Enzymatic Antioxidant Signatures in Hyperthermophilic Archaea





Abstract
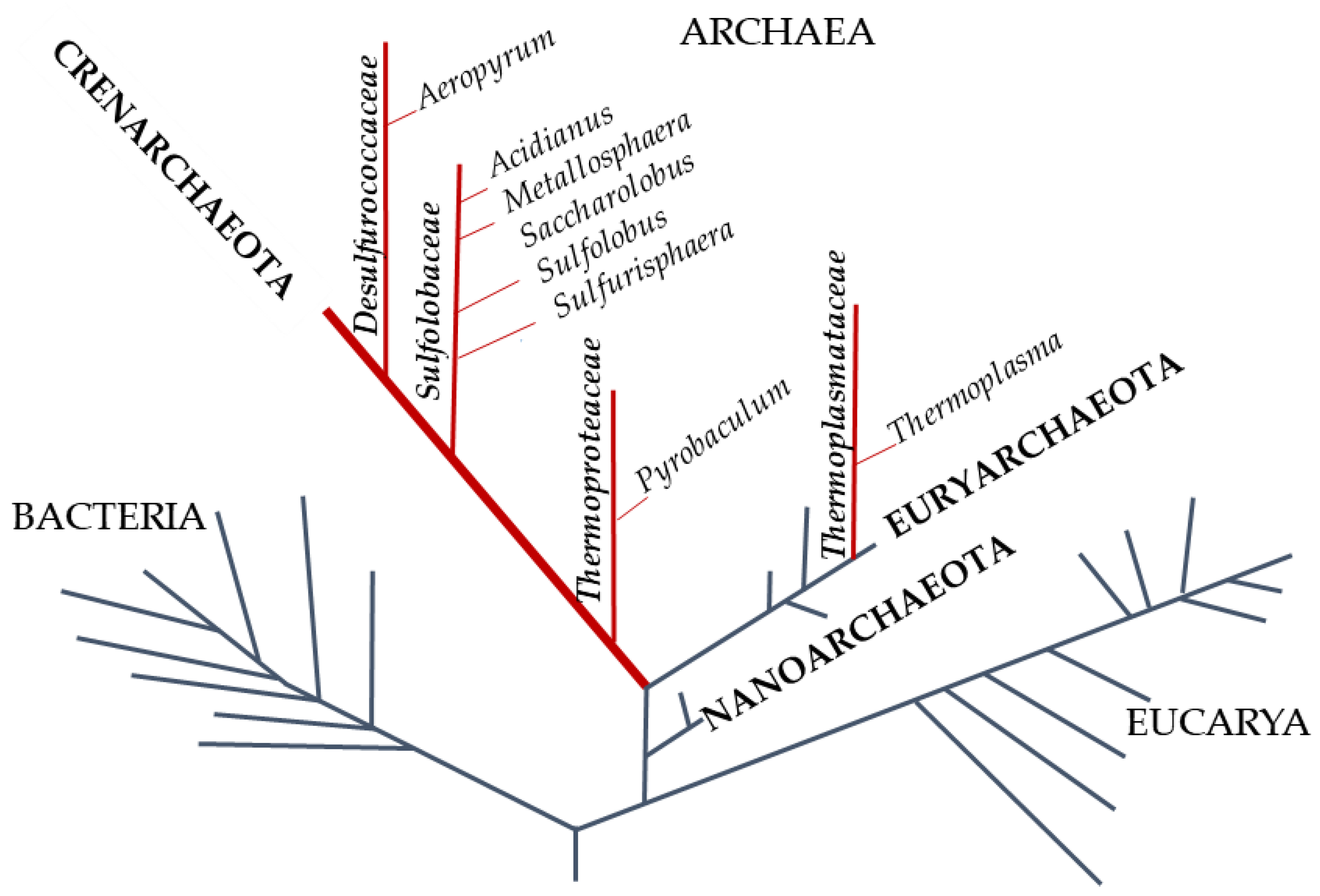
:1. Introduction
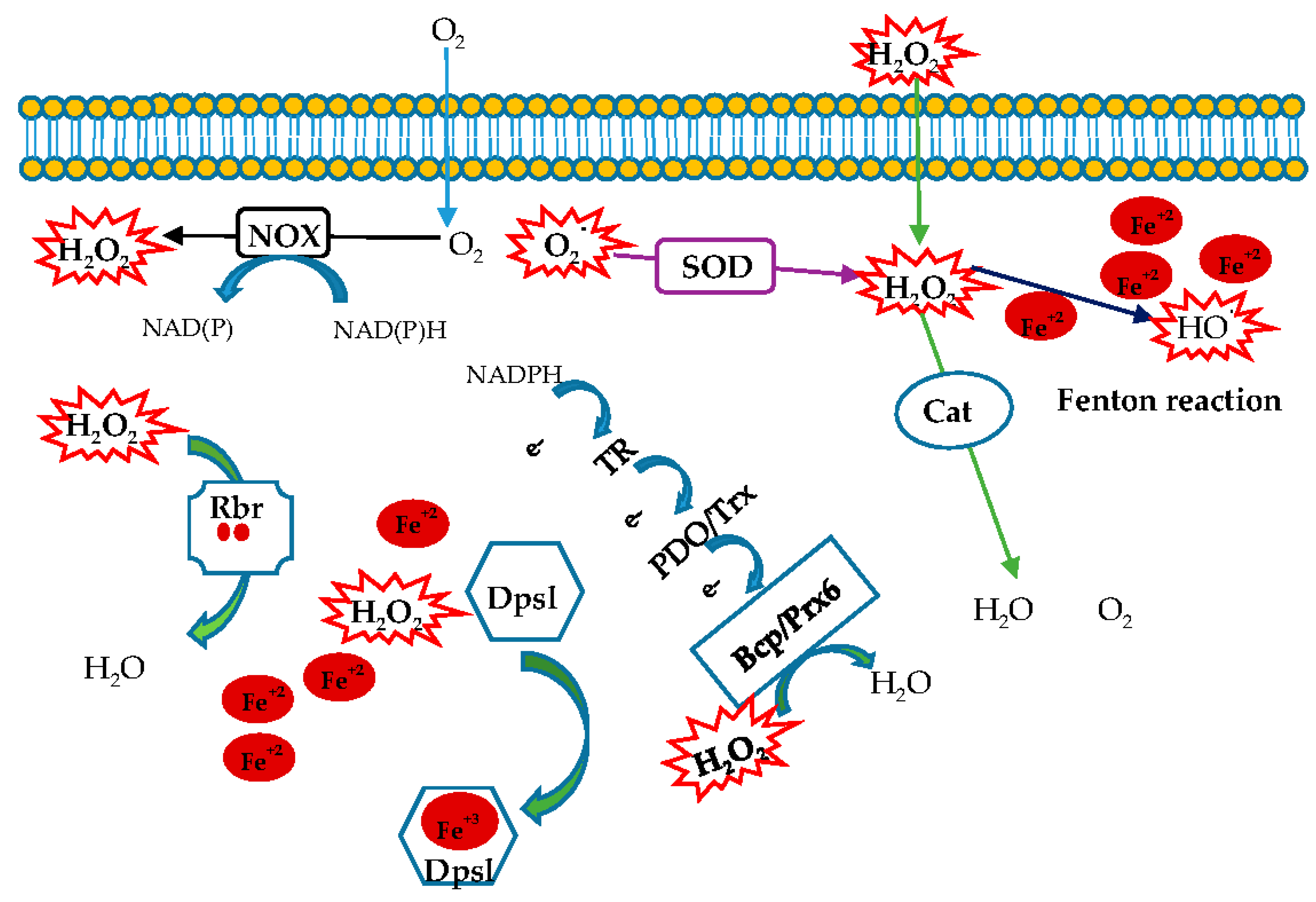
2. Superoxide Anion Scavengers
3. H2O2 Production
4. H2O2 Targets
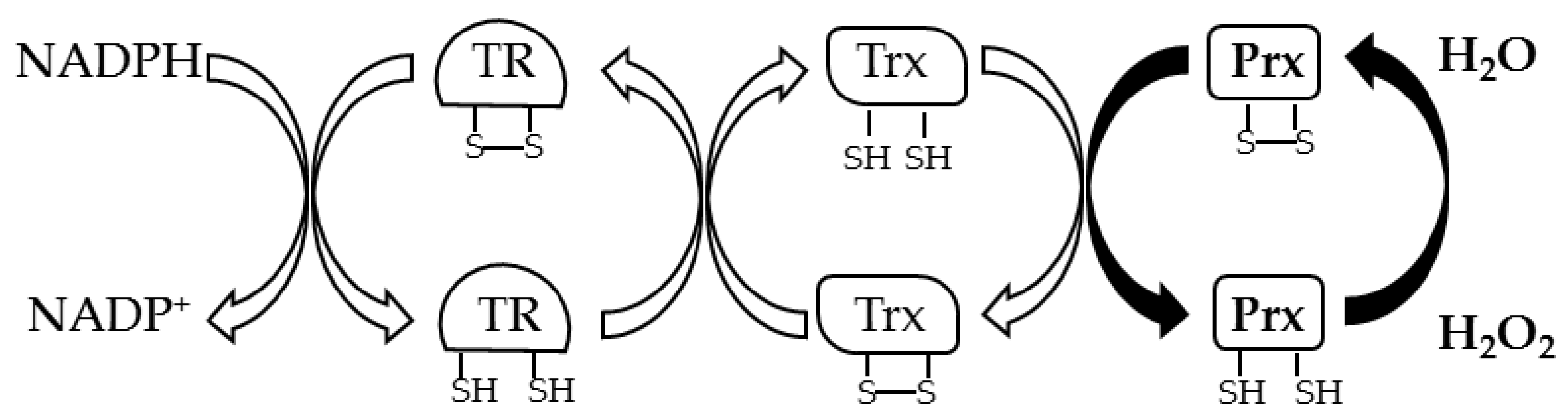
5. The Main Peroxide Scavengers in Hyperthermophilic Aerobic/Microaerophilic Archaea: Prxs
5.1. Prxs in Sulfolobaceae Family
5.2. Prxs in the Desulfurococcaceae Family
6. Other Enzymes Involved in the Cellular Protection against Oxidative Stress
6.1. DNA-Binding Protein from Starved Cells (Dps)
6.2. Rubrerythrin (Rbr)
7. Biotechnological Applications
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Lemire, J.; AlHasawi, A.; Appanna, V.P.; Tharmalingam, S.; Appanna, V. Metabolic defence against oxidative stress: The road less travelled so far. J. Appl. Microbiol. 2017, 123, 798–809. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.-F. Superoxide dismutases: Ancient enzymes and new insights. FEBS Lett. 2011, 586, 585–595. [Google Scholar] [CrossRef] [Green Version]
- Zamocky, M.; Gasselhuber, B.; Furtmüller, P.; Obinger, C. Molecular evolution of hydrogen peroxide degrading enzymes. Arch. Biochem. Biophys. 2012, 525, 131–144. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.G. Overview on Peroxiredoxin. Mol. Cells 2016, 39, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Hall, A.; Karplus, P.; Poole, L.B. Typical 2-Cys peroxiredoxins--structures, mechanisms and functions. FEBS J. 2009, 276, 2469–2477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DePonte, M. Glutathione catalysis and the reaction mechanisms of glutathione-dependent enzymes. Biochim. Biophys. Acta (BBA) Gen. Subj. 2013, 1830, 3217–3266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanschmann, E.-M.; Godoy, J.R.; Berndt, C.; Hudemann, C.; Lillig, C.H. Thioredoxins, Glutaredoxins, and Peroxiredoxins—Molecular Mechanisms and Health Significance: From Cofactors to Antioxidants to Redox Signaling. Antioxid. Redox Signal 2013, 19, 1539–1605. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free. Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef]
- Woese, C.R.; Kandler, O.; Wheelis, M.L. Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Natl. Acad. Sci. USA 1990, 87, 4576–4579. [Google Scholar] [CrossRef] [Green Version]
- Contursi, P.; Fusco, S.; Limauro, D.; Fiorentino, G. Host and viral transcriptional regulators in Sulfolobus: An overview. Extremophiles 2013, 17, 881–895. [Google Scholar] [CrossRef]
- Chaban, B.; Ng, S.Y.; Jarrell, K.F. Archaeal habitats—from the extreme to the ordinary. Can. J. Microbiol. 2006, 52, 73–116. [Google Scholar] [CrossRef] [PubMed]
- Stetter, K.O. Hyperthermophilic procaryotes. FEMS Microbiol. Rev. 1996, 18, 149–158. [Google Scholar] [CrossRef]
- Stetter, K.O. Hyperthermophiles in the history of life. Philos. Trans. R. Soc. B: Biol. Sci. 2006, 361, 1837–1843. [Google Scholar] [CrossRef] [PubMed]
- Jenney, F.E.; Verhagen, M.F.; Cui, X.; Adams, M.W. Anaerobic Microbes: Oxygen Detoxification Without Superoxide Dismutase. Science 1999, 286, 306–309. [Google Scholar] [CrossRef]
- Grunden, A.M.; Jenney, F.E.; Ma, K.; Ji, M.; Weinberg, M.V.; Adams, M.W.W. In Vitro Reconstitution of an NADPH-Dependent Superoxide Reduction Pathway from Pyrococcus Furiosus. Appl. Environ. Microbiol. 2005, 71, 1522–1530. [Google Scholar] [CrossRef] [Green Version]
- Adams, M.W.; Jenney, F.E.; Clay, M.D.; Johnson, M.K. Superoxide reductase: Fact or fiction? JBIC J. Biol. Inorg. Chem. 2002, 7, 647–652. [Google Scholar] [CrossRef]
