Antioxidative Effects of Thymus quinquecostatus CELAK through Mitochondrial Biogenesis Improvement in RAW 264.7 Macrophages



Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Extracts from the TQC
2.2. Cell Culture and Treatment
2.3. Cell Proliferation Assay
2.4. Nitric Oxide and H2O2 Assay
2.5. Mitochondrial ATP Assay
2.6. Immunocytochemistry (ICC)
2.7. Mitochondrial Staining
2.8. RNA Isolation and Real-Time PCR
2.9. Genomic DNA Preparation and Dot Blot
2.10. Western Blot
2.11. Flow Cytometry
2.12. Statistics
3. Results
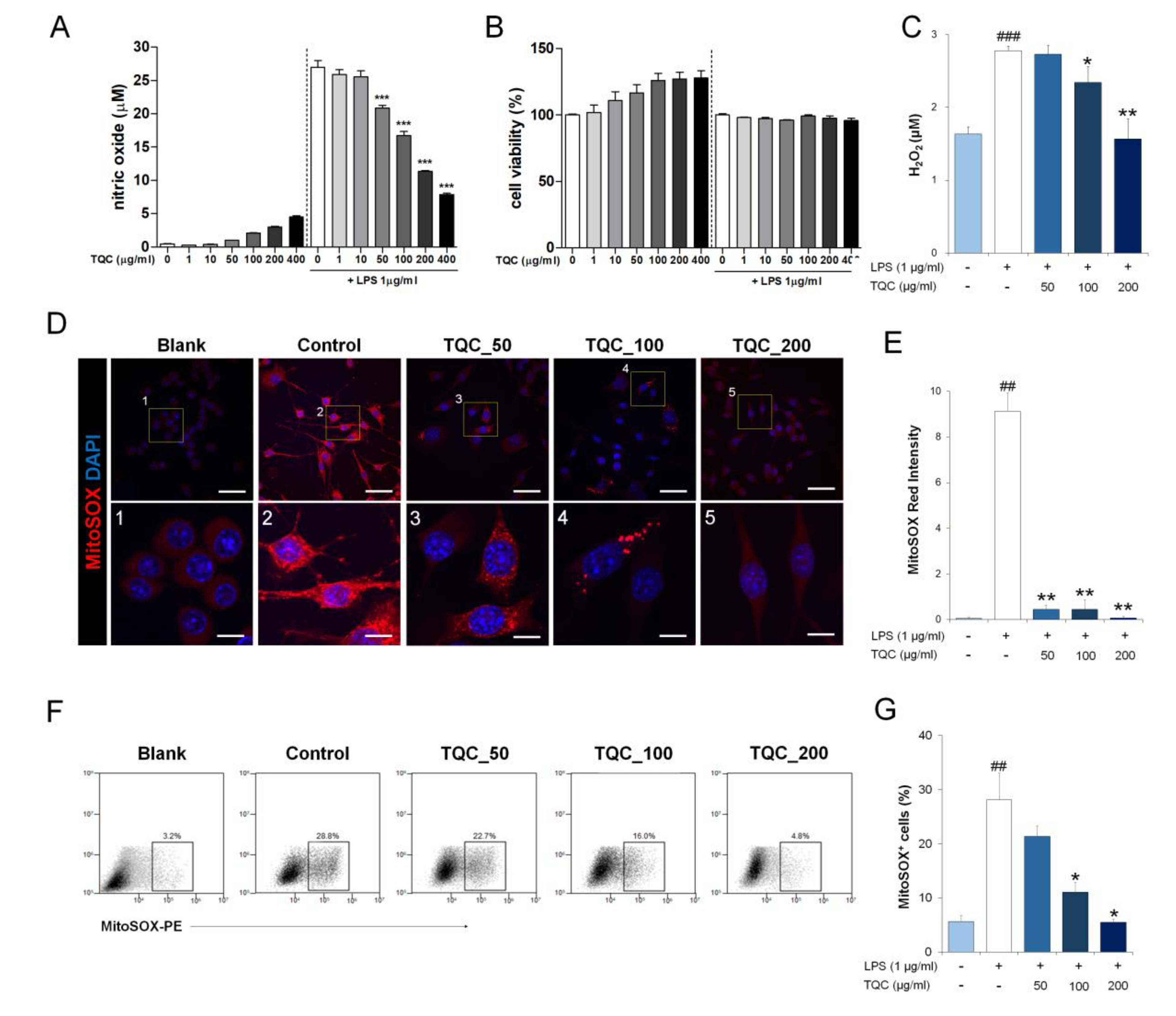
3.1. TQC Reduces Mitochondrial ROS and Nitric Oxide Production
3.2. TQC Inhibits the Oxidation of DNA in LPS-Activated RAW 264.7 Macrophages
3.3. TQC Induces Morphological Changes and Alterations in Mitochondria
3.4. TQC Enhances Mitochondrial Biogenesis in LPS-Activated RAW 264.7 Macrophages
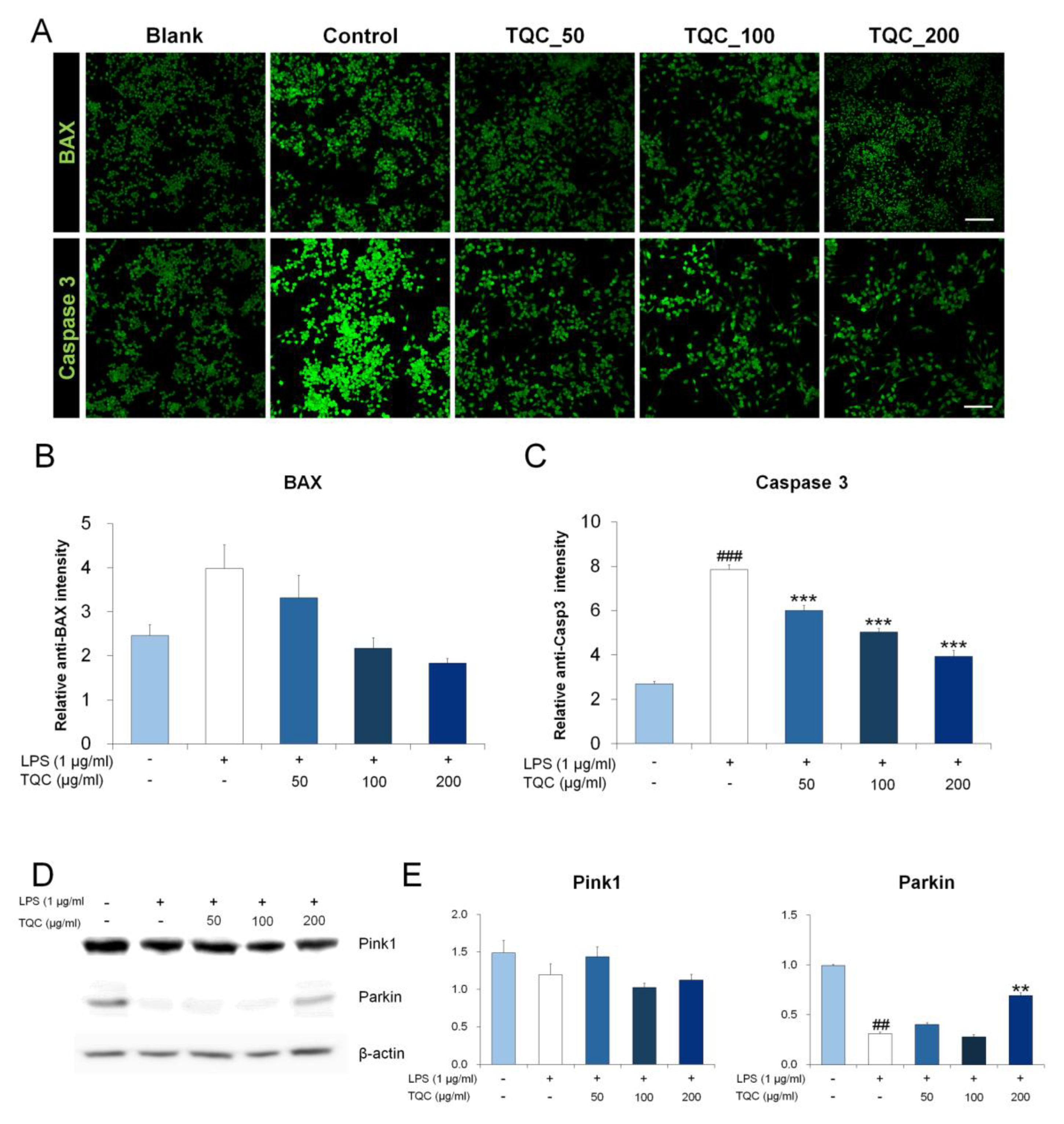
3.5. TQC Reduces Oxidative Stress and Mitochondrial Apoptosis in LPS-Activated RAW 264.7 Macrophages
3.6. TQC Prevents Apoptosis by Inhibiting BAX/Caspase-3 Expression and Induces Parkin Expression in LPS-Activated Macrophages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rao, A.V.; Balachandran, B. Role of oxidative stress and antioxidants in neurodegenerative diseases. Nutr. Neurosci. 2002, 5, 291–309. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Halliwell, B. Antioxidants in human health and disease. Annu. Rev. Nutr. 1996, 16, 33–50. [Google Scholar] [CrossRef]
- Raquel, R.; Bartz, C.A.P. Clinical review Oxygen as a signaling molecule. Crit. Care 2010, 14, 234. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, J.; Duchen, M.R. Mitochondrial oxidative stress and cell death in astrocytes--requirement for stored Ca2+ and sustained opening of the permeability transition pore. J. Cell Sci. 2002, 115, 1175–1188. [Google Scholar]
- Ott, M.; Gogvadze, V.; Orrenius, S.; Zhivotovsky, B. Mitochondria, oxidative stress and cell death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef]
- Ding, W.; Liu, T.; Bi, X.; Zhang, Z. Mitochondria-Targeted Antioxidant Mito-Tempo Protects Against Aldosterone-Induced Renal Injury In Vivo. Cell. Physiol. Biochem. 2017, 44, 741–750. [Google Scholar] [CrossRef]
