaldi, C.; Aragona, P.; Briuglia, S.; D’Ascola, A.; D’Angelo, R.; Sidoti, A. Stargardt Phenotype Associated With Two ELOVL4 Promoter Variants and ELOVL4 Downregulation: New Possible Perspective to Etiopathogenesis? Investig. Ophthalmol. Vis. Sci. 2018, 59, 843–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiktor, A.; Sarna, M.; Wnuk, D.; Sarna, T. Lipofuscin-mediated photodynamic stress induces adverse changes in nanomechanical properties of retinal pigment epithelium cells. Sci. Rep. 2018, 8, 17929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moiseyev, G.; Nikolaeva, O.; Chen, Y.; Farjo, K.; Takahashi, Y.; Ma, J.X. Inhibition of the visual cycle by A2E through direct interaction with RPE65 and implications in Stargardt disease. Proc. Natl. Acad. Sci. USA 2010, 107, 17551–17556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaarniranta, K.; Machalinska, A.; Vereb, Z.; Salminen, A.; Petrovski, G.; Kauppinen, A. Estrogen signalling in the pathogenesis of age-related macular degeneration. Curr. Eye Res. 2015, 40, 226–233. [Google Scholar] [CrossRef]
- Akanuma, S.; Soutome, T.; Hisada, E.; Tachikawa, M.; Kubo, Y.; Hosoya, K. Na+-independent nucleoside transporters regulate adenosine and hypoxanthine levels in Muller cells and the inner blood-retinal barrier. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1469–1477. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos-Rodrigues, A.; Pereira, M.R.; Brito, R.; de Oliveira, N.A.; Paes-de-Carvalho, R. Adenosine transporters and receptors: Key elements for retinal function and neuroprotection. Vitam. Horm. 2015, 98, 487–523. [Google Scholar] [CrossRef]
- Fliesler, S.J.; Keller, R.K. Isoprenoid metabolism in the vertebrate retina. Int. J. Biochem. Cell Biol. 1997, 29, 877–894. [Google Scholar] [CrossRef]
- Roysommuti, S.; Thaeomor, A.; Khimsuksri, S.; Lerdweeraphon, W.; Wyss, J.M. Perinatal taurine imbalance alters the interplay of renin-angiotensin system and estrogen on glucose-insulin regulation in adult female rats. Adv. Exp. Med. Biol. 2013, 776, 67–80. [Google Scholar] [CrossRef]