- Cannio, R.; D’Angelo, A.; Rossi, M.; Bartolucci, S. A superoxide dismutase from the archaeon Sulfolobus Solfataricus is an extracellular enzyme and prevents the deactivation by superoxide of cell-bound proteins. JBIC J. Biol. Inorg. Chem. 2000, 267, 235–243. [Google Scholar] [CrossRef]
- Russo, A.D.; Rullo, R.; Nitti, G.; Masullo, M.; Bocchini, V. Iron superoxide dismutase from the archaeon Sulfolobus solfataricus: Average hydrophobicity and amino acid weight are involved in the adaptation of proteins to extreme environments. Biochim. Biophys. Acta (BBA) Bioenerg. 1997, 1343, 23–30. [Google Scholar] [CrossRef]
- Case, A.J. On the Origin of Superoxide Dismutase: An Evolutionary Perspective of Superoxide-Mediated Redox Signaling. Antioxidants 2017, 6, 82. [Google Scholar] [CrossRef] [Green Version]
- Youn, H.-D.; Kim, E.-J.; Roe, J.-H.; Hah, Y.C.; Kang, S.-O. A novel nickel-containing superoxide dismutase from Streptomyces spp. Biochem. J. 1996, 318, 889–896. [Google Scholar] [CrossRef] [Green Version]
- Dupont, C.L.; Neupane, K.; Shearer, J.; Palenik, B. Diversity, function and evolution of genes coding for putative Ni-containing superoxide dismutases. Environ. Microbiol. 2008, 10, 1831–1843. [Google Scholar] [CrossRef] [PubMed]
- Wolfe-Simon, F.; Grzebyk, D.; Schofield, O.; Falkowski, P.G. The Role and Evolution of Superoxide Dismutases in Algae1. J. Phycol. 2005, 41, 453–465. [Google Scholar] [CrossRef]
- Wuerges, J.; Lee, J.-W.; Yim, Y.-I.; Yim, H.-S.; Kang, S.-O.; Carugo, K.D. Crystal structure of nickel-containing superoxide dismutase reveals another type of active site. Proc. Natl. Acad. Sci. USA 2004, 101, 8569–8574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedone, E.M. Sensing and adapting to environmental stress: The archaeal tactic. Front. Biosci. 2004, 9, 2909. [Google Scholar] [CrossRef] [Green Version]
- Contursi, P. Responding to toxic compounds: A genomic and functional overview of Archaea. Front. Biosci. 2013, 18, 165–189. [Google Scholar] [CrossRef] [Green Version]
- Ursby, T.; Adinolfi, B.S.; Al-Karadaghi, S.; De Vendittis, E.; Bocchini, V. Iron superoxide dismutase from the archaeon Sulfolobus Solfataricus: Analysis of structure and thermostability. J. Mol. Biol. 1999, 286, 189–205. [Google Scholar] [CrossRef]
- Lim, J.-H.; Yu, Y.G.; Han, Y.S.; Cho, S.-J.; Ahn, B.-Y.; Kim, S.-H.; Cho, Y. The crystal structure of an Fe-superoxide dismutase from the hyperthermophile Aquifex Pyrophilus at 1.9 A resolution: Structural basis for thermostability. J. Mol. Biol. 1997, 270, 259–274. [Google Scholar] [CrossRef]
- Gogliettino, M.A.; Tanfani, F.; Sciré, A.; Ursby, T.; Adinolfi, B.S.; Cacciamani, T.; De Vendittis, E. The Role of Tyr41 and His155 in the Functional Properties of Superoxide Dismutase from the Archaeon Sulfolobus Solfataricus. Biochemistry 2004, 43, 2199–2208. [Google Scholar] [CrossRef]
- De Vendittis, E.; Ursby, T.; Rullo, R.; Gogliettino, M.A.; Masullo, M.; Bocchini, V. Phenylmethanesulfonyl fluoride inactivates an archaeal superoxide dismutase by chemical modification of a specific tyrosine residue. JBIC J. Biol. Inorg. Chem. 2001, 268, 1794–1801. [Google Scholar] [CrossRef]
- Amo, T.; Atomi, H.; Imanaka, T. Biochemical Properties and Regulated Gene Expression of the Superoxide Dismutase from the Facultatively Aerobic Hyperthermophile Pyrobaculum calidifontis. J. Bacteriol. 2003, 185, 6340–6347. [Google Scholar] [CrossRef] [Green Version]
- Wheaton, G.H.; Mukherjee, A.; Kelly, R.M. Transcriptomes of the Extremely Thermoacidophilic Archaeon Metallosphaera Sedula Exposed to Metal “Shock” Reveal Generic and Specific Metal Responses. Appl. Environ. Microbiol. 2016, 82, 4613–4627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittaker, M.M.; Whittaker, J.W. Recombinant superoxide dismutase from a hyperthermophilic archaeon, Pyrobaculum aerophilium. JBIC J. Biol. Inorg. Chem. 2000, 5, 402–408. [Google Scholar] [PubMed]
