
The Delta and Omicron Variants of SARS-CoV-2: What We Know So Far

 ,
,  and
and
Abstract
:1. Introduction


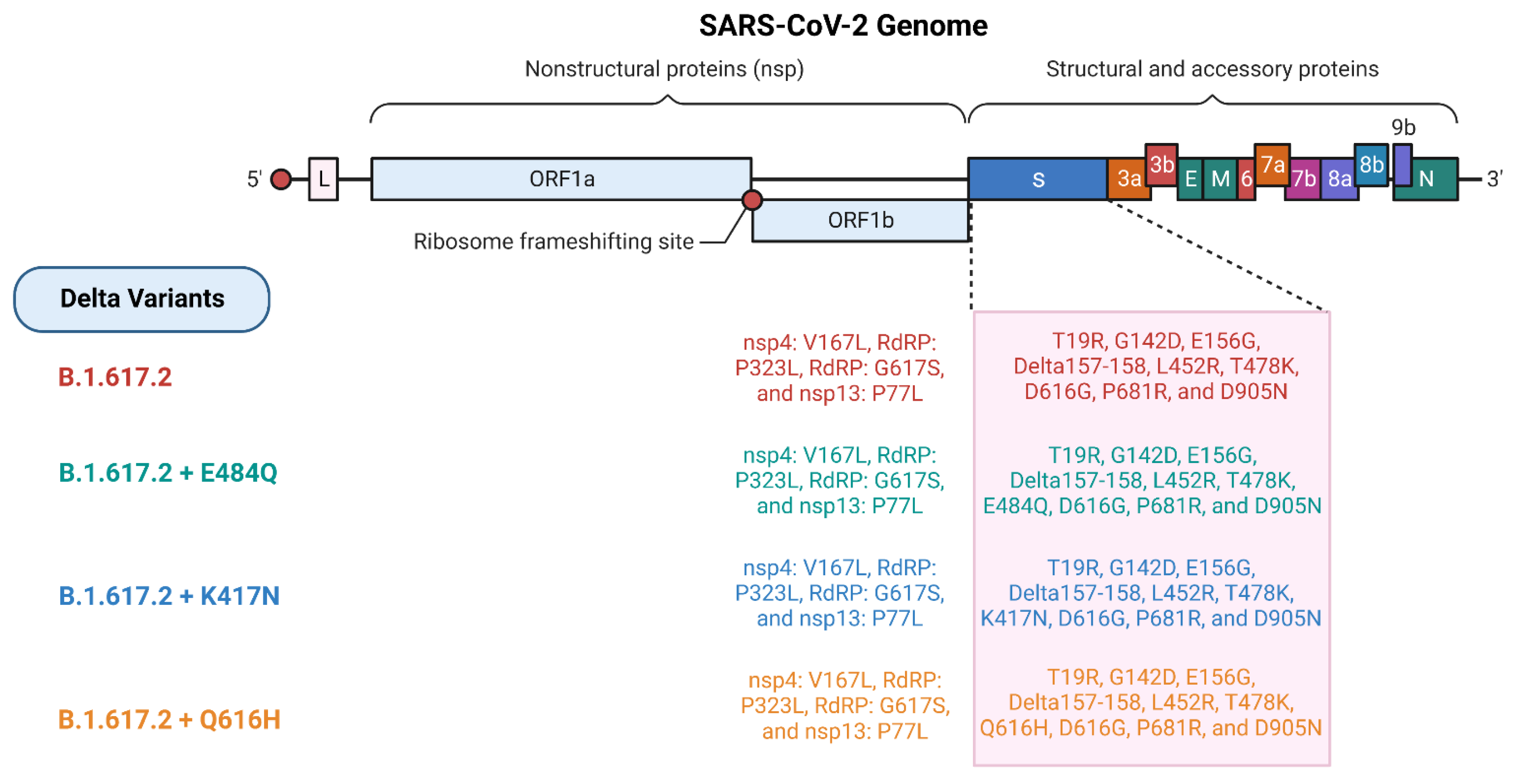
2. Mutation and Pathogenesis of Delta and Omicron Variants

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Amino Acid Mutation | Delta Variant Lineages | Mutation Impact |
|---|---|---|---|
| ORF1a | T3255I | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | The ability of viruses to adapt to internal interactions in host cells, such as replicating and transcribing viral genomes and budding by cellular exocytosis, as well as external interactions with host cells, such as recognizing a cell surface receptor, attaching to the host receptor, and fusing with cellular membranes [66]. |
| ORF1b | P314L | B.1.617.2; B.1.617.2 plus E484Q; B.1.617.2 plus K417N; B.1.617.2 plus Q613H | |
| G662S | B.1.617.2; B.1.617.2 plus E484Q; B.1.617.2 plus K417N; B.1.617.2 plus Q613H | ||
| P1000L | B.1.617.2; B.1.617.2 plus E484Q; B.1.617.2 plus K417N; B.1.617.2 plus Q613H | ||
| S | T19R (NTD) | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | Increase the immune evasion [67]. |
| G142D (NTD) | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | ||
| E156G (NTD) | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | ||
| del157/158 (NTD) | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | ||
| L452R (RBD) | B.1.617.2; B.1.617.2 plus E484Q; B.1.617.2 plus K417N; B.1.617.2 plus Q613H | Impacts neutralization by monoclonal antibodies [68]. | |
| T478K (RBD) | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | Aids in antibody emigration [69]. | |
| D614G (SD2) | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | Enhances the infectivity and density of virion spikes [70]. | |
| P681R (furin-cleavage site) | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | It improved the full-length spike’s cleavage to S1 and S2, which increased infection through cell surface penetration [64,71]. | |
| D950N (S2 region) | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | It might alter the spike protein’s structure to make it better able to shift its form and unite with human cells [72]. | |
| Q613H | B.1.617.2 plus Q613H | Upsurges transmissibility andPathogenicity [73,74]. | |
| K417N | B.1.617.2 plus K417N | Combining the K417N mutation with N501Y eliminated the antibody effect [75]. | |
| E484Q | B.1.617.2 plus E484Q | Exceedingly resistant to neutralization by mAb [76]. | |
| ORF3a | S26L | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | This mutation modifies Orf3a protein dynamics, protein disorder parameters, and secondary structure [77]. |
| M | I82T | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | Crucial for viral assembly, it may also affect glucose transport and decrease type I and type III interferon production, which severely reduces the innate immune response [78]. |
| ORF7a | V82A | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | It restricts host immune suppression with interferon antagonism and might be responsible for causing the expression of pro-inflammatory cytokines [79,80]. |
| T120I | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | ||
| ORF8 | S84L | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | The virus evades the immune system by altering its binding affinity with IRF3 and disrupting chromatin regulation, which speeds up reproduction [81,82]. |
| del119/120 | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | ||
| N | D63G | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | Alteration in secondary structure [83]. |
| R203M | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N | ||
| D377Y | B.1.617.2; B.1.617.2 plus Q613H; B.1.617.2 plus E484Q; B.1.617.2 plus K417N |
3. Omicron Variant vs. Delta Variant
4. Vaccine Efficacy
4.1. Delta Variants Influence Vaccine Efficacy
4.2. Omicron Variants Influence Vaccine Efficacy
5. Therapeutics for the Delta and Omicron Variants
6. Delta Omicron Recombinant Variant
7. Concluding Remarks and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, Y.C.; Chen, C.S.; Chan, Y.J. The Outbreak of COVID-19: An Overview. J. Chin. Med. Assoc. 2020, 83, 217. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Hasöksüz, M.; Kiliç, S.; Saraç, F. Coronaviruses and SARS-COV-2. Turk. J. Med. Sci. 2020, 50, 549–556. [Google Scholar] [CrossRef]
- Roy, B.; Dhillon, J.; Habib, N.; Pugazhandhi, B. Global Variants of COVID-19: Current Understanding. J. Biomed. Sci. 2021, 8, 8–11. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Naming the Coronavirus Disease (COVID-19) and the Virus That Causes It; World Health Organization: Geneva, Switzerland, 2020; Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/technical-guidance/naming-the-coronavirus-disease-(covid-2019)-and-the-virus-that-causes-it (accessed on 13 September 2022).
