Oral Administration of Sodium Nitrate to Metabolic Syndrome Patients Attenuates Mild Inflammatory and Oxidative Responses to Acute Exercise


 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Basal Measurements
2.3. Experimental Procedure
2.4. Plasma, PBMCs, and Neutrophils Isolation
2.5. Cytokine and Oxylipin Production by PBMCs and Neutrophils
2.6. Cytokine and Adhesion Molecules Determination
2.7. Oral Nitrate-Reducing Capability
2.8. Nitrite and Nitrate Concentrations
2.9. Oxylipins and Free Fatty Acid Determination
2.10. Statistical Analysis
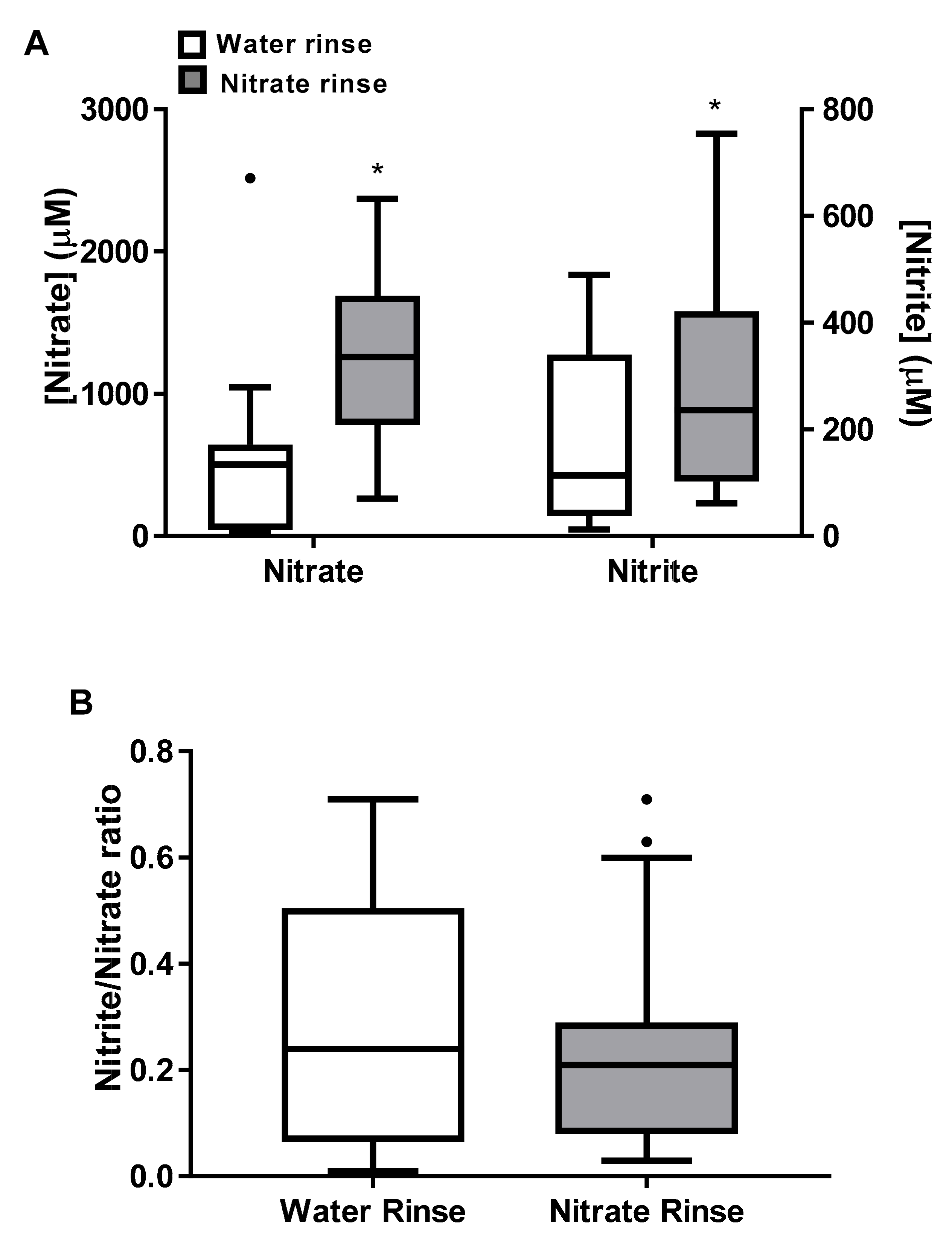
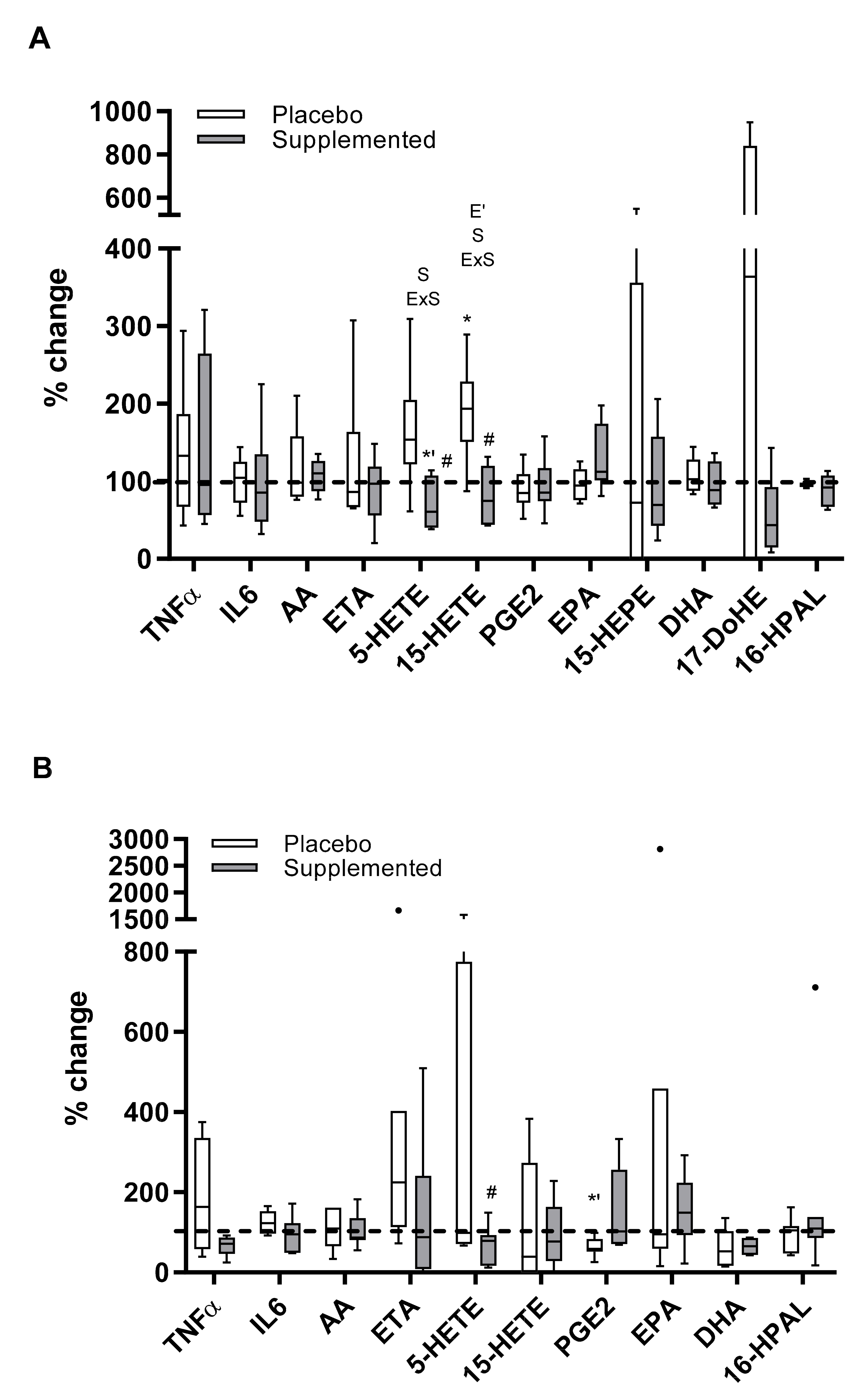
3. Results
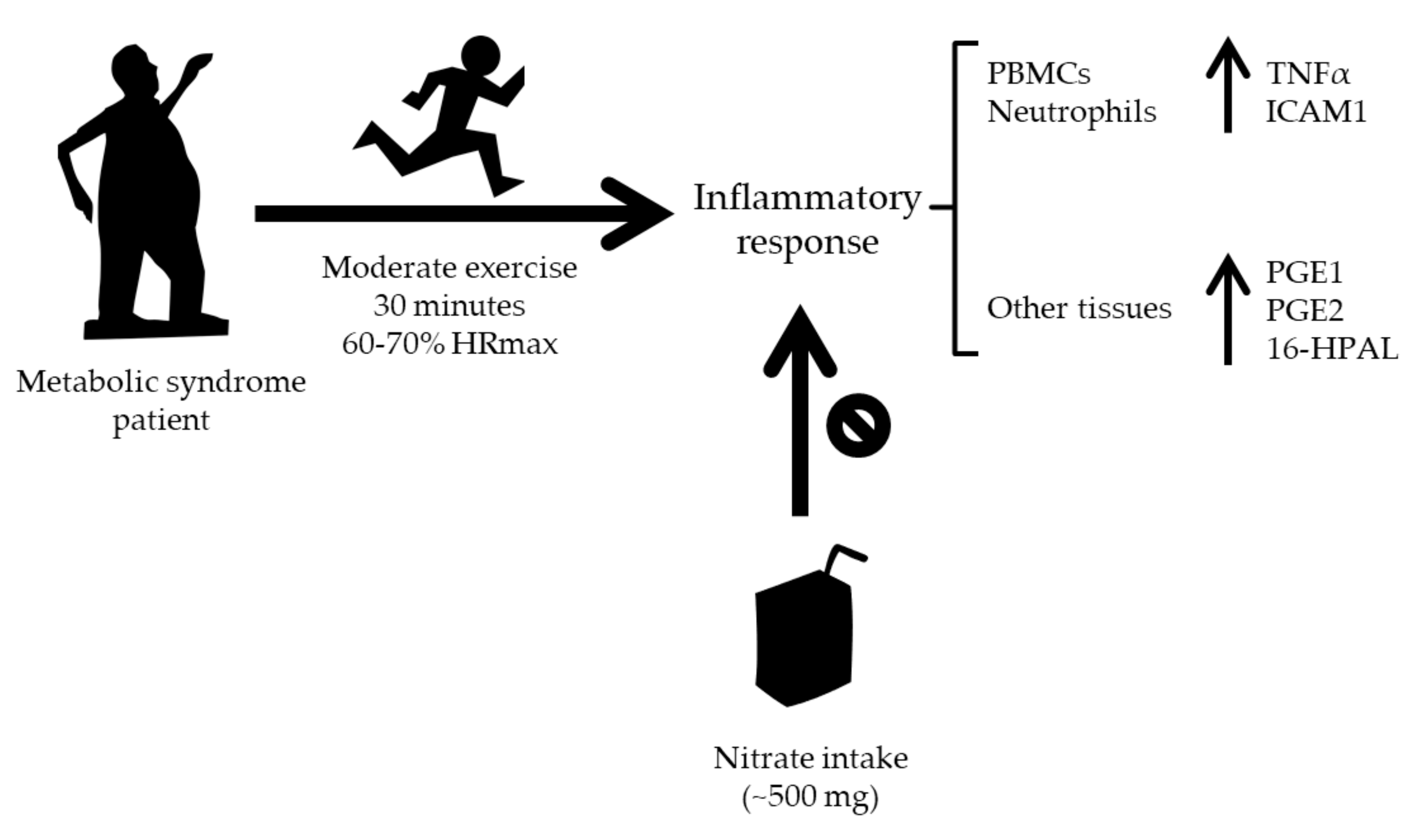
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gremeaux, V.; Gayda, M.; Lepers, R.; Sosner, P.; Juneau, M.; Nigam, A. Exercise and longevity. Maturitas 2012, 73, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.P.; Wai, J.P.; Tsai, M.K.; Yang, Y.C.; Cheng, T.Y.; Lee, M.C.; Chan, H.T.; Tsao, C.K.; Tsai, S.P.; Wu, X. Minimum amount of physical activity for reduced mortality and extended life expectancy: A prospective cohort study. Lancet 2011, 378, 1244–1253. [Google Scholar] [CrossRef]
- Vina, J.; Rodriguez-Manas, L.; Salvador-Pascual, A.; Tarazona-Santabalbina, F.J.; Gomez-Cabrera, M.C. Exercise: The lifelong supplement for healthy ageing and slowing down the onset of frailty. J. Physiol. 2016, 594, 1989–1999. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, M.D.; Capo, X.; Martorell, M.; Busquets-Cortes, C.; Bouzas, C.; Carreres, S.; Mateos, D.; Sureda, A.; Tur, J.A.; Pons, A. Regular Practice of Moderate Physical Activity by Older Adults Ameliorates Their Anti-Inflammatory Status. Nutrients 2018, 10, 1780. [Google Scholar] [CrossRef] [Green Version]
- Venditti, P.; Zhang, Y.; Radak, Z.; Magalhaes, J. Exercise in the Prevention and Management of Oxidative Stress-Linked Diseases. Oxidative Med. Cell. Longev. 2018, 2018, 7643721. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.E.; Rejeski, W.J.; Blair, S.N.; Duncan, P.W.; Judge, J.O.; King, A.C.; Macera, C.A.; Castaneda-Sceppa, C. Physical activity and public health in older adults: Recommendation from the American College of Sports Medicine and the American Heart Association. Circulation 2007, 116, 1094–1105. [Google Scholar] [CrossRef] [Green Version]
- Singh, R. The importance of exercise as a therapeutic agent. Malays. J. Med Sci. MJMS 2002, 9, 7–16. [Google Scholar]