- Yamano, S.; Sako, Y.; Nomura, N.; Maruyama, T. A cambialistic SOD in a strictly aerobic hyperthermophilic archaeon, Aeropyrum Pernix. J. Biochem. 1999, 126, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Imlay, J.A. Why do bacteria use so many enzymes to scavenge hydrogen peroxide? Arch. Biochem. Biophys. 2012, 525, 145–160. [Google Scholar] [CrossRef] [Green Version]
- Cabiscol, E.; Tamarit, J.; Ros, J. Oxidative stress in bacteria and protein damage by reactive oxygen species. Int. Microbiol. 2000, 3, 3–8. [Google Scholar]
- Imlay, J.A. Pathways of Oxidative Damage. Annu. Rev. Microbiol. 2003, 57, 395–418. [Google Scholar] [CrossRef]
- Kobori, H.; Ogino, M.; Orita, I.; Nakamura, S.; Imanaka, T.; Fukui, T. Characterization of NADH Oxidase/NADPH Polysulfide Oxidoreductase and Its Unexpected Participation in Oxygen Sensitivity in an Anaerobic Hyperthermophilic Archaeon. J. Bacteriol. 2010, 192, 5192–5202. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, Y.; Massey, V.; Takeda, K.; Kawasaki, S.; Sato, J.; Watanabe, T.; Niimura, Y. Hydrogen Peroxide-Forming NADH Oxidase Belonging to the Peroxiredoxin Oxidoreductase Family: Existence and Physiological Role in Bacteria. J. Bacteriol. 2001, 183, 2431–2438. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Park, S.-C.; Lee, S.; Pham, B.P.; Yu, R.; Le, T.L.; Han, S.W.; Yang, J.-K.; Choi, M.-S.; Baumeister, W.; et al. Hexameric ring structure of a thermophilic archaeon NADH oxidase that produces predominantly H2O. FEBS J. 2008, 275, 5355–5366. [Google Scholar] [CrossRef]
- Aguirre, J.; Lambeth, J.D. Nox enzymes from fungus to fly to fish and what they tell us about Nox function in mammals. Free Radic. Biol. Med. 2010, 49, 1342–1353. [Google Scholar] [CrossRef] [Green Version]
- Kengen, S.W.M.; Van Der Oost, J.; De Vos, W.M. Molecular characterization of H2O2-forming NADH oxidases from Archaeoglobus fulgidus. JBIC J. Biol. Inorg. Chem. 2003, 270, 2885–2894. [Google Scholar] [CrossRef] [PubMed]
- Arai, T.; Kimata, S.; Mochizuki, D.; Hara, K.; Zako, T.; Odaka, M.; Yohda, M.; Arisaka, F.; Kanamaru, S.; Matsumoto, T.; et al. NADH oxidase and alkyl hydroperoxide reductase subunit C (peroxiredoxin) from Amphibacillus Xylanus form an oligomeric assembly. FEBS Open Bio 2015, 5, 124–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, D.E.; Donnelly, C.J.; Mullendore, M.E.; Van Der Oost, J.; De Vos, W.M.; Iii, E.J.C. The NADH oxidase from Pyrococcus Furiosus. JBIC J. Biol. Inorg. Chem. 2001, 268, 5816–5823. [Google Scholar] [CrossRef]
- Jia, B.; Lee, S.; Pham, B.P.; Liu, J.; Pan, H.; Zhang, S.; Cheong, G.-W. Oxidized NADH Oxidase Inhibits Activity of an ATP/NAD Kinase from a Thermophilic Archaeon. Protein J. 2010, 29, 609–616. [Google Scholar] [CrossRef]
- Arcari, P.; Masullo, L.; Masullo, M.; Catanzano, F.; Bocchini, V. A NAD(P)H oxidase isolated from the archaeon Sulfolobus Solfataricus is not homologous with another NADH oxidase present in the same microorganism. Biochemical characterization of the enzyme and cloning of the encoding gene. J. Biol. Chem. 2000, 275, 895–900. [Google Scholar] [CrossRef] [Green Version]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.-F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Genet. 2017, 15, 385–396. [Google Scholar] [CrossRef]
- Maupin-Furlow, J.A. Methionine Sulfoxide Reductases of Archaea. Antioxidants 2018, 7, 124. [Google Scholar] [CrossRef] [Green Version]