- World Health Organization (WHO). WHO Director-General’s Opening Remarks at the Media Briefing on COVID-19; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Belsky, J.A.; Tullius, B.P.; Lamb, M.G.; Sayegh, R.; Stanek, J.R.; Auletta, J.J. COVID-19 in Immunocompromised Patients: A Systematic Review of Cancer, Hematopoietic Cell and Solid Organ Transplant Patients. J. Infect. 2021, 82, 329. [Google Scholar] [CrossRef]
- Baek, M.S.; Lee, M.T.; Kim, W.Y.; Choi, J.C.; Jung, S.Y. COVID-19-Related Outcomes in Immunocompromised Patients: A Nationwide Study in Korea. PLoS ONE 2021, 16, e0257641. [Google Scholar] [CrossRef]
- Davidson, A.M.; Wysocki, J.; Batlle, D. Interaction of SARS-CoV-2 and Other Coronavirus with ACE (Angiotensin-Converting Enzyme)-2 as Their Main Receptor. Hypertension 2020, 76, 1339–1349. [Google Scholar] [CrossRef]
- Chavda, V.P.; Pandya, R.; Apostolopoulos, V. DNA Vaccines for SARS-CoV-2: Toward Third-Generation Vaccination Era. Expert Rev. Vaccines 2021, 20, 1549–1560. [Google Scholar] [CrossRef]
- Chavda, V.P.; Vora, L.K.; Vihol, D.R. COVAX-19® Vaccine: Completely Blocks Virus Transmission to Non-Immune Individuals. Clin. Complement. Med. Pharmacol. 2021, 1, 100004. [Google Scholar] [CrossRef]
- Chavda, V.P.; Vora, L.K.; Pandya, A.K.; Patravale, V.B. Intranasal Vaccines for SARS-CoV-2: From Challenges to Potential in COVID-19 Management. Drug Discov. Today 2021, 26, 2619–2636. [Google Scholar] [CrossRef]
- Chavda, V.P.; Apostolopoulos, V. Mucormycosis—An Opportunistic Infection in the Aged Immunocompromised Individual: A Reason for Concern in COVID-19. Maturitas 2021, 58, 58–61. [Google Scholar] [CrossRef]
- Chavda, V.P.; Gajjar, N.; Shah, N.; Dave, D.J. Darunavir Ethanolate: Repurposing an Anti-HIV Drug in COVID-19 Treatment. Eur. J. Med. Chem. Rep. 2021, 3, 100013. [Google Scholar] [CrossRef]
- Chavda, V.P.; Feehan, J.; Apostolopoulos, V. A Veterinary Vaccine for SARS-CoV-2: The First COVID-19 Vaccine for Animals. Vaccines 2021, 9, 631. [Google Scholar] [CrossRef]
- Karim, S.S.A.; Karim, Q.A. Omicron SARS-CoV-2 Variant: A New Chapter in the COVID-19 Pandemic. Lancet 2021, 398, 2126–2128. [Google Scholar] [CrossRef]
- Jungreis, I.; Sealfon, R.; Kellis, M. SARS-CoV-2 Gene Content and COVID-19 Mutation Impact by Comparing 44 Sarbecovirus Genomes. Nat. Commun. 2021, 12, 2642. [Google Scholar] [CrossRef]
- Chavda, V.P.; Patel, A.B.; Vaghasiya, D.D. SARS-CoV-2 Variants and Vulnerability at the Global Level. J. Med. Virol. 2022, 94, 2986–3005. [Google Scholar] [CrossRef]
- Corey, L.; Beyrer, C.; Cohen, M.S.; Michael, N.L.; Bedford, T.; Rolland, M. SARS-CoV-2 Variants in Patients with Immunosuppression. N. Engl. J. Med. 2021, 385, 562–566. [Google Scholar] [CrossRef]
- Abbasi, J. Omicron Has Reached the US—Here’s What Infectious Disease Experts Know About the Variant. JAMA 2021, 326, 2460–2462. [Google Scholar] [CrossRef]
- (INSACOG), T.I.S.-C.-2 G.C. Genome Sequencing by INSACOG Shows Variants of Concern and a Novel Variant in India. Available online: https://pib.gov.in/PressReleaseIframePage.aspx?PRID=1707177 (accessed on 19 December 2021).
- Vashishtha, V. Are the New Variants Driving India’s Second COVID-19 Wave? Available online: https://science.thewire.in/the-sciences/are-the-new-variants-driving-indias-second-covid-19-wave/ (accessed on 20 May 2021).