- Okabe, K.; Kobayashi, S.; Yamada, T.; Kurihara, T.; Tai-Nagara, I.; Miyamoto, T.; Mukouyama, Y.S.; Sato, T.N.; Suda, T.; Ema, M.; et al. Neurons limit angiogenesis by titrating VEGF in retina. Cell 2014, 159, 584–596. [Google Scholar] [CrossRef] [Green Version]
- Joyal, J.S.; Gantner, M.L.; Smith, L.E.H. Retinal energy demands control vascular supply of the retina in development and disease: The role of neuronal lipid and glucose metabolism. Prog. Retin. Eye Res. 2018, 64, 131–156. [Google Scholar] [CrossRef]
- Preising, M.; Ayuso, C. Rab escort protein 1 (REP1) in intracellular traffic: A functional and pathophysiological overview. Ophthalmic Genet. 2004, 25, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Kauppinen, A.; Paterno, J.J.; Blasiak, J.; Salminen, A.; Kaarniranta, K. Inflammation and its role in age-related macular degeneration. Cell Mol. Life Sci. 2016, 73, 1765–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piippo, N.; Korhonen, E.; Hytti, M.; Skottman, H.; Kinnunen, K.; Josifovska, N.; Petrovski, G.; Kaarniranta, K.; Kauppinen, A. Hsp90 inhibition as a means to inhibit activation of the NLRP3 inflammasome. Sci. Rep. 2018, 8, 6720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subrizi, A.; Toropainen, E.; Ramsay, E.; Airaksinen, A.J.; Kaarniranta, K.; Urtti, A. Oxidative stress protection by exogenous delivery of rhHsp70 chaperone to the retinal pigment epithelium (RPE), a possible therapeutic strategy against RPE degeneration. Pharm. Res. 2015, 32, 211–221. [Google Scholar] [CrossRef]
- Dhingra, A.; Alexander, D.; Reyes-Reveles, J.; Sharp, R.; Boesze-Battaglia, K. Microtubule-Associated Protein 1 Light Chain 3 (LC3) Isoforms in RPE and Retina. Adv. Exp. Med. Biol. 2018, 1074, 609–616. [Google Scholar] [CrossRef]