- Ezraty, B.; Aussel, L.; Barras, F. Methionine sulfoxide reductases in prokaryotes. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2005, 1703, 221–229. [Google Scholar] [CrossRef]
- Tarrago, L.; Laugier, E.; Zaffagnini, M.; Marchand, C.H.; Le Maréchal, P.; Rouhier, N.; Lemaire, S.; Rey, P. Regeneration Mechanisms of Arabidopsis thaliana Methionine Sulfoxide Reductases B by Glutaredoxins and Thioredoxins. J. Biol. Chem. 2009, 284, 18963–18971. [Google Scholar] [CrossRef] [Green Version]
- Guan, N.; Li, J.; Du, G.; Chen, J.; Liu, L.; Shin, H.-D. Microbial response to environmental stresses: From fundamental mechanisms to practical applications. Appl. Microbiol. Biotechnol. 2017, 101, 3991–4008. [Google Scholar] [CrossRef]
- Shima, S.; Netrusov, A.I.; Sordel, M.; Wicke, M.; Hartmann, G.C.; Thauer, R.K. Purification, characterization, and primary structure of a monofunctional catalase from Methanosarcina Barkeri. Arch. Microbiol. 1999, 171, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Shima, S.; Sordel-Klippert, M.; Brioukhanov, A.L.; Netrusov, A.; Linder, D.; Thauer, R.K. Characterization of a Heme-Dependent Catalase from Methanobrevibacter Arboriphilus. Appl. Environ. Microbiol. 2001, 67, 3041–3045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown-Peterson, N.J.; Salin, M.L. Purification of a catalase-peroxidase from Halobacterium halobium: Characterization of some unique properties of the halophilic enzyme. J. Bacteriol. 1993, 175, 4197–4202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kengen, S.W.M.; Bikker, F.J.; Hagen, W.; Vos, W.M.; Oost, J. Characterization of a catalase-peroxidase from the hyperthermophilic archaeon Archaeoglobus Fulgidus. Extremophiles 2001, 5, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Sensen, C.W.; Charlebois, R.L.; Chow, C.; Clausen, I.G.; Curtis, B.; Doolittle, W.F.; Duguet, M.; Erauso, G.; Gaasterland, T.; Garrett, R.A.; et al. Completing the sequence of the Sulfolobus Solfataricus P2 genome. Extremophiles 1998, 2, 305–312. [Google Scholar] [CrossRef]
- Kawarabayasi, Y.; Hino, Y.; Horikawa, H.; Jin-No, K.; Takahashi, M.; Sekine, M.; Baba, S.-I.; Ankai, A.; Kosugi, H.; Hosoyama, A.; et al. Complete Genome Sequence of an Aerobic Thermoacidophilic Crenarchaeon, Sulfolobus Tokodaii strain7. DNA Res. 2001, 8, 123–140. [Google Scholar] [CrossRef] [Green Version]
- Kawarabayasi, Y.; Hino, Y.; Horikawa, H.; Yamazaki, S.; Haikawa, Y.; Jin-No, K.; Takahashi, M.; Sekine, M.; Baba, S.-I.; Ankai, A.; et al. Complete Genome Sequence of an Aerobic Hyper-thermophilic Crenarchaeon, Aeropyrum Pernix K1. DNA Res. 1999, 6, 83–101. [Google Scholar] [CrossRef]
- Ruepp, A.; Graml, W.; Santos, L.; Koretke, K.K.; Volker, C.; Mewes, H.-W.; Frishman, D.; Stocker, S.; Lupas, A.N.; Baumeister, W. The genome sequence of the thermoacidophilic scavenger Thermoplasma Acidophilum. Nature 2000, 407, 508–513. [Google Scholar] [CrossRef]
- Santos, L.; Frickey, T.; Peters, J.; Baumeister, W.; Lupas, A.; Zwickl, P. Thermoplasma Acidophilum TAA43 is an archaeal member of the eukaryotic meiotic branch of AAA ATPases. Biol. Chem. 2004, 385, 1105–1111. [Google Scholar] [CrossRef]
- Fitz-Gibbon, S.T.; Ladner, H.; Kim, U.-J.; Stetter, K.O.; Simon, M.I.; Miller, J.H. Genome sequence of the hyperthermophilic crenarchaeon Pyrobaculum Aerophilum. Proc. Natl. Acad. Sci. USA 2002, 99, 984–989. [Google Scholar] [CrossRef] [Green Version]