- Pedersen, B.K.; Febbraio, M.A. Muscles, exercise and obesity: Skeletal muscle as a secretory organ. Nat. Rev. Endocrinol. 2012, 8, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Catoire, M.; Mensink, M.; Kalkhoven, E.; Schrauwen, P.; Kersten, S. Identification of human exercise-induced myokines using secretome analysis. Physiol. Genom. 2014, 46, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Pourteymour, S.; Eckardt, K.; Holen, T.; Langleite, T.; Lee, S.; Jensen, J.; Birkeland, K.I.; Drevon, C.A.; Hjorth, M. Global mRNA sequencing of human skeletal muscle: Search for novel exercise-regulated myokines. Mol. Metab. 2017, 6, 352–365. [Google Scholar] [CrossRef]
- Barquissau, V.; Ghandour, R.A.; Ailhaud, G.; Klingenspor, M.; Langin, D.; Amri, E.Z.; Pisani, D.F. Control of adipogenesis by oxylipins, GPCRs and PPARs. Biochimie 2017, 136, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Shichiri, M.; Adkins, Y.; Ishida, N.; Umeno, A.; Shigeri, Y.; Yoshida, Y.; Fedor, D.M.; Mackey, B.E.; Kelley, D.S. DHA concentration of red blood cells is inversely associated with markers of lipid peroxidation in men taking DHA supplement. J. Clin. Biochem. Nutr. 2014, 55, 196–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vane, J.R. Adventures and excursions in bioassay: The stepping stones to prostacyclin. Nobel Lecture, 8 December 1982. Biosci. Rep. 1983, 3, 683–711. [Google Scholar] [CrossRef]
- Trappe, T.A.; Fluckey, J.D.; White, F.; Lambert, C.P.; Evans, W.J. Skeletal muscle PGF(2)(alpha) and PGE(2) in response to eccentric resistance exercise: Influence of ibuprofen acetaminophen. J. Clin. Endocrinol. Metab. 2001, 86, 5067–5070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trappe, T.A.; Standley, R.A.; Jemiolo, B.; Carroll, C.C.; Trappe, S.W. Prostaglandin and myokine involvement in the cyclooxygenase-inhibiting drug enhancement of skeletal muscle adaptations to resistance exercise in older adults. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R198–R205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’El-Rei, J.; Cunha, A.R.; Trindade, M.; Neves, M.F. Beneficial Effects of Dietary Nitrate on Endothelial Function and Blood Pressure Levels. Int. J. Hypertens. 2016, 2016, 6791519. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, J.O.; Weitzberg, E.; Gladwin, M.T. The nitrate-nitrite-nitric oxide pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 2008, 7, 156–167. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Carlstrom, M.; Larsen, F.J.; Weitzberg, E. Roles of dietary inorganic nitrate in cardiovascular health and disease. Cardiovasc. Res. 2011, 89, 525–532. [Google Scholar] [CrossRef]