- Chavda, V.P.; Kapadia, C.; Soni, S.; Prajapati, R.; Chauhan, S.C.; Yallapu, M.M.; Apostolopoulos, V. A Global Picture: Therapeutic Perspectives for COVID-19. Immunotherapy 2022, 14, 351–371. [Google Scholar] [CrossRef]
- Ferreira, I.; Datir, R.; Papa, G.; Kemp, S.; Meng, B.; Rakshit, P.; Singh, S.; Pandey, R.; Ponnusamy, K.; Radhakrishnan, V.S.; et al. SARS-CoV-2 B.1.617 Emergence and Sensitivity to Vaccine-Elicited Antibodies. BioRxiv 2021. [Google Scholar] [CrossRef]
- Kai, K.; Meredith, W. Delta Variant Trigfgers New Phase in the Pandemic. Science 2021, 372, 1375–1376. [Google Scholar] [CrossRef]
- Kumar, S.; Thambiraja, T.S.; Karuppanan, K.; Subramaniam, G. Omicron and Delta Variant of SARS-CoV-2: A Comparative Computational Study of Spike Protein. J. Med. Virol. 2022, 94, 1641–1649. [Google Scholar] [CrossRef]
- Chavda, V.P.; Apostolopoulos, V. Global Impact of Delta plus Variant and Vaccination. Expert Rev. Vaccines 2022, 21, 597–600. [Google Scholar] [CrossRef]
- Elflein, J. Number of SARS-CoV-2 Delta Variant Cases Worldwide. Available online: https://www.statista.com/statistics/1245971/number-delta-variant-worldwide-by-country/ (accessed on 15 December 2021).
- Susmita Pakrasi Delta Variant Expected to Be Dominant COVID-19 Strain Globally, Says WHO Chief. Available online: https://www.hindustantimes.com/world-news/delta-variant-expected-to-be-dominant-covid-19-strain-globally-says-who-chief-101626147042896.html (accessed on 15 July 2021).
- Shiehzadegan, S.; Alaghemand, N.; Fox, M.; Venketaraman, V. Analysis of the Delta Variant, B.1.617.2 COVID-19. Clin. Pract. 2021, 11, 778–784. [Google Scholar] [CrossRef]
- Ranjan, R.; Sharma, A.; Verma, M.K. Characterization of the Second Wave of COVID-19 in India. Curr. Sci. 2021, 121. [Google Scholar] [CrossRef]
- Cherian, S.; Potdar, V.; Jadhav, S.; Yadav, P.; Gupta, N.; Das, M.; Rakshit, P.; Singh, S.; Abraham, P.; Panda, S.; et al. Convergent Evolution of SARS-CoV-2 Spike Mutations, L452R, E484Q and P681R, in the Second Wave of COVID-19 in Maharashtra, India. Microorganisms 2021, 9, 1542. [Google Scholar] [CrossRef]
- Edara, V.V.; Lai, L.; Sahoo, M.; Floyd, K.; Sibai, M.; Solis, D.; Flowers, M.W.; Hussaini, L.; Ciric, C.R.; Bechnack, S.; et al. Infection and Vaccine-Induced Neutralizing Antibody Responses to the SARS-CoV-2 B. 1.617. 1 Variant. BioRxiv 2021. Available online: https://www.Biorxiv.or (accessed on 17 September 2022). [CrossRef]
- Hagen, A. How Dangerous Is the Delta Variant (B.1.617.2)? Available online: https://asm.org/Articles/2021/July/How-Dangerous-is-the-Delta-Variant-B-1-617-2 (accessed on 15 December 2021).
- Fisman, D.N.; Tuite, A.R. Evaluation of the Relative Virulence of Novel SARS-CoV-2 Variants: A Retrospective Cohort Study in Ontario, Canada. Can. Med. Assoc. J. 2021, 193, E1619–E1625. [Google Scholar] [CrossRef]
- He, X.; He, C.; Hong, W.; Zhang, K.; Wei, X. The Challenges of COVID-19 Delta Variant: Prevention and Vaccine Development. MedComm 2021, 2, 846–854. [Google Scholar] [CrossRef]
- WHO. Classification of Omicron (B.1.1.529): SARS-CoV-2 Variant of Concern. Available online: https://www.who.int/news/item/26-11-2021-classification-of-omicron-(b.1.1.529)-sars-cov-2-variant-of-concern (accessed on 30 November 2021).
- Shrestha, L.B.; Foster, C.; Rawlinson, W.; Tedla, N.; Bull, R.A. Evolution of the SARS-CoV-2 Omicron Variants BA.1 to BA.5: Implications for Immune Escape and Transmission. Rev. Med. Virol. 2022, 32, e2381. [Google Scholar] [CrossRef]
- WHO. Director-General’s Opening Remarks at Media Briefing; WHO: Geneva, Switzerland, 2022. [Google Scholar]
- Tian, D.; Sun, Y.; Xu, H.; Ye, Q. The Emergence and Epidemic Characteristics of the Highly Mutated SARS-CoV-2 Omicron Variant. J. Med. Virol. 2022, 94, 2376–2383. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; COVID-19 Genomics UK (COG-UK) Consortium; et al. SARS-CoV-2 Variants, Spike Mutations and Immune Escape. Nat. Rev. Microbiol. 2021, 614, 409–424. [Google Scholar] [CrossRef]
- Chi, X.; Yan, R.; Zhang, J.; Zhang, G.; Zhang, Y.; Hao, M.; Zhang, Z.; Fan, P.; Dong, Y.; Yang, Y.; et al. A Neutralizing Human Antibody Binds to the N-Terminal Domain of the Spike Protein of SARS-CoV-2. Science 2020, 369, 650–655. [Google Scholar] [CrossRef]
- Tian, D.; Sun, Y.H.; Zhou, J.M.; Ye, Q. The Global Epidemic of SARS-CoV-2 Variants and Their Mutational Immune Escape. J. Med. Virol. 2022, 94, 847–857. [Google Scholar] [CrossRef]
- Tian, D.; Sun, Y.; Zhou, J.; Ye, Q. The Global Epidemic of the SARS-CoV-2 Delta Variant, Key Spike Mutations and Immune Escape. Front. Immunol. 2021, 12, 751778. [Google Scholar] [CrossRef]