- Cozen, A.E.; Weirauch, M.T.; Pollard, K.S.; Bernick, D.L.; Stuart, J.M.; Lowe, T.M. Transcriptional Map of Respiratory Versatility in the Hyperthermophilic Crenarchaeon Pyrobaculum Aerophilum. J. Bacteriol. 2008, 191, 782–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amo, T.; Atomi, H.; Imanaka, T. Unique Presence of a Manganese Catalase in a Hyperthermophilic Archaeon, Pyrobaculum Calidifontis VA1. J. Bacteriol. 2002, 184, 3305–3312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, S.-J.; Ishikawa, K. Characterization of Novel Hexadecameric Thioredoxin Peroxidase from Aeropyrum Pernix K1. J. Biol. Chem. 2003, 278, 24174–24180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soito, L.; Williamson, C.; Knutson, S.T.; Fetrow, J.S.; Poole, L.B.; Nelson, K.J. PREX: PeroxiRedoxin classification indEX, a database of subfamily assignments across the diverse peroxiredoxin family. Nucleic Acids Res. 2010, 39, D332–D337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limauro, D.; Pedone, E.M.; Galdi, I.; Bartolucci, S. Peroxiredoxins as cellular guardians in Sulfolobus solfataricus—Characterization of Bcp1, Bcp3 and Bcp4. FEBS J. 2008, 275, 2067–2077. [Google Scholar] [CrossRef] [PubMed]
- Limauro, D.; Pedone, E.M.; Pirone, L.; Bartolucci, S. Identification and characterization of 1-Cys peroxiredoxin from Sulfolobus solfataricus and its involvement in the response to oxidative stress. FEBS J. 2006, 273, 721–731. [Google Scholar] [CrossRef]
- Stroobants, S.; Van Molle, I.; Saidi, Q.; Jonckheere, K.; Maes, D.; Peeters, E. Structure of the Prx6-subfamily 1-Cys peroxiredoxin from Sulfolobus islandicus. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2019, 75, 428–434. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Jia, B.; Liu, J.; Pham, B.P.; Kwak, J.M.; Xuan, Y.H.; Cheong, G.-W. A 1-Cys Peroxiredoxin from a Thermophilic Archaeon Moonlights as a Molecular Chaperone to Protect Protein and DNA against Stress-Induced Damage. PLoS ONE 2015, 10, e0125325. [Google Scholar] [CrossRef]
- D’Ambrosio, K.; Limauro, D.; Pedone, E.M.; Galdi, I.; Pedone, C.; Bartolucci, S.; De Simone, G. Insights into the catalytic mechanism of the Bcp family: Functional and structural analysis of Bcp1 from Sulfolobus solfataricus. Proteins Struct. Funct. Bioinform. 2009, 76, 995–1006. [Google Scholar] [CrossRef]
- Limauro, D.; D’Ambrosio, K.; Langella, E.; De Simone, G.; Galdi, I.; Pedone, C.; Pedone, E.; Bartolucci, S. Exploring the catalytic mechanism of the first dimeric Bcp: Functional, structural and docking analyses of Bcp4 from Sulfolobus solfataricus. Biochimie 2010, 92, 1435–1444. [Google Scholar] [CrossRef]
- Pedone, E.M.; Limauro, D.; D’Alterio, R.; Rossi, M.; Bartolucci, S. Characterization of a multifunctional protein disulfide oxidoreductase from Sulfolobus solfataricus. FEBS J. 2006, 273, 5407–5420. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Tibbelin, G.; De Pascale, D.; Rossi, M.; Bartolucci, S.; Ladenstein, R. A protein disulfide oxidoreductase from the archaeon Pyrococcus furiosus contains two thioredoxin fold units. Nat. Genet. 1998, 5, 602–611. [Google Scholar] [CrossRef]
- Pedone, E.; Limauro, D.; Bartolucci, S. The Machinery for Oxidative Protein Folding in Thermophiles. Antioxid. Redox Signal. 2008, 10, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Jorda, J.; Yeates, T.O. Widespread Disulfide Bonding in Proteins from Thermophilic Archaea. Archaea 2011, 1472–3646, 409156. [Google Scholar] [CrossRef]
- Beeby, M.; O’Connor, B.D.; Ryttersgaard, C.; Boutz, D.R.; Perry, L.J.; Yeates, T.O. The genomics of disulfide bonding and protein stabilization in thermophiles. PLoS Biol. 2005, 3, e309. [Google Scholar] [CrossRef] [Green Version]
- Limauro, D.; Saviano, M.; Galdi, I.; Rossi, M.; Bartolucci, S.; Pedone, E.M. Sulfolobus solfataricus protein disulphide oxidoreductase: Insight into the roles of its redox sites. Protein Eng. Des. Sel. 2008, 22, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, X.; Wang, L. Protein disulfide–isomerase, a folding catalyst and a redox-regulated chaperone. Free. Radic. Biol. Med. 2015, 83, 305–313. [Google Scholar] [CrossRef]