- Sureda, A.; Batle, J.M.; Tauler, P.; Ferrer, M.D.; Tur, J.A.; Pons, A. Vitamin C supplementation influences the antioxidant response and nitric oxide handling of erythrocytes and lymphocytes to diving apnea. Eur. J. Clin. Nutr. 2006, 60, 838–846. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Govoni, M. Inorganic nitrate is a possible source for systemic generation of nitric oxide. Free Radic. Biol. Med. 2004, 37, 395–400. [Google Scholar] [CrossRef]
- Thomas, B.; Smallwood, S.; Cutler, C.; Bescos, R. The oral nitrate-reducing capacity correlates with peak power output and peak oxygen uptake in healthy humans. Nitric. Oxide 2019, 87, 43–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Velzen, A.G.; Sips, A.J.; Schothorst, R.C.; Lambers, A.C.; Meulenbelt, J. The oral bioavailability of nitrate from nitrate-rich vegetables in humans. Toxicol. Lett. 2008, 181, 177–181. [Google Scholar] [CrossRef]
- Li, H.; Samouilov, A.; Liu, X.; Zweier, J.L. Characterization of the magnitude and kinetics of xanthine oxidase-catalyzed nitrate reduction: Evaluation of its role in nitrite and nitric oxide generation in anoxic tissues. Biochemistry 2003, 42, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- McNally, B.; Griffin, J.L.; Roberts, L.D. Dietary inorganic nitrate: From villain to hero in metabolic disease? Mol. Nutr. Food Res. 2016, 60, 67–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bescos, R.; Rodriguez, F.A.; Iglesias, X.; Ferrer, M.D.; Iborra, E.; Pons, A. Acute administration of inorganic nitrate reduces VO(2peak) in endurance athletes. Med. Sci. Sports Exerc. 2011, 43, 1979–1986. [Google Scholar] [CrossRef] [Green Version]
- Larsen, F.J.; Weitzberg, E.; Lundberg, J.O.; Ekblom, B. Effects of dietary nitrate on oxygen cost during exercise. Acta Physiol. 2007, 191, 59–66. [Google Scholar] [CrossRef]
- Larsen, F.J.; Schiffer, T.A.; Borniquel, S.; Sahlin, K.; Ekblom, B.; Lundberg, J.O.; Weitzberg, E. Dietary inorganic nitrate improves mitochondrial efficiency in humans. Cell Metab. 2011, 13, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Bailey, S.J.; Fulford, J.; Vanhatalo, A.; Winyard, P.G.; Blackwell, J.R.; DiMenna, F.J.; Wilkerson, D.P.; Benjamin, N.; Jones, A.M. Dietary nitrate supplementation enhances muscle contractile efficiency during knee-extensor exercise in humans. J. Appl. Physiol. 2010, 109, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Peleli, M.; Zollbrecht, C.; Giulietti, A.; Terrando, N.; Lundberg, J.O.; Weitzberg, E.; Carlstrom, M. Inorganic nitrite attenuates NADPH oxidase-derived superoxide generation in activated macrophages via a nitric oxide-dependent mechanism. Free Radic. Biol. Med. 2015, 83, 159–166. [Google Scholar] [CrossRef]