- Ranjan, R. Omicron Impact in India: Analysis of the Ongoing COVID-19 Third Wave Based on Global Data. medRxiv 2022, 1–17. [Google Scholar] [CrossRef]
- UNICEF. What We Know about the Omicron Variant; UNICEF: New York, NY, USA, 2022. [Google Scholar]
- Chavda, V.P.; Apostolopoulos, V. Omicron Variant (B.1.1.529) of SARS-CoV-2: Threat for the Elderly? Maturitas 2022, 158, 78–81. [Google Scholar] [CrossRef]
- Sheward, D.J.; Kim, C.; Fischbach, J.; Muschiol, S.; Ehling, R.A.; Björkström, N.K.; Hedestam, G.B.K.; Reddy, S.T.; Albert, J.; Peacock, T.P.; et al. Evasion of Neutralising Antibodies by Omicron Sublineage BA.2.75. Lancet Infect. Dis. 2022, 22, 1421–1422. [Google Scholar] [CrossRef]
- Chavda, V.P.; Bezbaruah, R.; Athalye, M.; Parikh, P.K.; Chhipa, A.S.; Patel, S.; Apostolopoulos, V. Replicating Viral Vector-Based Vaccines for COVID-19: Potential Avenue in Vaccination Arena. Viruses 2022, 14, 759. [Google Scholar] [CrossRef]
- Aydogdu, M.O.; Rohn, J.L.; Jafari, N.V.; Brako, F.; Homer-Vanniasinkam, S.; Edirisinghe, M. Severe Acute Respiratory Syndrome Type 2-Causing Coronavirus: Variants and Preventive Strategies. Adv. Sci. 2022, 9, 2104495. [Google Scholar] [CrossRef]
- Abdool Karim, S.S.; de Oliveira, T. New SARS-CoV-2 Variants—Clinical, Public Health, and Vaccine Implications. N. Engl. J. Med. 2021, 384, 1866–1868. [Google Scholar] [CrossRef]
- Gupta, R.K.; Nwachuku, E.L.; Zusman, B.E.; Jha, R.M.; Puccio, A.M. Drug Repurposing for COVID-19 Based on an Integrative Meta-Analysis of SARS-CoV-2 Induced Gene Signature in Human Airway Epithelium. PLoS ONE 2021, 16, e0257784. [Google Scholar] [CrossRef]
- Chavda, V.P.; Hossain, M.K.; Beladiya, J.; Apostolopoulos, V. Nucleic Acid Vaccines for COVID-19: A Paradigm Shift in the Vaccine Development Arena. Biologics 2021, 1, 337–356. [Google Scholar] [CrossRef]
- Dong, Y.; Dai, T.; Wei, Y.; Zhang, L. A Systematic Review of SARS-CoV-2 Vaccine Candidates. Signal Transduct. Target. Ther. 2020, 5, 2–14. [Google Scholar] [CrossRef]
- Richman, D.D. COVID-19 Vaccines: Implementation, Limitations and Opportunities. Glob. Health Med. 2021, 3, 1–5. [Google Scholar] [CrossRef]
- Chavda, V.P.; Apostolopoulos, V. Is Booster Dose Strategy Sufficient for Omicron Variant of SARS-CoV-2? Vaccines 2022, 10, 367. [Google Scholar] [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 Spike Receptor-Binding Domain Bound to the ACE2 Receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Shah, V.K.; Firmal, P.; Alam, A.; Ganguly, D.; Chattopadhyay, S. Overview of Immune Response During SARS-CoV-2 Infection: Lessons From the Past. Front. Immunol. 2020, 11, 1949. [Google Scholar] [CrossRef]
- Lubinski, B.; Frazier, L.E.; Phan, M.V.T.; Bugembe, D.L.; Tang, T.; Daniel, S.; Cotten, M.; Jaimes, J.A.; Whittaker, G.R. Spike Protein Cleavage-Activation Mediated by the SARS-CoV-2 P681R Mutation: A Case-Study from Its First Appearance in Variant of Interest (VOI) A.23.1 Identified in Uganda. bioRxiv 2021. [Google Scholar] [CrossRef]
- Johnson, B.A.; Xie, X.; Bailey, A.L.; Kalveram, B.; Lokugamage, K.G.; Muruato, A.; Zou, J.; Zhang, X.; Juelich, T.; Smith, J.K.; et al. Loss of Furin Cleavage Site Attenuates SARS-CoV-2 Pathogenesis. Nature 2021, 591, 293–299. [Google Scholar] [CrossRef]
- Bian, L.; Gao, Q.; Gao, F.; Wang, Q.; He, Q.; Wu, X.; Mao, Q.; Xu, M.; Liang, Z. Impact of the Delta Variant on Vaccine Efficacy and Response Strategies. Expert Rev. Vaccines 2021, 20, 1201–1209. [Google Scholar] [CrossRef]
- Yaqinuddin, A.; Shafqat, A.; Kashir, J.; Alkattan, K. Effect of SARS-CoV-2 Mutations on the Efficacy of Antibody Therapy and Response to Vaccines. Vaccines 2021, 9, 914. [Google Scholar] [CrossRef]
- Noori, M.; Nejadghaderi, S.A.; Arshi, S.; Carson-Chahhoud, K.; Ansarin, K.; Kolahi, A.-A.; Safiri, S. Potency of BNT162b2 and MRNA-1273 Vaccine-Induced Neutralizing Antibodies against Severe Acute Respiratory Syndrome-CoV-2 Variants of Concern: A Systematic Review of in Vitro Studies. Rev. Med. Virol. 2021, 32, e2277. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.; Johnson, B.A.; Xia, H.; Ku, Z.; Schindewolf, C.; Widen, S.G.; An, Z.; Weaver, S.C.; Menachery, V.D.; et al. Delta Spike P681R Mutation Enhances SARS-CoV-2 Fitness over Alpha Variant. Cell Rep. 2022, 39, 110829. [Google Scholar] [CrossRef]
- Latif, A.A.; Mullen, J.L.; Alkuzweny, M.; Tsueng, G.; Cano, M.; Haag, E.; Zhou, J.; Zeller, M.; Hufbauer, E.; Matteson, N.; et al. The Center for Viral Systems Biology, Lineage comparison. outbreak.info. Available online: https://outbreak.info/compare-lineages (accessed on 9 September 2022).