- Broome, S.C.; Woodhead, J.S.T.; Merry, T.L. Mitochondria-Targeted Antioxidants and Skeletal Muscle Function. Antioxidants 2018, 7, 107. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Jiang, Z.; Xia, B.; Zhang, L.; Zhang, C.; Leng, J. Mitochondria-targeted antioxidant therapy for an animal model of PCOS-IR. Int. J. Mol. Med. 2019, 43, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Kezic, A.; Spasojevic, I.; Lezaic, V.; Bajcetic, M. Mitochondria-Targeted Antioxidants: Future Perspectives in Kidney Ischemia Reperfusion Injury. Oxid. Med. Cell. Longev. 2016, 2016, 2950503. [Google Scholar] [CrossRef] [Green Version]
- Ranjbar, A.; Kheiripour, N.; Ghasemi, H.; Seif Rabiei, M.A.; Dadras, F.; Khoshjou, F. Antioxidative Effects of Tempol on Mitochondrial Dysfunction in Diabetic Nephropathy. Iran. J. Kidney Dis. 2018, 12, 84–90. [Google Scholar]
- Im, A.R.; Kim, Y.H.; Uddin, M.R.; Lee, H.W.; Chae, S.W.; Kim, Y.H.; Jung, W.S.; Kang, B.J.; Mun, C.S.; Lee, M.Y. Scutellaria baicalensis Extracts and Flavonoids Protect Rat L6 Cells from Antimycin A-Induced Mitochondrial Dysfunction. Evid. Based Complement. Altern. Med. 2012, 2012, 517965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Hwang, J.W.; Kang, S.H.; Kim, E.H.; Jeon, Y.J.; Jeong, J.H.; Kim, H.R.; Moon, S.H.; Jeon, B.T.; Park, P.J. Thymol from Thymus quinquecostatus Celak. protects against tert-Butyl hydroperoxide-Induced oxidative stress in Chang cells. J. Nat. Med. 2014, 68, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Hyun, T.K.; Kim, H.-C.; Kim, J.-S. Antioxidant and antidiabetic activity of Thymus quinquecostatus Celak. Ind. Crop. Prod. 2014, 52, 611–616. [Google Scholar] [CrossRef]
- Kim, Y.S.; Lee, S.J.; Hwang, J.W.; Kim, E.K.; Kim, S.E.; Kim, E.H.; Moon, S.H.; Jeon, B.T.; Park, P.J. In vitro protective effects of Thymus quinquecostatus Celak extracts on t-BHP-induced cell damage through antioxidant activity. Food Chem. Toxicol. 2012, 50, 4191–4198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, R.; Li, L.; Zhu, L.; Gao, S.; Lu, Q.; Gu, Y.; Zhang, Y.; Yang, H.; Hou, T.; et al. Macrophage migration inhibitory factor (MIF) inhibitor, Z-590 suppresses cartilage destruction in adjuvant-induced arthritis via inhibition of macrophage inflammatory activation. Immunopharmacol. Immunotoxicol. 2018, 40, 149–157. [Google Scholar] [CrossRef]