- Steensberg, A.; Keller, C.; Hillig, T.; Frosig, C.; Wojtaszewski, J.F.; Pedersen, B.K.; Pilegaard, H.; Sander, M. Nitric oxide production is a proximal signaling event controlling exercise-induced mRNA expression in human skeletal muscle. FASEB J. 2007, 21, 2683–2694. [Google Scholar] [CrossRef]
- Baum, O.; Vieregge, M.; Koch, P.; Gul, S.; Hahn, S.; Huber-Abel, F.A.; Pries, A.R.; Hoppeler, H. Phenotype of capillaries in skeletal muscle of nNOS-knockout mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R1175–R1182. [Google Scholar] [CrossRef] [Green Version]
- Iglesias Bonilla, P.; Mayoral Sanchez, E.; Lapetra Peralta, J.; Iborra Oquendo, M.; Villalba Alcala, F.; Cayuela Dominguez, A. [Validation of two systems of self-measurement of blood pressure, the OMRON HEM-705 CP and OMRON M1 (HEM 422C2-E) models]. Aten. Primaria 2002, 30, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Capo, X.; Martorell, M.; Sureda, A.; Llompart, I.; Tur, J.A.; Pons, A. Diet supplementation with DHA-enriched food in football players during training season enhances the mitochondrial antioxidant capabilities in blood mononuclear cells. Eur. J. Nutr. 2015, 54, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Capo, X.; Martorell, M.; Sureda, A.; Tur, J.A.; Pons, A. Effects of docosahexaenoic supplementation and in vitro vitamin C on the oxidative and inflammatory neutrophil response to activation. Oxid. Med. Cell Longev. 2015, 187849, 19. [Google Scholar]
- Liddle, L.; Monaghan, C.; Burleigh, M.C.; McIlvenna, L.C.; Muggeridge, D.J.; Easton, C. Changes in body posture alter plasma nitrite but not nitrate concentration in humans. Nitric. Oxide 2018, 72, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Sureda, A.; Batle, J.M.; Capo, X.; Martorell, M.; Cordova, A.; Tur, J.A.; Pons, A. Scuba diving induces nitric oxide synthesis and the expression of inflammatory and regulatory genes of the immune response in neutrophils. Physiol. Genom. 2014, 46, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Shinde, D.D.; Kim, K.B.; Oh, K.S.; Abdalla, N.; Liu, K.H.; Bae, S.K.; Shon, J.H.; Kim, H.S.; Kim, D.H.; Shin, J.G. LC-MS/MS for the simultaneous analysis of arachidonic acid and 32 related metabolites in human plasma: Basal plasma concentrations and aspirin-induced changes of eicosanoids. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2012, 911, 113–121. [Google Scholar] [CrossRef]
- Yue, H.; Jansen, S.A.; Strauss, K.I.; Borenstein, M.R.; Barbe, M.F.; Rossi, L.J.; Murphy, E. A liquid chromatography/mass spectrometric method for simultaneous analysis of arachidonic acid and its endogenous eicosanoid metabolites prostaglandins, dihydroxyeicosatrienoic acids, hydroxyeicosatetraenoic acids, and epoxyeicosatrienoic acids in rat brain tissue. J. Pharm. Biomed. Anal. 2007, 43, 1122–1134. [Google Scholar]