- Kim, J.-S.; Jang, J.-H.; Kim, J.-M.; Chung, Y.-S.; Yoo, C.-K.; Han, M.-G. Genome-Wide Identification and Characterization of Point Mutations in the SARS-CoV-2 Genome. Osong Public HealTH Res. Perspect. 2020, 11, 101–111. [Google Scholar] [CrossRef]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Puech, J.; et al. Reduced Sensitivity of SARS-CoV-2 Variant Delta to Antibody Neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef]
- Starr, T.N.; Greaney, A.J.; Dingens, A.S.; Bloom, J.D. Complete Map of SARS-CoV-2 RBD Mutations That Escape the Monoclonal Antibody LY-CoV555 and Its Cocktail with LY-CoV016. Cell Rep. Med. 2021, 2, 100255. [Google Scholar] [CrossRef]
- Plante, J.A.; Mitchell, B.M.; Plante, K.S.; Debbink, K.; Weaver, S.C.; Menachery, V.D. The Variant Gambit: COVID-19’s next Move. Cell Host Microbe 2021, 29, 508–515. [Google Scholar] [CrossRef]
- Zhang, L.; Jackson, C.B.; Mou, H.; Ojha, A.; Peng, H.; Quinlan, B.D.; Rangarajan, E.S.; Pan, A.; Vanderheiden, A.; Suthar, M.S.; et al. SARS-CoV-2 Spike-Protein D614G Mutation Increases Virion Spike Density and Infectivity. Nat. Commun. 2020, 11, 6013. [Google Scholar] [CrossRef]
- Saito, A.; Irie, T.; Suzuki, R.; Maemura, T.; Nasser, H.; Uriu, K.; Kosugi, Y.; Shirakawa, K.; Sadamasu, K.; Kimura, I.; et al. Enhanced Fusogenicity and Pathogenicity of SARS-CoV-2 Delta P681R Mutation. Nature 2022, 602, 300–306. [Google Scholar] [CrossRef]
- Bai, C.; Wang, J.; Chen, G.; Zhang, H.; An, K.; Xu, P.; Du, Y.; Ye, R.D.; Saha, A.; Zhang, A.; et al. Predicting Mutational Effects on Receptor Binding of the Spike Protein of SARS-CoV-2 Variants. J. Am. Chem. Soc. 2021, 143, 17646–17654. [Google Scholar] [CrossRef]
- Guruprasad, L. Human SARS CoV-2 Spike Protein Mutations. Proteins Struct. Funct. Bioinforma. 2021, 89, 569–576. [Google Scholar] [CrossRef]
- Volz, E.; Hill, V.; McCrone, J.T.; Price, A.; Jorgensen, D.; O’Toole, Á.; Southgate, J.; Johnson, R.; Jackson, B.; Nascimento, F.F.; et al. Evaluating the Effects of SARS-CoV-2 Spike Mutation D614G on Transmissibility and Pathogenicity. Cell 2021, 184, 64.e11–75.e11. [Google Scholar] [CrossRef]
- Fratev, F. N501Y and K417N Mutations in the Spike Protein of SARS-CoV-2 Alter the Interactions with Both HACE2 and Human-Derived Antibody: A Free Energy of Perturbation Retrospective Study. J. Chem. Inf. Model. 2021, 61, 6079–6084. [Google Scholar] [CrossRef]
- Ding, C.; He, J.; Zhang, X.; Jiang, C.; Sun, Y.; Zhang, Y.; Chen, Q.; He, H.; Li, W.; Xie, J.; et al. Crucial Mutations of Spike Protein on SARS-CoV-2 Evolved to Variant Strains Escaping Neutralization of Convalescent Plasmas and RBD-Specific Monoclonal Antibodies. Front. Immunol. 2021, 12, 3231. [Google Scholar] [CrossRef]
- Azad, G.K.; Khan, P.K. Variations in Orf3a Protein of SARS-CoV-2 Alter Its Structure and Function. Biochem. Biophys. Rep. 2021, 26, 100933. [Google Scholar] [CrossRef]
- Shen, L.; Bard, J.D.; Triche, T.J.; Judkins, A.R.; Biegel, J.A.; Gai, X. Emerging Variants of Concern in SARS-CoV-2 Membrane Protein: A Highly Conserved Target with Potential Pathological and Therapeutic Implications. Emerg. Microbes Infect. 2021, 10, 885–893. [Google Scholar] [CrossRef]
- Kannan, S.R.; Spratt, A.N.; Cohen, A.R.; Naqvi, S.H.; Chand, H.S.; Quinn, T.P.; Lorson, C.L.; Byrareddy, S.N.; Singh, K. Evolutionary Analysis of the Delta and Delta Plus Variants of the SARS-CoV-2 Viruses. J. Autoimmun. 2021, 124, 102715. [Google Scholar] [CrossRef]
- Nemudryi, A.; Nemudraia, A.; Wiegand, T.; Nichols, J.; Snyder, D.T.; Hedges, J.F.; Cicha, C.; Lee, H.; Vanderwood, K.K.; Bimczok, D.; et al. SARS-CoV-2 Genomic Surveillance Identifies Naturally Occurring Truncations of ORF7a That Limit Immune Suppression. Cell Rep. 2021, 35, 109197. [Google Scholar] [CrossRef]
- Kee, J.; Thudium, S.; Renner, D.; Glastad, K.; Palozola, K.; Zhang, Z.; Li, Y.; Cesare, J.; Lan, Y.; Truitt, R.; et al. SARS-CoV-2 ORF8 Encoded Protein Contains a Histone Mimic, Disrupts Chromatin Regulation, and Enhances Replication. bioRxiv 2021. [Google Scholar] [CrossRef]
- Rashid, F.; Suleman, M.; Shah, A.; Dzakah, E.E.; Wang, H.; Chen, S.; Tang, S. Mutations in SARS-CoV-2 ORF8 Altered the Bonding Network with Interferon Regulatory Factor 3 to Evade Host Immune System. Front. Microbiol. 2021, 12, 1811. [Google Scholar] [CrossRef] [PubMed]
- Azad, G.K. The Molecular Assessment of SARS-CoV-2 Nucleocapsid Phosphoprotein Variants among Indian Isolates. Heliyon 2021, 7, e06167. [Google Scholar] [CrossRef]
- Butowt, R.; Bilińska, K.; von Bartheld, C. Why Does the Omicron Variant Largely Spare Olfactory Function? Implications for the Pathogenesis of Anosmia in Coronavirus Disease 2019. J. Infect. Dis. 2022, 226, 1304–1308. [Google Scholar] [CrossRef]
- Khan, N.A.; Al-Thani, H.; El-Menyar, A. The Emergence of New SARS-CoV-2 Variant (Omicron) and Increasing Calls for COVID-19 Vaccine Boosters-The Debate Continues. Travel Med. Infect. Dis. 2022, 45, 102246. [Google Scholar] [CrossRef]
- He, X.; Hong, W.; Pan, X.; Lu, G.; Wei, X. SARS-CoV-2 Omicron Variant: Characteristics and Prevention. MedComm 2021, 2, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Li, X.; Zhang, L.; Wan, S.; Zhang, L.; Zhou, F. SARS-CoV-2 Omicron Variant: Recent Progress and Future Perspectives. Signal Transduct. Target. Ther. 2022, 7, 141. [Google Scholar] [CrossRef]
- Hsieh, C.L.; Goldsmith, J.A.; Schaub, J.M.; DiVenere, A.M.; Kuo, H.C.; Javanmardi, K.; Le, K.C.; Wrapp, D.; Lee, A.G.; Liu, Y.; et al. Structure-Based Design of Prefusion-Stabilized SARS-CoV-2 Spikes. Science 2020, 369, 1501–1505. [Google Scholar] [CrossRef]
- Cao, Y.; Yisimayi, A.; Jian, F.; Song, W.; Xiao, T.; Wang, L.; Du, S.; Wang, J.; Li, Q.; Chen, X.; et al. BA.2.12.1, BA.4 and BA.5 Escape Antibodies Elicited by Omicron Infection. Nature 2022, 608, 593–602. [Google Scholar] [CrossRef]
- CoVariants: 21L (Omicron). Available online: https://covariants.org/variants/21L.Omicron (accessed on 9 September 2022).