- Wang, H.M.; Fu, L.; Cheng, C.C.; Gao, R.; Lin, M.Y.; Su, H.L.; Belinda, N.E.; Nguyen, T.H.; Lin, W.H.; Lee, P.C.; et al. Inhibition of LPS-Induced Oxidative Damages and Potential Anti-Inflammatory Effects of Phyllanthus emblica Extract via Down-Regulating NF-kappaB, COX-2, and iNOS in RAW 264.7 Cells. Antioxidants 2019, 8, 270. [Google Scholar] [CrossRef] [Green Version]
- Vuong, L.D.; Nguyen, Q.N.; Truong, V.L. Anti-Inflammatory and anti-Oxidant effects of combination between sulforaphane and acetaminophen in LPS-stimulated RAW 264.7 macrophage cells. Immunopharmacol. Immunotoxicol. 2019, 41, 413–419. [Google Scholar] [CrossRef]
- Kauffman, M.E.; Kauffman, M.K.; Traore, K.; Zhu, H.; Trush, M.A.; Jia, Z.; Li, Y.R. MitoSOX-Based Flow Cytometry for Detecting Mitochondrial ROS. React. Oxyg. Species (Apex) 2016, 2, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Hull, T.D.; Boddu, R.; Guo, L.; Tisher, C.C.; Traylor, A.M.; Patel, B.; Joseph, R.; Prabhu, S.D.; Suliman, H.B.; Piantadosi, C.A.; et al. Heme oxygenase-1 regulates mitochondrial quality control in the heart. JCI Insight 2016, 1, e85817. [Google Scholar] [CrossRef] [Green Version]
- Converso, D.P.; Taille, C.; Carreras, M.C.; Jaitovich, A.; Poderoso, J.J.; Boczkowski, J. HO-1 is located in liver mitochondria and modulates mitochondrial heme content and metabolism. FASEB J. 2006, 20, 1236–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopper, C.P.; Meinel, L.; Steiger, C.; Otterbein, L.E. Where is the Clinical Breakthrough of Heme Oxygenase-1/Carbon Monoxide Therapeutics? Curr. Pharm. Des. 2018, 24, 2264–2282. [Google Scholar] [CrossRef] [PubMed]
- Ott, M.; Zhivotovsky, B.; Orrenius, S. Role of cardiolipin in cytochrome c release from mitochondria. Cell Death Differ. 2007, 14, 1243–1247. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, K.M.; Jeong, J.U.; Shin, J.H.; Shin, J.M.; Bang, K.T. Nrf2-Heme oxygenase-1 modulates autophagy and inhibits apoptosis triggered by elevated glucose levels in renal tubule cells. Kidney Res. Clin. Pract. 2019, 38, 318–325. [Google Scholar] [CrossRef] [Green Version]
- Piantadosi, C.A.; Withers, C.M.; Bartz, R.R.; MacGarvey, N.C.; Fu, P.; Sweeney, T.E.; Welty-Wolf, K.E.; Suliman, H.B. Heme oxygenase-1 couples activation of mitochondrial biogenesis to anti-Inflammatory cytokine expression. J. Biol. Chem. 2011, 286, 16374–16385. [Google Scholar] [CrossRef] [Green Version]