- Koh, Y.; Park, J. Cell adhesion molecules and exercise. J. Inflamm. Res. 2018, 11, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Capo, X.; Martorell, M.; Sureda, A.; Tur, J.A.; Pons, A. Effects of dietary Docosahexaenoic, training and acute exercise on lipid mediators. J. Int. Soc. Sports Nutr. 2016, 13, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blobaum, A.L.; Marnett, L.J. Structural and functional basis of cyclooxygenase inhibition. J. Med. Chem. 2007, 50, 1425–1441. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, M.D.; Busquets-Cortes, C.; Capo, X.; Tejada, S.; Tur, J.A.; Pons, A.; Sureda, A. Cyclooxygenase-2 Inhibitors as a Therapeutic Target in Inflammatory Diseases. Curr. Med. Chem. 2019, 26, 3225–3241. [Google Scholar] [CrossRef]
- Trappe, T.A.; Liu, S.Z. Effects of prostaglandins and COX-inhibiting drugs on skeletal muscle adaptations to exercise. J. Appl. Physiol. (1985) 2013, 115, 909–919. [Google Scholar] [CrossRef] [Green Version]
- Carroll, C.C.; O’Connor, D.T.; Steinmeyer, R.; Del Mundo, J.D.; McMullan, D.R.; Whitt, J.A.; Ramos, J.E.; Gonzales, R.J. The influence of acute resistance exercise on cyclooxygenase-1 and -2 activity and protein levels in human skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R24–R30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wertz, P.W. Naturally occurring omega-Hydroxyacids. Int. J. Cosmet. Sci. 2018, 40, 31–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanders, R.J.; Komen, J.; Kemp, S. Fatty acid omega-oxidation as a rescue pathway for fatty acid oxidation disorders in humans. FEBS J. 2011, 278, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Butovich, I.A.; McMahon, A.; Wojtowicz, J.C.; Lin, F.; Mancini, R.; Itani, K. Dissecting lipid metabolism in meibomian glands of humans and mice: An integrative study reveals a network of metabolic reactions not duplicated in other tissues. Biochim. Biophys. Acta 2016, 1861, 538–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiva, S. Nitrite: A Physiological Store of Nitric Oxide and Modulator of Mitochondrial Function. Redox Biol. 2013, 1, 40–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlstrom, M.; Larsen, F.J.; Nystrom, T.; Hezel, M.; Borniquel, S.; Weitzberg, E.; Lundberg, J.O. Dietary inorganic nitrate reverses features of metabolic syndrome in endothelial nitric oxide synthase-deficient mice. Proc. Natl. Acad. Sci. USA 2010, 107, 17716–17720. [Google Scholar] [CrossRef] [Green Version]