- Wise, J. COVID-19: The E484K Mutation and the Risks It Poses. BMJ 2021, 372, n359. [Google Scholar] [CrossRef]
- Gong, S.Y.; Chatterjee, D.; Richard, J.; Prévost, J.; Tauzin, A.; Gasser, R.; Bo, Y.; Vézina, D.; Goyette, G.; Gendron-Lepage, G.; et al. Contribution of Single Mutations to Selected SARS-CoV-2 Emerging Variants Spike Antigenicity. Virology 2021, 563, 134–145. [Google Scholar] [CrossRef]
- Zahradník, J.; Marciano, S.; Shemesh, M.; Zoler, E.; Harari, D.; Chiaravalli, J.; Meyer, B.; Rudich, Y.; Li, C.; Marton, I.; et al. SARS-CoV-2 Variant Prediction and Antiviral Drug Design Are Enabled by RBD in Vitro Evolution. Nat. Microbiol. 2021, 6, 1188–1198. [Google Scholar] [CrossRef] [PubMed]
- Leary, S.; Gaudieri, S.; Parker, M.D.; Chopra, A.; James, I.; Pakala, S.; Alves, E.; John, M.; Lindsey, B.B.; Keeley, A.J.; et al. Generation of a Novel SARS-CoV-2 Sub-Genomic RNA Due to the R203K/G204R Variant in Nucleocapsid. bioRxiv 2021. [Google Scholar] [CrossRef] [Green Version]
- Mourier, T.; Shuaib, M.; Hala, S.; Mfarrej, S.; Alofi, F.; Naeem, R.; Alsomali, A.; Jorgensen, D.; Subudhi, A.K.; Rached, F.B.; et al. Saudi Arabian SARS-CoV-2 Genomes Implicate a Mutant Nucleocapsid Protein in Modulating Host Interactions and Increased Viral Load in COVID-19 Patients. Nat. Commun. 2021, 13, 601. [Google Scholar] [CrossRef]
- Benvenuto, D.; Angeletti, S.; Giovanetti, M.; Bianchi, M.; Pascarella, S.; Cauda, R.; Ciccozzi, M.; Cassone, A. Evolutionary Analysis of SARS-CoV-2: How Mutation of Non-Structural Protein 6 (NSP6) Could Affect Viral Autophagy. J. Infect. 2020, 81, e24–e27. [Google Scholar] [CrossRef]
- Beyer, D.K.; Forero, A. Mechanisms of Antiviral Immune Evasion of SARS-CoV-2. J. Mol. Biol. 2022, 434, 167265. [Google Scholar] [CrossRef] [PubMed]
- Thorne, L.G.; Bouhaddou, M.; Reuschl, A.-K.; Zuliani-Alvarez, L.; Polacco, B.; Pelin, A.; Batra, J.; Whelan, M.V.X.; Ummadi, M.; Rojc, A.; et al. Evolution of Enhanced Innate Immune Evasion by the SARS-CoV-2 B.1.1.7 UK Variant. bioRxiv 2021. [Google Scholar] [CrossRef]
- Collins, F. Latest on Omicron Variant and COVID-19 Vaccine Protection. Available online: https://directorsblog.nih.gov/2021/12/14/the-latest-on-the-omicron-variant-and-vaccine-protection/ (accessed on 16 December 2021).
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Rickeard, T.; Gallagher, E.; Gower, C.; Kall, M.; Groves, N.; O’Connell, A.-M.; et al. Effectiveness of COVID-19 Vaccines against the Omicron (B.1.1.529) Variant of Concern. N. Engl. J. Med. 2022, 386, 1532–1546. [Google Scholar] [CrossRef]
- Cele, S.; Jackson, L.; Khan, K.; Khoury, D.; Moyo-Gwete, T.; Tegally, H.; Scheepers, C.; Amoako, D.; Karim, F.; Bernstein, M.; et al. SARS-CoV-2 Omicron Has Extensive but Incomplete Escape of Pfizer BNT162b2 Elicited Neutralization and Requires ACE2 for Infection. medRxiv 2021. [Google Scholar] [CrossRef]
- Singanayagam, A.; Hakki, S.; Dunning, J.; Madon, K.J.; Crone, M.A.; Koycheva, A.; Derqui-Fernandez, N.; Barnett, J.L.; Whitfield, M.G.; Varro, R.; et al. Community Transmission and Viral Load Kinetics of the SARS-CoV-2 Delta (B.1.617.2) Variant in Vaccinated and Unvaccinated Individuals in the UK: A Prospective, Longitudinal, Cohort Study. Lancet Infect. Dis. 2021, 22, 183–195. [Google Scholar] [CrossRef]
- Elflein, J. Number of SARS-CoV-2 Omicron Variant Cases Worldwide as of 16 December 2021, by Country or Territory. Available online: https://www.statista.com/statistics/1279100/number-omicron-variant-worldwide-by-country/ (accessed on 16 December 2021).