- Wagner, M.; Cadetg, P.; Ruf, R.; Mazzucchelli, L.; Ferrari, P.; Redaelli, C.A. Heme oxygenase-1 attenuates ischemia/reperfusion-induced apoptosis and improves survival in rat renal allografts. Kidney Int. 2003, 63, 1564–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finucane, D.M.; Bossy-Wetzel, E.; Waterhouse, N.J.; Cotter, T.G.; Green, D.R. Bax-induced caspase activation and apoptosis via cytochrome c release from mitochondria is inhibitable by Bcl-xL. J. Biol. Chem. 1999, 274, 2225–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vince, J.E.; De Nardo, D.; Gao, W.; Vince, A.J.; Hall, C.; McArthur, K.; Simpson, D.; Vijayaraj, S.; Lindqvist, L.M.; Bouillet, P.; et al. The Mitochondrial Apoptotic Effectors BAX/BAK Activate Caspase-3 and -7 to Trigger NLRP3 Inflammasome and Caspase-8 Driven IL-1beta Activation. Cell Rep. 2018, 25, 2339–2353 e2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.M.; Youle, R.J. PINK1-and Parkin-mediated mitophagy at a glance. J. Cell Sci. 2012, 125, 795–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eiyama, A.; Okamoto, K. PINK1/Parkin-mediated mitophagy in mammalian cells. Curr. Opin. Cell Biol. 2015, 33, 95–101. [Google Scholar] [CrossRef]
- Truban, D.; Hou, X.; Caulfield, T.R.; Fiesel, F.C.; Springer, W. PINK1, Parkin, and Mitochondrial Quality Control: What can we Learn about Parkinson’s Disease Pathobiology? J. Parkinsons Dis. 2017, 7, 13–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickrell, A.M.; Youle, R.J. The roles of PINK1, parkin, and mitochondrial fidelity in Parkinson’s disease. Neuron 2015, 85, 257–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narendra, D.; Tanaka, A.; Suen, D.F.; Youle, R.J. Parkin is recruited selectively to impaired mitochondria and promotes their autophagy. J. Cell Biol. 2008, 183, 795–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albers, D.S.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in aging and neurodegenerative disease. J. Neural Transm. Suppl. 2000, 59, 133–154. [Google Scholar] [CrossRef]