- Vanhatalo, A.; Bailey, S.J.; Blackwell, J.R.; DiMenna, F.J.; Pavey, T.G.; Wilkerson, D.P.; Benjamin, N.; Winyard, P.G.; Jones, A.M. Acute and chronic effects of dietary nitrate supplementation on blood pressure and the physiological responses to moderate-intensity and incremental exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R1121–R1131. [Google Scholar] [CrossRef] [Green Version]
- Fulford, J.; Winyard, P.G.; Vanhatalo, A.; Bailey, S.J.; Blackwell, J.R.; Jones, A.M. Influence of dietary nitrate supplementation on human skeletal muscle metabolism and force production during maximum voluntary contractions. Pflug. Arch. 2013, 465, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Zweier, J.L.; Li, H.; Samouilov, A.; Liu, X. Mechanisms of nitrite reduction to nitric oxide in the heart and vessel wall. Nitric. Oxide 2010, 22, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Velmurugan, S.; Gan, J.M.; Rathod, K.S.; Khambata, R.S.; Ghosh, S.M.; Hartley, A.; Van Eijl, S.; Sagi-Kiss, V.; Chowdhury, T.A.; Curtis, M.; et al. Dietary nitrate improves vascular function in patients with hypercholesterolemia: A randomized, double-blind, placebo-controlled study. Am. J. Clin. Nutr. 2016, 103, 25–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundberg, J.O.; Carlstrom, M.; Weitzberg, E. Metabolic Effects of Dietary Nitrate in Health and Disease. Cell Metab. 2018, 28, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Wylie, L.J.; Mohr, M.; Krustrup, P.; Jackman, S.R.; Ermiotadis, G.; Kelly, J.; Black, M.I.; Bailey, S.J.; Vanhatalo, A.; Jones, A.M. Dietary nitrate supplementation improves team sport-specific intense intermittent exercise performance. Eur. J. Appl. Physiol. 2013, 113, 1673–1684. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Thompson, C.; Wylie, L.J.; Vanhatalo, A. Dietary Nitrate and Physical Performance. Annu. Rev. Nutr. 2018, 38, 303–328. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Patient | Age (Years) | Weight (kg) | Height (m) | BMI (kg/m2) | BP Syst/Diast (mmHg) | WC (cm) | HDLC (mg/dL) | TG (mg/dL) | Glucose (mg/dL) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 65 | 74.4 | 1.62 | 28,0 | 15.3/8.2 | 100 | 40 | 174 | 87 |
| 2 | 60 | 89.7 | 1.78 | 28,3 | 13.1/7.7 | 102 | 59 | 152 | 85 |
| 3 | 57 | 80.6 | 1.69 | 28.1 | 14.3/8.9 | 102 | 31 | 108 | 95 |
| 4 | 63 | 98.8 | 1.72 | 33.2 | 15.3/8.3 | 122 | 52 | 231 | 153 |
| 5 | 73 | 95.2 | 1.59 | 37.6 | 14.7/8.6 | 122 | 38 | 346 | 102 |
| 6 | 63 | 118.7 | 1.73 | 39.3 | 14.4/8.6 | 127 | 63 | 149 | 126 |
| 7 | 59 | 94.2 | 1.65 | 34.5 | 13.0/8.1 | 116 | 40 | 232 | 104 |