- Frost, L.S.; Lopes, V.S.; Bragin, A.; Reyes-Reveles, J.; Brancato, J.; Cohen, A.; Mitchell, C.H.; Williams, D.S.; Boesze-Battaglia, K. The Contribution of Melanoregulin to Microtubule-Associated Protein 1 Light Chain 3 (LC3) Associated Phagocytosis in Retinal Pigment Epithelium. Mol. Neurobiol. 2015, 52, 1135–1151. [Google Scholar] [CrossRef] [Green Version]
- Sparrow, J.R.; Zhou, J.; Cai, B. DNA is a target of the photodynamic effects elicited in A2E-laden RPE by blue-light illumination. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2245–2251. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Feng, Y.; Han, P.; Wang, F.; Luo, X.; Liang, J.; Sun, X.; Ye, J.; Lu, Y.; Sun, X. Photosensitization of A2E triggers telomere dysfunction and accelerates retinal pigment epithelium senescence. Cell Death Dis. 2018, 9, 178. [Google Scholar] [CrossRef] [Green Version]
- Cornish, E.E.; Natoli, R.C.; Hendrickson, A.; Provis, J.M. Differential distribution of fibroblast growth factor receptors (FGFRs) on foveal cones: FGFR-4 is an early marker of cone photoreceptors. Mol. Vis. 2004, 10, 1–14. [Google Scholar]
- Donato, L.; Bramanti, P.; Scimone, C.; Rinaldi, C.; Giorgianni, F.; Beranova-Giorgianni, S.; Koirala, D.; D’Angelo, R.; Sidoti, A. miRNAexpression profile of retinal pigment epithelial cells under oxidative stress conditions. FEBS Open Bio 2018, 8, 219–233. [Google Scholar] [CrossRef] [Green Version]
- Olejniczak, M.; Kotowska-Zimmer, A.; Krzyzosiak, W. Stress-induced changes in miRNA biogenesis and functioning. Cell Mol. Life Sci. 2018, 75, 177–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westlund, B.S.; Cai, B.; Zhou, J.; Sparrow, J.R. Involvement of c-Abl, p53 and the MAP kinase JNK in the cell death program initiated in A2E-laden ARPE-19 cells by exposure to blue light. Apoptosis 2009, 14, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCann, K.L.; Charette, J.M.; Vincent, N.G.; Baserga, S.J. A protein interaction map of the LSU processome. Genes Dev. 2015, 29, 862–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchesi, N.; Thongon, N.; Pascale, A.; Provenzani, A.; Koskela, A.; Korhonen, E.; Smedowski, A.; Govoni, S.; Kauppinen, A.; Kaarniranta, K.; et al. Autophagy Stimulus Promotes Early HuR Protein Activation and p62/SQSTM1 Protein Synthesis in ARPE-19 Cells by Triggering Erk1/2, p38(MAPK), and JNK Kinase Pathways. Oxid. Med. Cell Longev. 