- Quinonez-Flores, C.M.; Gonzalez-Chavez, S.A.; Del Rio Najera, D.; Pacheco-Tena, C. Oxidative Stress Relevance in the Pathogenesis of the Rheumatoid Arthritis: A Systematic Review. Biomed. Res. Int. 2016, 2016, 6097417. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Youle, R.J. The role of mitochondria in apoptosis*. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Uren, R.T.; Dewson, G.; Bonzon, C.; Lithgow, T.; Newmeyer, D.D.; Kluck, R.M. Mitochondrial release of pro-apoptotic proteins: Electrostatic interactions can hold cytochrome c but not Smac/DIABLO to mitochondrial membranes. J. Biol. Chem. 2005, 280, 2266–2274. [Google Scholar] [CrossRef] [Green Version]
- Garrido, C.; Galluzzi, L.; Brunet, M.; Puig, P.E.; Didelot, C.; Kroemer, G. Mechanisms of cytochrome c release from mitochondria. Cell Death Differ. 2006, 13, 1423–1433. [Google Scholar] [CrossRef] [Green Version]
- Ott, M.; Robertson, J.D.; Gogvadze, V.; Zhivotovsky, B.; Orrenius, S. Cytochrome c release from mitochondria proceeds by a two-Step process. Proc. Natl. Acad. Sci. USA 2002, 99, 1259–1263. [Google Scholar] [CrossRef] [Green Version]
- Jia, P.; Gao, T.; Xin, H. Changes in structure and histochemistry of glandular trichomes of Thymus quinquecostatus Celak. ScientificWorldJournal 2012, 2012, 187261. [Google Scholar] [CrossRef] [Green Version]
- Deb, D.D.; Parimala, G.; Saravana Devi, S.; Chakraborty, T. Effect of thymol on peripheral blood mononuclear cell PBMC and acute promyelotic cancer cell line HL-60. Chem. Biol. Interact. 2011, 193, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Reddy, P.H.; Iijima, M.; Sesaki, H. Mitochondrial division and fusion in metabolism. Curr. Opin. Cell Biol. 2015, 33, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, A.R.; Burke, N.; Dongworth, R.K.; Hausenloy, D.J. Mitochondrial fusion and fission proteins: Novel therapeutic targets for combating cardiovascular disease. Br. J. Pharmacol. 2014, 171, 1890–1906. [Google Scholar] [CrossRef]
- Zhan, M.; Brooks, C.; Liu, F.; Sun, L.; Dong, Z. Mitochondrial dynamics: Regulatory mechanisms and emerging role in renal pathophysiology. Kidney Int. 2013, 83, 568–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacGarvey, N.C.; Suliman, H.B.; Bartz, R.R.; Fu, P.; Withers, C.M.; Welty-Wolf, K.E.; Piantadosi, C.A. Activation of mitochondrial biogenesis by heme oxygenase-1-mediated NF-E2-related factor-2 induction rescues mice from lethal Staphylococcus aureus sepsis. Am. J. Respir. Crit. Care Med. 2012, 185, 851–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piantadosi, C.A.; Carraway, M.S.; Babiker, A.; Suliman, H.B. Heme oxygenase-1 regulates cardiac mitochondrial biogenesis via Nrf2-mediated transcriptional control of nuclear respiratory factor-1. Circ. Res. 2008, 103, 1232–1240. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | 5′-3′ | Primer Sequence |