| 8 | 60 | 90.3 | 1.76 | 29.0 | 14.6/6.3 | 105 | 47 | 119 | 145 |
| 9 | 61 | 73.9 | 1.61 | 28.5 | 13.0/8.2 | 103 | 74 | 167 | 91 |
| 10 | 55 | 121.5 | 1.88 | 34.2 | 14.2/9.7 | 122 | 28 | 463 | 120 |
| 11 | 59 | 83.8 | 1.69 | 29.3 | 14.1/8.5 | 103 | 28 | 125 | 121 |
| 12 | 68 | 82.6 | 1.66 | 29.9 | 13.5/6.6 | 105 | 46 | 446 | 96 |
| 13 | 58 | 88.8 | 1.64 | 32.8 | 14.7/9.6 | 114 | 46 | 177 | 100 |
| 14 | 70 | 75.3 | 1.63 | 28.07 | 19.5/7.0 | 108 | 52 | 82 | 85 |
| Parameter | Placebo | Nitrate |
|---|---|---|
| Test duration (min) | 30.3 ± 0.1 | 30.5 ± 0.1 |
| Speed (km/h) | 4.4 ± 0.3 | 4.4 ± 0.3 |
| Heart Rate (Beats/min) | 103 ± 4.7 | 97.0 ± 3.5 * |
| VO2 (mL/min) | 1408 ± 79 | 1245 ± 73 * |
| VO2 (mL/kg min) | 16.2 ± 1.0 | 14.4 ± 1.0 * |
| VCO2 (mL/min) | 1259 ± 69 | 1110 ± 61 * |
| Energy Expenditure (kcal/min) | 6.9 ± 0.4 | 6.1 ± 0.4 * |
| Total Energy Expenditure (kcal) | 210 ± 12 | 187 ± 11 * |
| Energy Efficiency (kcal/km) | 97.6 ± 9.7 | 85.7 ± 5.6 * |
| Respiratory exchange ratio | 0.89 ± 0.01 | 0.89 ±0.006 |
| Energy from lipids (%) | 37.3 ± 3.6 | 38.3 ± 2.1 |
| NOx | Placebo | Supplemented | |||
|---|---|---|---|---|---|
| Pre-Exercise | Post-Exercise | Pre- Exercise | Post-Exercise | ANOVA | |
| Nitrate (µM) | 54.5 ± 11.6 | 53.8 ± 14.1 | 26.2 ± 6.1 | 261 ± 17 *# | E, ExS |
| Nitrite (µM) | 0.14 ± 0.01 | 0.12 ± 0.01 | 0.13 ± 0.02 | 0.24 ± 0.05 *# | E’, ExS’ |
| Nitrite/Nitrate Ratio (×100) | 0.36 ± 0.11 | 0.29 ± 0.06 | 0.57 ± 0.12 | 0.09 ± 0.1 *# | E |
| Parameter | Placebo | Supplemented | |||
|---|---|---|---|---|---|
| Pre-Exercise | Post-Exercise | Pre-Exercise | Post-Exercise | ANOVA | |
| Neutrophils (103/mm3) a | 4.00 ± 0.58 | 4.78 ± 0.57 | 3.81 ± 0.29 | 3.89 ± 0.08 | |
| PBMCs (103/mm3) a | 3.78 ± 0.45 | 3.09 ± 0.50 | 3.76 ± 0.35 | 3.41 ± 0.55 | |
| TNFα (pg/mL) b | 198 (283) | 248 (332) *’ | 212 (158) | 176 (137) | N/A |
| IL6 (pg/mL) a | 6.82 ± 0.64 | 6.85 ± 0.54 | 6.11 ± 0.20 | 6.54 ± 0.53 | |
| ICAM1 (pg/mL) a | 298 ± 20 | 387 ± 31 * | 300 ± 21 | 306 ± 30 | E |
| C Reactive Protein (mg/dL) a | 0.30 ± 0.20 | 0.29 ± 0.20 | 0.21 ± 0.09 | 0.21 ± 0.09 | |
| Compound | Placebo | Supplemented | ANOVA | ||
|---|---|---|---|---|---|
| Pre-Exercise | Post-Exercise | Pre-Exercise | Post-Exercise | ||
| AA (nM) a | 1939 ± 174 | 2073 ± 142 | 1897 ± 367 | 1883 ± 312 | |
| ETA (nM) a | 20.3 ± 2.4 | 24.0 ± 5.1 | 15.8 ± 3.0 | 18.6 ± 3.3 | |
| 5-HETE (nM) a | 31.5 ± 3.6 | 42.0 ± 6.1 | 30.2 ± 6.1 | 33.6 ± 5.0 | |
| 15-HETE (nM) a | 8.54 ± 1.07 | 9.14 ± 1.11 | 6.69 ± 1.20 | 7.22 ± 0.98 | |
| 20-HETE (nM) a | 2.35 ± 0.36 | 2.62 ± 0.58 | 2.37 ± 0.79 | 1.94 ± 0.65 | |
| PGE2 (nM) b | 30.6 (13.3) | 95 (157) *’ | 31.5 (17.7) | 36.8 (26.0) | N/A |