2018, 2018, 4956080. [Google Scholar] [CrossRef] [Green Version]
- McGrew, D.A.; Hedstrom, L. Towards a pathological mechanism for IMPDH1-linked retinitis pigmentosa. Adv. Exp. Med. Biol. 2012, 723, 539–545. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
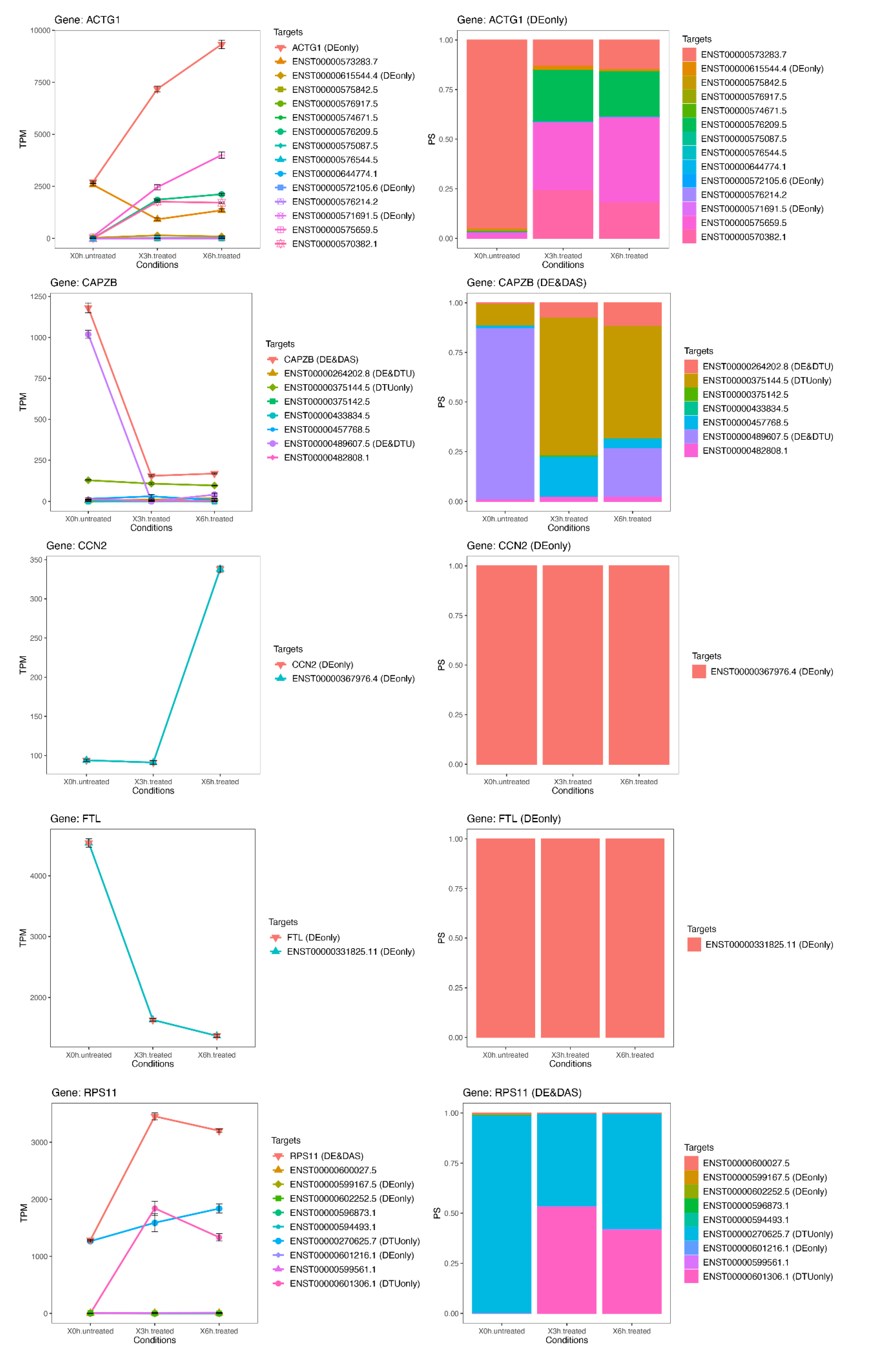{kind=link}
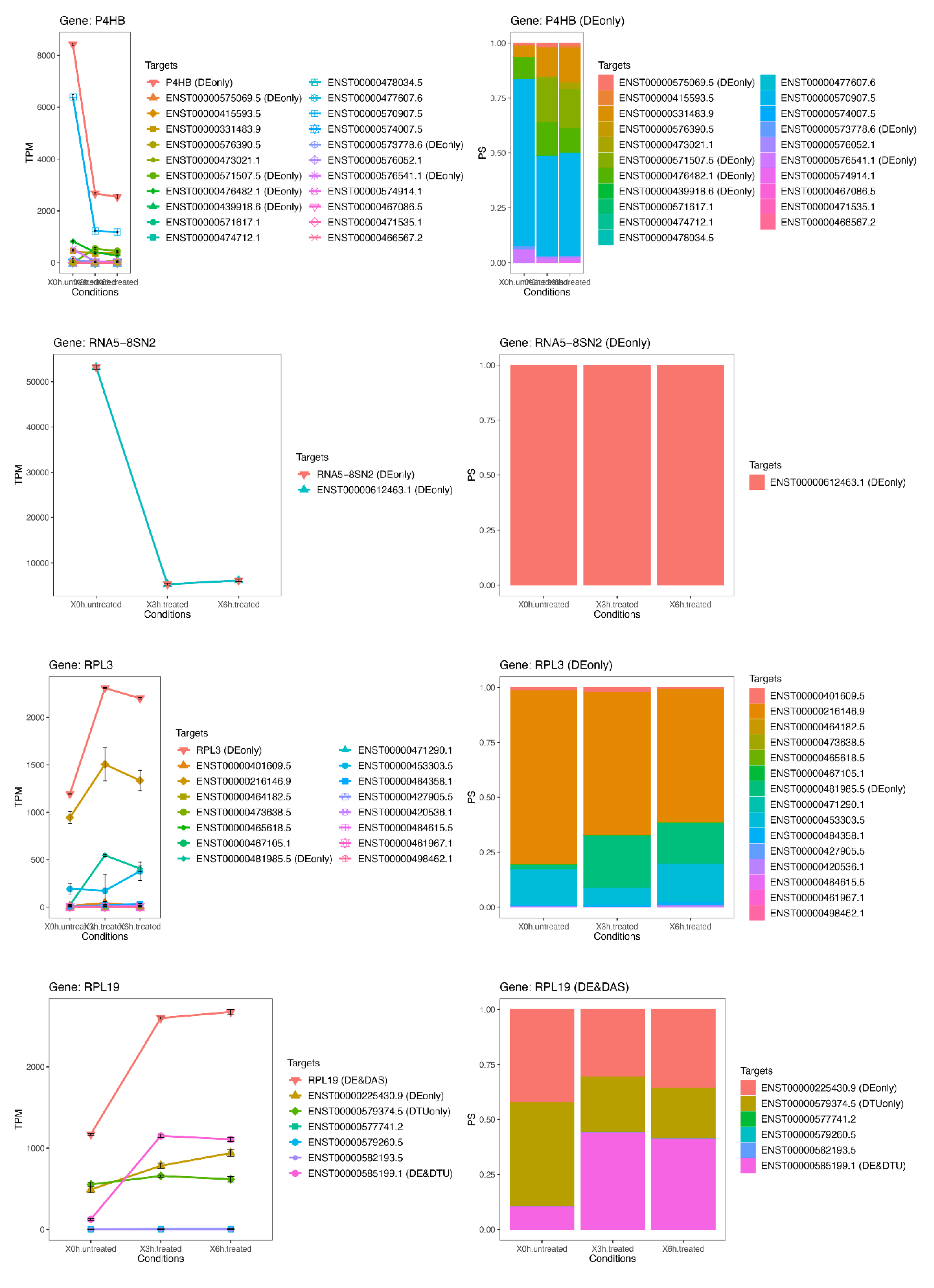{kind=link}
| Rank | Gene Symbol | GeneId | GO: Mol. Func. Score | GO: Mol. Func. p-Value | GO: Bio. Proc. Score | GO: Bio. Proc. p-Value | GO: Cell. Comp. Score | GO: Cell. Comp. p-Value | Human Pheno. Score | Human Pheno. p-Value | Pathway Score | Pathway p-Value | Pubmed Score | Pubmed p-Value | Disease Score | Disease p-Value | Average Score | Overall p-Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | PAX6 | 5080 | 0.0422 | 0.0931 | 1.0 | 0.0102 | 0.2814 | 0.0807 | 1.0 | 0.0135 | 0.0 | 0.503 | 1.0 | 0.001 | 0.999 | 7.75 × 10−10 | 0.6176 | 1 88 × 10−12 |
| 2 | ACTG1 | 71 | 0.8143 | 0.0106 | 0.996 | 0.0278 | 0.9686 | 0.0127 | 0.999 | 0.0135 | 0.0 | 0.503 | 0.330 | 0.032 | 0.730 | 0.006 | 0.7267 | 5 15 × 10−11 |
| 3 | TGFBI | 7045 | 0.6329 | 0.0288 | 0.982 | 0.0462 | 0.4683 | 0.0534 | 0.988 | 0.0331 | 0.0 | 0.503 | 0.204 | 0.073 | 0.819 | 0.003 | 0.6226 | 0.002 |
| 4 | CCN2 | 1490 | 0.4318 | 0.0491 | 0.999 | 0.0209 | 0.4362 | 0.0544 | 0.999 | 0.0253 | 0.0 | 0.503 | 0.204 | 0.073 | 0.551 | 0.011 | 0.5529 | 0.003 |
| 5 | CTSH | 1512 | 0.2796 | 0.0677 | 0.999 | 0.0185 | 0.9986 | 0.0036 | 0.976 | 0.0358 | 0.0 | 0.503 | 0.0 | 0.536 | 0.445 | 0.016 | 0.5302 | 0.003 |
| 6 | GNAI2 | 2771 | 0.9263 | 0.0029 | 0.962 | 0.0573 | 0.9500 | 0.0166 | 0.999 | 0.0195 | 0.0 | 0.503 | 0.095 | 0.073 | 0.0 | 0.512 | 0.5951 | 0.004 |