- Jeon, S.-J.; Ishikawa, K. Identification and characterization of thioredoxin and thioredoxin reductase from Aeropyrum pernix K1. JBIC J. Biol. Inorg. Chem. 2002, 269, 5423–5430. [Google Scholar] [CrossRef]
- Mizohata, E.; Sakai, H.; Fusatomi, E.; Terada, T.; Murayama, K.; Shirouzu, M.; Yokoyama, S. Crystal Structure of an Archaeal Peroxiredoxin from the Aerobic Hyperthermophilic Crenarchaeon Aeropyrum pernix K1. J. Mol. Biol. 2005, 354, 317–329. [Google Scholar] [CrossRef]
- Ebrahimi, K.H.; Hagedoorn, P.; Van Der Weel, L.; Verhaert, P.; Hagen, W. A novel mechanism of iron-core formation by Pyrococcus furiosus archaeoferritin, a member of an uncharacterized branch of the ferritin-like superfamily. JBIC J. Biol. Inorg. Chem. 2012, 17, 975–985. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S.C. The Ferritin-like superfamily: Evolution of the biological iron storeman from a rubrerythrin-like ancestor. Biochim. Biophys. Acta (BBA) Gen. Subj. 2010, 1800, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Zeth, K. Dps biomineralizing proteins: Multifunctional architects of nature. Biochem. J. 2012, 445, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.; Filman, D.J.; Finkel, S.; Kolter, R.; Hogle, J.M. The crystal structure of Dps, a ferritin homolog that binds and protects DNA. Nat. Genet. 1998, 5, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Pesek, J.; Büchler, R.; Albrecht, R.; Boland, W.; Zeth, K. Structure and Mechanism of Iron Translocation by a Dps Protein from Microbacterium arborescens. J. Biol. Chem. 2011, 286, 34872–34882. [Google Scholar] [CrossRef] [Green Version]
- Minato, T.; Teramoto, T.; Kakuta, Y.; Ogo, S.; Yoon, K. Biochemical and structural characterization of a thermostable Dps protein with His-type ferroxidase centers and outer metal-binding sites. FEBS Open Bio 2020. [Google Scholar] [CrossRef] [Green Version]
- Haikarainen, T.; Papageorgiou, T. Dps-like proteins: Structural and functional insights into a versatile protein family. Cell. Mol. Life Sci. 2009, 67, 341–351. [Google Scholar] [CrossRef]
- Perrin, C.; Guimont, C.; Bracquart, P.; Gaillard, J.-L. Expression of a new cold shock protein of 21.5 kDa and of the major cold shock protein by Streptococcus thermophilus after cold shock. Curr. Microbiol. 1999, 39, 342–347. [Google Scholar] [CrossRef]
- Perrin, C.; Hols, P.; Bracquart, P.; Gaillard, J.-L. Identification of an Iron-Binding Protein of the Dps Family Expressed by Streptococcus thermophilus. Curr. Microbiol. 2004, 48, 51–56. [Google Scholar] [CrossRef]
- Gauss, G.H.; Benas, P.; Wiedenheft, B.; Young, M.; Douglas, T.; Lawrence, C.M. Structure of the DPS-Like Protein from Sulfolobus solfataricus Reveals a Bacterioferritin-Like Dimetal Binding Site within a DPS-Like Dodecameric Assembly†,‡. Biochemistry 2006, 45, 10815–10827. [Google Scholar] [CrossRef] [Green Version]
- Maaty, W.S.; Wiedenheft, B.; Tarlykov, P.; Schaff, N.; Heinemann, J.; Robison-Cox, J.; Valenzuela, J.; Dougherty, A.; Blum, P.; Lawrence, C.M.; et al. Something Old, Something New, Something Borrowed; How the Thermoacidophilic Archaeon Sulfolobus solfataricus Responds to Oxidative Stress. PLoS ONE 2009, 4, e6964. [Google Scholar] [CrossRef] [Green Version]