| EPA (nM) a | 453 ± 80 | 450 ± 45 | 342 ± 90 | 435 ± 113 | |
| 15-HEPE (nM) a | 6.02 ± 0.40 | 8.92 ± 1.41 | 4.46 ± 0.76 | 5.12 ± 0.59 # | E, S |
| DHA (nM) a | 5263 ± 842 | 4832 ± 632 | 5153 ± 682 | 4216 ± 1045 | |
| 17-DoHE (nM) a | 13.3 ± 2.1 | 15.6 ± 2.0 | 11.4 ± 1.4 | 9.83 ± 1.12 | |
| RvD2 (nM) a | 0.38 ± 0.09 | 0.29 ± 0.10 | 0.39 ± 0.15 | 0.40 ± 0.13 | |
| 16-HPAL (nM) a | 29.6 ± 2.8 | 66.2 ± 10.0 * | 35.7 ± 4.3 | 37.3 ± 6.5 #’ | E, ExS |
| PGE1 (nM) a | 36.3 ± 5.7 | 50.4 ± 8.9 * | 31.2 ± 2.7 | 31.9 ± 1.1 | E |
| Compound | PBMCs | NEUTROPHILS |
|---|---|---|
| TNFα (pg/mL) a | 301 ± 38 | 127 ± 26 * |
| IL6 (pg/mL) b | 92.9 (84.9) | 24.9 (21.2) * |
| AA (nM) b | 12.1 (6.5) | 6.16 (14.9) |
| ETA (nM) b | 0.040 (0.095) | 0.016 (0.154) |
| 5-HETE (nM) b | 1.85 (2.66) | 0.120 (0.567) * |
| 15-HETE (nM) b | 1.29 (1.98) | 0.13 (1.30) * |
| PGE2 (nM) b | 0.607 (0.82) | 4.52 (4.55) * |
| EPA (nM) b | 3.41 (4.07) | 4.25 (41.4) |
| 15-HEPE (nM) b | 0.17 (1.21) | 0.000 (0.049) * |
| DHA (nM) b | 4.95 (3.14) | 1.03 (1.49) * |
| 17-DoHE (nM) b | 1.60 (9.64) | 0.000 (0.006) * |
| 16-HPAL (nM) b | 24.5 (15.7) | 3.10 (4.98) * |
| PGE1 (nM) b | 0.030 (0.052) | 0.000 (0.031) * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capó, X.; Ferrer, M.D.; Olek, R.A.; Salaberry, E.; Suau, R.; Marí, B.; Llompart, I.; Tur, J.A.; Sureda, A.; Pons, A. Oral Administration of Sodium Nitrate to Metabolic Syndrome Patients Attenuates Mild Inflammatory and Oxidative Responses to Acute Exercise. Antioxidants 2020, 9, 596. https://doi.org/10.3390/antiox9070596
Capó X, Ferrer MD, Olek RA, Salaberry E, Suau R, Marí B, Llompart I, Tur JA, Sureda A, Pons A. Oral Administration of Sodium Nitrate to Metabolic Syndrome Patients Attenuates Mild Inflammatory and Oxidative Responses to Acute Exercise. Antioxidants. 2020; 9(7):596. https://doi.org/10.3390/antiox9070596
Chicago/Turabian StyleCapó, Xavier, Miguel D. Ferrer, Robert A. Olek, Eduardo Salaberry, Rafael Suau, Bartolomé Marí, Isabel Llompart, Josep A. Tur, Antoni Sureda, and Antoni Pons. 2020. "Oral Administration of Sodium Nitrate to Metabolic Syndrome Patients Attenuates Mild Inflammatory and Oxidative Responses to Acute Exercise" Antioxidants 9, no. 7: 596. https://doi.org/10.3390/antiox9070596
APA StyleCapó, X., Ferrer, M. D., Olek, R. A., Salaberry, E., Suau, R., Marí, B., Llompart, I., Tur, J. A., Sureda, A., & Pons, A. (2020). Oral Administration of Sodium Nitrate to Metabolic Syndrome Patients Attenuates Mild Inflammatory and Oxidative Responses to Acute Exercise. Antioxidants, 9(7), 596. https://doi.org/10.3390/antiox9070596