|---|---|---|
| Drp1 | Forward | AGGTTGCCCGTGACAAATGA |
| Reverse | CACAGGCATCAGCAAAGTCG | |
| Fis1 | Forward | GCCTGGTTCGAAGCAAATAC |
| Reverse | CACGGCCAGGTAGAAGACAT | |
| Mfn1 | Forward | TTGCCACAAGCTGTGTTCGG |
| Reverse | TCTAGGGACCTGAAAGATGGGC | |
| Mfn2 | Forward | GGGGCCTACATCCAAGAGAG |
| Reverse | GCAGAACTTTGTCCCAGAGC | |
| HO-1 | Forward | CCCACCAAGTTCAAACAGCTC |
| Reverse | AGGAAGGCGGTCTTAGCCTC | |
| PGC1α | Forward | CACCTGAGTTTTGATGTTGATGG |
| Reverse | TCCTGAAAGTAGCCCTGTCTTGT | |
| Nrf1 | Forward | GCAGGTGGTTTATGGGATGTTT |
| Reverse | TTTGGGTTCAGGAGTTGTTGTG | |
| TFAM | Forward | CACTGAGCATCTCCCTCACA |
| Reverse | GAGGGTGCAGCGAACTTTAT | |
| iNOS | Forward | CAGATCGAGCCCTGGAAGAC |
| Reverse | CTGGTCCATGCAGACAACCT | |
| Nrf2 | Forward | GATCCGCCAGCTACTCCCAGGTTG |
| Reverse | CAGGGCAAGCGACTCATGGTCATC | |
| β-actin | Forward | GTGATGGTGGGAATGGGTCAG |
| Reverse | TCACGGTTGGCCTTAGGGTTC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, J.Y.; Kim, H.; Jeon, W.-J.; Baek, S.; Ha, I.-H. Antioxidative Effects of Thymus quinquecostatus CELAK through Mitochondrial Biogenesis Improvement in RAW 264.7 Macrophages. Antioxidants 2020, 9, 548. https://doi.org/10.3390/antiox9060548
Hong JY, Kim H, Jeon W-J, Baek S, Ha I-H. Antioxidative Effects of Thymus quinquecostatus CELAK through Mitochondrial Biogenesis Improvement in RAW 264.7 Macrophages. Antioxidants. 2020; 9(6):548. https://doi.org/10.3390/antiox9060548
Chicago/Turabian StyleHong, Jin Young, Hyunseong Kim, Wan-Jin Jeon, Seungho Baek, and In-Hyuk Ha. 2020. "Antioxidative Effects of Thymus quinquecostatus CELAK through Mitochondrial Biogenesis Improvement in RAW 264.7 Macrophages" Antioxidants 9, no. 6: 548. https://doi.org/10.3390/antiox9060548
APA StyleHong, J. Y., Kim, H., Jeon, W. -J., Baek, S., & Ha, I. -H. (2020). Antioxidative Effects of Thymus quinquecostatus CELAK through Mitochondrial Biogenesis Improvement in RAW 264.7 Macrophages. Antioxidants, 9(6), 548. https://doi.org/10.3390/antiox9060548