- Dejnirattisai, W.; Shaw, R.H.; Supasa, P.; Liu, C.; Stuart, A.S.V.; Pollard, A.J.; Liu, X.; Lambe, T.; Crook, D.; Stuart, D.I.; et al. Reduced Neutralisation of SARS-COV-2 Omicron-B.1.1.529 Variant by Post-Immunisation Serum. Lancet 2022, 399, 234–236. [Google Scholar] [CrossRef]
- Chavda, V.P.; Vuppu, S.; Mishra, T.; Kamaraj, S.; Patel, A.B.; Sharma, N.; Chen, Z.-S. Recent Review of COVID-19 Management: Diagnosis, Treatment and Vaccination. Pharmacol. Rep. 2022, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Li, J.; Zhang, L.; Hu, W.; Zong, L.; Xu, X.; Qiao, J.; Zheng, M.; Jiang, X.; Liang, Z.; et al. Identification of SARS-CoV-2 Variants and Their Clinical Significance in Hefei, China. Front. Med. 2022, 8, 784632. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.P.; Yao, Q.; Vora, L.K.; Apostolopoulos, V.; Patel, C.A.; Bezbaruah, R.; Patel, A.B.; Chen, Z.-S. Fast-Track Development of Vaccines for SARS-CoV-2: The Shots That Saved the World. Front. Immunol. 2022, 13, 961198. [Google Scholar] [CrossRef]
- Chitsike, L.; Duerksen-Hughes, P. Keep out! SARS-CoV-2 Entry Inhibitors: Their Role and Utility as COVID-19 Therapeutics. Virol. J. 2021, 18, 154. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.P.; Prajapati, R.; Lathigara, D.; Nagar, B.; Kukadiya, J.; Redwan, E.M.; Uversky, V.N.; Kher, M.N.; Patel, R. Therapeutic Monoclonal Antibodies for COVID-19 Management: An Update. Expert Opin. Biol. Ther. 2022, 22, 763–780. [Google Scholar] [CrossRef]
- Chavda, V.P.; Apostolopoulos, V. COVID-19 Vaccine Design and Vaccination Strategy for Emerging Variants. Expert Rev. Vaccines 2022, 21, 1359–1361. [Google Scholar] [CrossRef]
- Campbell, F.; Archer, B.; Laurenson-Schafer, H.; Jinnai, Y.; Konings, F.; Batra, N.; Pavlin, B.; Vandemaele, K.; Van Kerkhove, M.D.; Jombart, T.; et al. Increased Transmissibility and Global Spread of SARS-CoV-2 Variants of Concern as at June 2021. Euro Surveill. Bull. Eur. sur les Mal. Transm. = Eur. Commun. Dis. Bull. 2021, 26, 2100509. [Google Scholar] [CrossRef]
- Hacisuleyman, E.; Hale, C.; Saito, Y.; Blachere, N.E.; Bergh, M.; Conlon, E.G.; Schaefer-Babajew, D.J.; DaSilva, J.; Muecksch, F.; Gaebler, C.; et al. Vaccine Breakthrough Infections with SARS-CoV-2 Variants. N. Engl. J. Med. 2021, 384, 2212–2218. [Google Scholar] [CrossRef]
- Ahmad, L. Implication of SARS-CoV-2 Immune Escape Spike Variants on Secondary and Vaccine Breakthrough Infections. Front. Immunol. 2021, 12, 742167. [Google Scholar] [CrossRef]
- Dingemans, J.; van der Veer, B.M.J.W.; Gorgels, K.M.F.; Hackert, V.; Hensels, A.Y.J.; den Heijer, C.D.J.; Hoebe, C.J.P.A.; Savelkoul, P.H.M.; van Alphen, L.B. Investigating SARS-CoV-2 Breakthrough Infections per Variant and Vaccine Type. medRxiv 2021. [Google Scholar] [CrossRef]
- Chen, C.-J.; Yang, L.-Y.; Chang, W.-Y.; Huang, Y.-C.; Chiu, C.-H.; Shih, S.-R.; Huang, C.-G.; Huang, K.-Y.A. A Randomized Controlled Trial of Heterologous ChAdOx1 NCoV-19 and Recombinant Subunit Vaccine MVC-COV1901 against COVID-19. Nat. Commun. 2022, 13, 5466. [Google Scholar] [CrossRef] [PubMed]
- Groß, R.; Zanoni, M.; Seidel, A.; Conzelmann, C.; Gilg, A.; Krnavek, D.; Erdemci-Evin, S.; Mayer, B.; Hoffmann, M.; Pöhlmann, S.; et al. Heterologous ChAdOx1 NCoV-19 and BNT162b2 Prime-Boost Vaccination Elicits Potent Neutralizing Antibody Responses and T Cell Reactivity. eBioMedicine 2022, 75, 103761. [Google Scholar] [CrossRef] [PubMed]
- Sapkal, G.N.; Yadav, P.D.; Sahay, R.R.; Deshpande, G.; Gupta, N.; Nyayanit, D.A.; Patil, D.Y.; Kumar, S.; Abraham, P.; Panda, S.; et al. Neutralization of Delta Variant with Sera of Covishield Vaccinees and COVID-19 Recovered Vaccinated Individuals. J. Travel Med. 2021, 28, taab119. [Google Scholar] [CrossRef] [PubMed]
- Wall, E.C.; Wu, M.; Harvey, R.; Kelly, G.; Warchal, S.; Sawyer, C.; Daniels, R.; Adams, L.; Hobson, P.; Hatipoglu, E.; et al. AZD1222-Induced Neutralising Antibody Activity against SARS-CoV-2 Delta VOC. Lancet 2021, 398, 207–209. [Google Scholar] [CrossRef]
- Barouch, D.H.; Stephenson, K.E.; Sadoff, J.; Yu, J.; Chang, A.; Gebre, M.; McMahan, K.; Liu, J.; Chandrashekar, A.; Patel, S.; et al. Durable Humoral and Cellular Immune Responses Following Ad26.COV2.S Vaccination for COVID-19. N. Engl. J. Med. 2021, 385, 951–953. [Google Scholar] [CrossRef]
- Jongeneelen, M.; Kaszas, K.; Veldman, D.; Huizingh, J.; van der Vlugt, R.; Schouten, T.; Zuijdgeest, D.; Uil, T.; van Roey, G.; Guimera, N.; et al. Ad26.COV2.S Elicited Neutralizing Activity against Delta and Other SARS-CoV-2 Variants of Concern. bioRxiv 2021. [Google Scholar] [CrossRef]
- Wall, E.C.; Wu, M.; Harvey, R.; Kelly, G.; Warchal, S.; Sawyer, C.; Daniels, R.; Hobson, P.; Hatipoglu, E.; Ngai, Y.; et al. Neutralising Antibody Activity against SARS-CoV-2 VOCs, B.1.617.2 and B.1.351 by BNT162b2 Vaccination. Lancet 2021, 397, 2331–2333. [Google Scholar] [CrossRef]