- Cárdenas, J.; Quatrini, R.; Holmes, D.S. Aerobic Lineage of the Oxidative Stress Response Protein Rubrerythrin Emerged in an Ancient Microaerobic, (Hyper)Thermophilic Environment. Front. Microbiol. 2016, 7, 1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtz, D.M. Avoiding high-valent iron intermediates: Superoxide reductase and rubrerythrin. J. Inorg. Biochem. 2006, 100, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Dillard, B.D.; Demick, J.M.; Adams, M.W.; Lanzilotta, W.N. A cryo-crystallographic time course for peroxide reduction by rubrerythrin from Pyrococcus furiosus. JBIC J. Biol. Inorg. Chem. 2011, 16, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Kameya, M.; Fushinobu, S.; Wakagi, T.; Arai, H.; Ishii, M.; Igarashi, Y. A Novel Enzymatic System against Oxidative Stress in the Thermophilic Hydrogen-Oxidizing Bacterium Hydrogenobacter thermophilus. PLoS ONE 2012, 7, e34825. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Ye, Z.; Zhao, J. RbrA, a cyanobacterial rubrerythrin, functions as a FNR-dependent peroxidase in heterocysts in protection of nitrogenase from damage by hydrogen peroxide in Anabaena sp. PCC 7120. Mol. Microbiol. 2007, 66, 1219–1230. [Google Scholar] [CrossRef]
- Weinberg, M.V.; Jenney, F.E.; Cui, X.; Adams, M.W. Rubrerythrin from the Hyperthermophilic Archaeon Pyrococcus Furiosus Is a Rubredoxin-Dependent, Iron-Containing Peroxidase. J. Bacteriol. 2004, 186, 7888–7895. [Google Scholar] [CrossRef] [Green Version]
- Iyer, R.B.; Silaghi-Dumitrescu, R.; Kurtz, D.M.; Lanzilotta, W.N. High-resolution crystal structures of Desulfovibrio vulgaris (Hildenborough) nigerythrin: Facile, redox-dependent iron movement, domain interface variability, and peroxidase activity in the rubrerythrins. JBIC J. Biol. Inorg. Chem. 2005, 10, 407–416. [Google Scholar] [CrossRef]
- Wakagi, T. Sulerythrin, the smallest member of the rubrerythrin family, from a strictly aerobic and thermoacidophilic archaeon, Sulfolobus tokodaiistrain 7. FEMS Microbiol. Lett. 2003, 222, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Hurtado-Gallego, J.; Martin-Betancor, K.; Rodea-Palomares, I.; Leganés, F.; Rosal, R.; Fernandez-Pinas, F. Two novel cyanobacterial bioluminescent whole-cell bioreporters based on superoxide dismutases MnSod and FeSod to detect superoxide anion. Chemosphere 2018, 201, 772–779. [Google Scholar] [CrossRef] [Green Version]
- Antonucci, I.; Gallo, G.; Limauro, D.; Contursi, P.; Ribeiro, A.L.; Blesa, A.; Berenguer, J.; Bartolucci, S.; Fiorentino, G. Characterization of a promiscuous cadmium and arsenic resistance mechanism in Thermus Thermophilus HB27 and potential application of a novel bioreporter system. Microb. Cell Factories 2018, 17, 78. [Google Scholar] [CrossRef] [Green Version]
- Radyuk, S.N.; Orr, W.C. The Multifaceted Impact of Peroxiredoxins on Aging and Disease. Antioxid. Redox Signal. 2018, 29, 1293–1311. [Google Scholar] [CrossRef] [PubMed]
- Sarcinelli, C.; Fiorentino, G.; Pizzo, E.; Bartolucci, S.; Limauro, D. Discovering Antioxidant Molecules in the Archaea Domain: Peroxiredoxin Bcp1 from Sulfolobus solfataricus Protects H9c2 Cardiomyoblasts from Oxidative Stress. Archaea 2016, 2016, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Laer, K.; Dick, T.P. Utilizing Natural and Engineered Peroxiredoxins As Intracellular Peroxide Reporters. Mol. Cells 2016, 39, 46–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Zhou, Y.; Li, J.; Yu, H.; Takaya, N.; Wang, P.; Zhou, S. Novel peroxiredoxin-based sensor for sensitive detection of hydrogen peroxide. Biochem. Biophys. Res. Commun. 2019, 517, 260–265. [Google Scholar] [CrossRef]
- Lanzilli, M.; Donadio, G.; Fusco, F.A.; Sarcinelli, C.; Limauro, D.; Ricca, E.; Isticato, R. Display of the peroxiredoxin Bcp1 of Sulfolobus solfataricus on probiotic spores of Bacillus megaterium. New Biotechnol. 2018, 46, 38–44. [Google Scholar] [CrossRef]