| 7 | LTBP2 | 4053 | 0.5918 | 0.0375 | 0.813 | 0.1049 | 0.0350 | 0.2254 | 0.999 | 0.0135 | 0.0 | 0.503 | 0.204 | 0.073 | 0.636 | 0.009 | 0.5168 | 0.007 |
| 8 | HNRNPA1 | 3178 | 0.0421 | 0.0931 | 0.910 | 0.0772 | 0.5481 | 0.0499 | 0.999 | 0.0214 | 0.0 | 0.503 | 0.801 | 0.006 | 0.0 | 0.512 | 0.4913 | 0.011 |
| 9 | GPI | 2821 | 0.0421 | 0.0931 | 0.965 | 0.0563 | 0.9563 | 0.0156 | 0.986 | 0.0335 | 0.0 | 0.503 | 0.490 | 0.019 | 0.0 | 0.512 | 0.5249 | 0.011 |
| 10 | CD81 | 975 | 0.6702 | 0.0213 | 0.999 | 0.0209 | 0.3057 | 0.0782 | 0.999 | 0.0232 | 0.0 | 0.503 | 0.095 | 0.073 | 0.0 | 0.512 | 0.4691 | 0.011 |
| 11 | FTL | 2512 | 0.2622 | 0.0766 | 0.587 | 0.1403 | 0.2370 | 0.0979 | 0.999 | 0.0205 | 0.0 | 0.503 | 0.095 | 0.073 | 0.331 | 0.021 | 0.4151 | 0.012 |
| 12 | ITGAV | 3685 | 0.4732 | 0.0476 | 0.999 | 0.0156 | 0.9885 | 0.0094 | −1.0 | 0.0 | 0.0 | 0.503 | 0.076 | 0.073 | 0.0 | 0.512 | 0.4563 | 0.014 |
| 13 | CAPZB | 832 | 0.5061 | 0.0426 | 0.998 | 0.0242 | 0.9934 | 0.0079 | −1.0 | 0.0 | 0.0 | 0.503 | 0.205 | 0.073 | 0.0 | 0.512 | 0.4919 | 0.015 |
| 14 | LTBP3 | 4054 | 0.0421 | 0.0931 | 0.982 | 0.0458 | 0.0350 | 0.2254 | 0.999 | 0.0178 | 0.0 | 0.503 | 0.0 | 0.536 | 0.636 | 0.009 | 0.3864 | 0.018 |
| 15 | CTSB | 1508 | 0.2996 | 0.0642 | 0.915 | 0.0753 | 0.3862 | 0.0600 | 0.183 | 0.0553 | 0.0 | 0.503 | 0.343 | 0.032 | 0.0 | 0.512 | 0.3794 | 0.026 |
| 16 | P4HB | 5034 | 0.2996 | 0.0642 | 0.851 | 0.0960 | 0.2370 | 0.0979 | 0.941 | 0.0400 | 0.0 | 0.503 | 0.351 | 0.032 | 0.0 | 0.512 | 0.4317 | 0.031 |
| 17 | MATN2 | 4147 | 0.4747 | 0.0464 | 0.987 | 0.0408 | 0.3796 | 0.0606 | −1.0 | 0.0 | 0.0 | 0.503 | 0.204 | 0.073 | 0.0 | 0.512 | 0.3834 | 0.042 |
| 18 | TMEM189-UBE2V1 | 387522 | −1.0 | 0.0 | −1.0 | 0.0 | −1.0 | 0.0 | −1.0 | 0.0 | 0.0 | 0.503 | 0.059 | 0.073 | 0.579 | 0.011 | 0.2746 | 0.050 |
| 19 | ACADVL | 37 | 0.6272 | 0.0290 | 0.903 | 0.0792 | 0.2422 | 0.0857 | 0.980 | 0.0348 | 0.0 | 0.503 | 0.0 | 0.536 | 0.0 | 0.512 | 0.4014 | 0.053 |
| 20 | CD151 | 977 | 0.3015 | 0.0639 | 0.776 | 0.1113 | 0.4489 | 0.0542 | 0.994 | 0.0313 | 0.0 | 0.503 | 0.0 | 0.536 | 0.0 | 0.512 | 0.3755 | 0.065 |
| 21 | CAPNS1 | 826 | 0.0421 | 0.0931 | 0.819 | 0.1039 | 0.1428 | 0.1432 | −1.0 | 0.0 | 0.0 | 0.503 | 0.076 | 0.073 | 0.0 | 0.512 | 0.2277 | 0.103 |
| 22 | CD63 | 967 | 0.0 | 0.5652 | 0.986 | 0.0416 | 0.3390 | 0.0613 | −1.0 | 0.0 | 0.0 | 0.503 | 0.095 | 0.073 | 0.0 | 0.512 | 0.2735 | 0.104 |
| 23 | RNA5-8SN2 | 109864281 | 0.3250 | 0.0559 | −1.0 | 0.0 | 0.2466 | 0.0846 | −1.0 | 0.0 | −1.0 | 0.0 | 0.0 | 0.536 | −1.0 | 0.0 | 0.1905 | 0.112 |
| 24 | RPL3 | 6122 | 0.3250 | 0.0559 | 0.952 | 0.0609 | 0.2814 | 0.0807 | −1.0 | 0.0 | 0.0 | 0.503 | 0.0 | 0.536 | 0.0 | 0.512 | 0.2637 | 0.117 |
| 25 | RPS11 | 6205 | 0.3250 | 0.0559 | 0.877 | 0.0861 | 0.2814 | 0.0807 | −1.0 | 0.0 | 0.0 | 0.503 | 0.0 | 0.536 | 0.0 | 0.512 | 0.2571 | 0.132 |
| 26 | RPL19 | 6143 | 0.3250 | 0.0559 | 0.877 | 0.0861 | 0.2814 | 0.0807 | −1.0 | 0.0 | 0.0 | 0.503 | 0.0 | 0.536 | 0.0 | 0.512 | 0.2571 | 0.132 |
| 27 | SLC16A3 | 9123 | 0.0 | 0.5652 | 0.212 | 0.2039 | 0.3057 | 0.0782 | −1.0 | 0.0 | 0.0 | 0.503 | 0.065 | 0.073 | 0.0 | 0.512 | 0.1703 | 0.192 |
| 28 | UTP14C | 9724 | 0.0 | 0.5652 | 0.220 | 0.2037 | 0.2814 | 0.0807 | −1.0 | 0.0 | 0.0 | 0.503 | 0.0 | 0.536 | 0.0 | 0.512 | 0.1251 | 0.353 |
| 29 | TMSB4XP6 | 7120 | −1.0 | 0.0 | −1.0 | 0.0 | −1.0 | 0.0 | −1.0 | 0.0 | −1.0 | 0.0 | 0.0 | 0.536 | −1.0 | 0.0 | 0.0 | 1.0 |
| De Gene-Involved Pathways | Expression Changes | Das Gene-Involved Pathways |
|---|---|---|
| RNA methyltransferase | 3 h and 6 h = DOWN-REGULATED | Endosomal sorting complex required for transport (ESCRT) and RAB geranylgeranylation |
| TRAF6 mediated NF-kB activation and activation of IKK by MEKK1 | Phagopore assembly site membrane; C-terminal protein lipidation; protein localization to microtubule cytoskeleton; regulation of TNFR1 signaling; TNF signaling | |
| Condensed chromosome outer kinetochore and kinesin complex | Methylation; activation of chaperone genes; nucleotide-sugar biosynthetic process | |