- Bafana, A.; Dutt, S.; Kumar, S.; Ahuja, P.S. Superoxide dismutase: An industrial perspective. Crit. Rev. Biotechnol. 2010, 31, 65–76. [Google Scholar] [CrossRef]
- Schilrreff, P.; Simioni, Y.R.; Jerez, H.E.; Caimi, A.T.; De Farias, M.A.; Portugal, R.V.; Romero, E.L.; Morilla, M.J. Superoxide dismutase in nanoarchaeosomes for targeted delivery to inflammatory macrophages. Colloids Surf. B Biointerfaces 2019, 179, 479–487. [Google Scholar] [CrossRef]
- Fiorentino, G.; Cannio, R.; Rossi, M.; Bartolucci, S. Decreasing the stability and changing the substrate specificity of the Bacillus Stearothermophilus alcohol dehydrogenase by single amino acid replacements. Protein Eng. 1998, 11, 925–930. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.-H.; Hwang, K.Y.; Choi, J.; Lee, D.Y.; Ahn, B.-Y.; Cho, Y.; Kim, K.-S.; Han, Y.S. Mutational Effects on Thermostable Superoxide Dismutase from Aquifex pyrophilus: Understanding the Molecular Basis of Protein Thermostability. Biochem. Biophys. Res. Commun. 2001, 288, 263–268. [Google Scholar] [CrossRef]
- Li, M.; Zhu, L.; Wang, W. Improving the thermostability and stress tolerance of an archaeon hyperthermophilic superoxide dismutase by fusion with a unique N-terminal domain. SpringerPlus 2016, 5, 241. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Huang, C.; Wang, B.; Guo, H.; Sun, Q.; Xia, F.; Xu, G.; Xia, Q. Enhanced heat tolerance in transgenic silkworm via overexpression of Pyrococcus Furiosus superoxide reductase. Insect Biochem. Mol. Biol. 2018, 92, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, J.; Liu, C.; Luo, J.; Yan, X.; Ai, A.; Cai, Y.; Xie, H.; Ding, X.; Peng, X. Over-expression of a protein disulfide isomerase gene from Methanothermobacter Thermautotrophicus, enhances heat stress tolerance in rice. Gene 2019, 684, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Straub, C.T.; Counts, J.A.; Nguyen, D.M.N.; Wu, C.-H.; Zeldes, B.M.; Crosby, J.R.; Conway, J.M.; Otten, J.K.; Lipscomb, G.L.; Schut, G.J.; et al. Biotechnology of extremely thermophilic archaea. FEMS Microbiol. Rev. 2018, 42, 543–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fe-SOD | Cambialistic SOD |
|---|---|
| Saccharolobus solfataricus | Pyrobaculum aerophilum |
| Sulfolobus acidocaldarius | P. calidifontis |
| Acidianus ambivalens | Aeropyrum pernix |
| Thermoplasma acidophilum |
| Saccharolobus solfataricus | Sulfolobus islandicus | Sulfolobus acidicaldarius | Sulfurisphaera tokodaii |
|---|---|---|---|
| SSO_RS10090 (Bcp1) [69] | SIRE_RS03670 | SACI_RS10765 | STK_RS04035 |
| SSO_RS10350 (Bcp2) [65,66] | SIRE_RS01755(SiPr) [67] | - | STK_RS13435 |
| SSO_RS11005(Bcp3) [65] | SIRE_RS00330 | SACI_RS00260 | STK_RS11650 |
| SSO_RS12680 (Bcp4) [70] | SIRE_RS13090 | SACI_RS05365 | STK_RS09985 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pedone, E.; Fiorentino, G.; Bartolucci, S.; Limauro, D. Enzymatic Antioxidant Signatures in Hyperthermophilic Archaea. Antioxidants 2020, 9, 703. https://doi.org/10.3390/antiox9080703
Pedone E, Fiorentino G, Bartolucci S, Limauro D. Enzymatic Antioxidant Signatures in Hyperthermophilic Archaea. Antioxidants. 2020; 9(8):703. https://doi.org/10.3390/antiox9080703
Chicago/Turabian StylePedone, Emilia, Gabriella Fiorentino, Simonetta Bartolucci, and Danila Limauro. 2020. "Enzymatic Antioxidant Signatures in Hyperthermophilic Archaea" Antioxidants 9, no. 8: 703. https://doi.org/10.3390/antiox9080703
APA StylePedone, E., Fiorentino, G., Bartolucci, S., & Limauro, D. (2020). Enzymatic Antioxidant Signatures in Hyperthermophilic Archaea. Antioxidants, 9(8), 703. https://doi.org/10.3390/antiox9080703