| Transport of nucleoside and free purine and pyrimidine; histone pre-mRNA DCP binding; formation of AT-AC complex; respiratory electron transport | 3 h and 6 h = UP-REGULATED | Negative regulation of FGFR1 signaling |
| Regulation of telomerase RNA localization to Cajal body; maturation of LSU-rRNA; miRNAs involved in DNA damage response | ||
| Mithocondrial intermembrane space and TP53 regulates transcription of cell death genes | ||
| Polysomal ribosome | ||
| Mevalonate pathway and cholesterol biosynthesis | 3 h = UP-REGULATED 6 h = DOWN-REGULATED | Cholesterol biosynthesis |
| CBL binds and ubiquinates Sprouty and MAP kinase phosphatase activity | 3 h = DOWN-REGULATED 6 h = UP-REGULATED | / |
| PTK2/SRC-1 phosphorylates BCAR1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donato, L.; D’Angelo, R.; Alibrandi, S.; Rinaldi, C.; Sidoti, A.; Scimone, C. Effects of A2E-Induced Oxidative Stress on Retinal Epithelial Cells: New Insights on Differential Gene Response and Retinal Dystrophies. Antioxidants 2020, 9, 307. https://doi.org/10.3390/antiox9040307
Donato L, D’Angelo R, Alibrandi S, Rinaldi C, Sidoti A, Scimone C. Effects of A2E-Induced Oxidative Stress on Retinal Epithelial Cells: New Insights on Differential Gene Response and Retinal Dystrophies. Antioxidants. 2020; 9(4):307. https://doi.org/10.3390/antiox9040307
Chicago/Turabian StyleDonato, Luigi, Rosalia D’Angelo, Simona Alibrandi, Carmela Rinaldi, Antonina Sidoti, and Concetta Scimone. 2020. "Effects of A2E-Induced Oxidative Stress on Retinal Epithelial Cells: New Insights on Differential Gene Response and Retinal Dystrophies" Antioxidants 9, no. 4: 307. https://doi.org/10.3390/antiox9040307
APA StyleDonato, L., D’Angelo, R., Alibrandi, S., Rinaldi, C., Sidoti, A., & Scimone, C. (2020). Effects of A2E-Induced Oxidative Stress on Retinal Epithelial Cells: New Insights on Differential Gene Response and Retinal Dystrophies. Antioxidants, 9(4), 307. https://doi.org/10.3390/antiox9040307