- Davis, C.; Logan, N.; Tyson, G.; Orton, R.; Harvey, W.; Haughney, J.; Perkins, J.; COVID-19 Genomics UK (COG-UK) Consortium; Peacock, T.P.; Barclay, W.S.; et al. Reduced Neutralisation of the Delta (B.1.617.2) SARS-CoV-2 Variant of Concern Following Vaccination. PLoS Pathog. 2021, 17, e1010022. [Google Scholar] [CrossRef]
- Choi, A.; Koch, M.; Wu, K.; Dixon, G.; Oestreicher, J.; Legault, H.; Stewart-Jones, G.B.E.; Colpitts, T.; Pajon, R.; Bennett, H.; et al. Serum Neutralizing Activity of MRNA-1273 against SARS-CoV-2 Variants. J. Virol. 2021, 95, e01313-21. [Google Scholar] [CrossRef]
- Yadav, P.D.; Sapkal, G.N.; Ella, R.; Sahay, R.R.; Nyayanit, D.A.; Patil, D.Y.; Deshpande, G.; Shete, A.M.; Gupta, N.; Mohan, V.K.; et al. Neutralization against B.1.351 and B.1.617.2 with Sera of COVID-19 Recovered Cases and Vaccinees of BBV152. bioRxiv 2021. [Google Scholar] [CrossRef]
- Vacharathit, V.; Aiewsakun, P.; Manopwisedjaroen, S.; Srisaowakarn, C.; Laopanupong, T.; Ludowyke, N.; Phuphuakrat, A.; Setthaudom, C.; Ekronarongchai, S.; Srichatrapimuk, S.; et al. CoronaVac Induces Lower Neutralising Activity against Variants of Concern than Natural Infection. Lancet Infect. Dis. 2021, 21, 1352–1354. [Google Scholar] [CrossRef]
- Niyomnaitham, S.; Toh, Z.Q.; Licciardi, P.V.; Wongprompitak, P.; Srisutthisamphan, K.; Copeland, K.K.; Chokephaibulkit, K. Immunogenicity of a Single Dose of BNT162b2, ChAdOx1 NCoV-19, or CoronaVac against SARS-CoV-2 Delta and Omicron Variants among Previously Infected Adults: A Randomized Trial. J. Infect. 2022, 85, 436–480. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Azze, R.; Chang-Monteagudo, A.; Climent-Ruiz, Y.; Macías-Abraham, C.; Valenzuela-Silva, C.; de los Ángeles García-García, M.; Jerez-Barceló, Y.; Triana-Marrero, Y.; Ruiz-Villegas, L.; Dairon Rodríguez-Prieto, L.; et al. Safety and Immunogenicity of the FINLAY-FR-1A Vaccine in COVID-19 Convalescent Participants: An Open-Label Phase 2a and Double-Blind, Randomised, Placebo-Controlled, Phase 2b, Seamless, Clinical Trial. Lancet Respir. Med. 2022, 10, 785–795. [Google Scholar] [CrossRef]
- Smolenov, I.; Han, H.H.; Li, P.; Baccarini, C.; Verhoeven, C.; Rockhold, F.; Clemens, S.A.C.; Ambrosino, D.; Richmond, P.; Siber, G.; et al. Impact of Previous Exposure to SARS-CoV-2 and of S-Trimer (SCB-2019) COVID-19 Vaccination on the Risk of Reinfection: A Randomised, Double-Blinded, Placebo-Controlled, Phase 2 and 3 Trial. Lancet. Infect. Dis. 2022, 22, 990–1001. [Google Scholar] [CrossRef]
- Duc Dang, A.; Dinh Vu, T.; Hai Vu, H.; Thanh Ta, V.; Thi Van Pham, A.; Thi Ngoc Dang, M.; Van Le, B.; Huu Duong, T.; Van Nguyen, D.; Lawpoolsri, S.; et al. Safety and Immunogenicity of an Egg-Based Inactivated Newcastle Disease Virus Vaccine Expressing SARS-CoV-2 Spike: Interim Results of a Randomized, Placebo-Controlled, Phase 1/2 Trial in Vietnam. Vaccine 2022, 40, 3621–3632. [Google Scholar] [CrossRef] [PubMed]
- Bernal, J.L.; Andrews, N.; Gower, C.; Gallagher, E.; Simmons, R.; Thelwall, S.; Stowe, J.; Tessier, E.; Groves, N.; Dabrera, G.; et al. Effectiveness of COVID-19 Vaccines against the B.1.617.2 Variant. N. Engl. J. Med. 2021, 385, 585–594. [Google Scholar] [CrossRef]
- Li, X. Omicron: Call for Updated Vaccines. J. Med. Virol. 2022, 94, 1261–1263. [Google Scholar] [CrossRef]
- Garcia-Beltran, W.F.; Denis, K.J.S.; Hoelzemer, A.; Lam, E.C.; Nitido, A.D.; Sheehan, M.L.; Berrios, C.; Ofoman, O.; Chang, C.C.; Hauser, B.M.; et al. MRNA-Based COVID-19 Vaccine Boosters Induce Neutralizing Immunity against SARS-CoV-2 Omicron Variant. Cell 2022, 185, 457.E4–466.E4. [Google Scholar] [CrossRef]
- Gruell, H.; Vanshylla, K.; Tober-Lau, P.; Hillus, D.; Schommers, P.; Lehmann, C.; Kurth, F.; Sander, L.E.; Klein, F. MRNA Booster Immunization Elicits Potent Neutralizing Serum Activity against the SARS-CoV-2 Omicron Variant. Nat. Med. 2022, 28, 477–480. [Google Scholar] [CrossRef]
- Wang, X.Y.; Mahmood, S.F.; Jin, F.; Cheah, W.K.; Ahmad, M.; Sohail, M.A.; Ahmad, W.; Suppan, V.K.; Sayeed, M.A.; Luxmi, S.; et al. Efficacy of Heterologous Boosting against SARS-CoV-2 Using a Recombinant Interferon-Armed Fusion Protein Vaccine (V-01): A Randomized, Double-Blind and Placebo-Controlled Phase III Trial. Emerg. Microbes Infect. 2022, 11, 1910–1919. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, X.; Li, S.; Dong, Y.; Liu, Y.; Li, J.; Zhao, Y.; Feng, Y. A Third High Dose of Inactivated COVID-19 Vaccine Induces Higher Neutralizing Antibodies in Humans against the Delta and Omicron Variants: A Randomized, Double-Blinded Clinical Trial. Sci. China Life Sci. 2022, 65, 1677–1679. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Jia, Z.; Bao, L.; Wang, L.; Cao, L.; Chi, H.; Hu, Y.; Li, Q.; Zhou, Y.; Jiang, Y.; et al. Memory B Cell Repertoire from Triple Vaccinees against Diverse SARS-CoV-2 Variants. Nature 2022, 603, 919. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Hwang, K.P.; Kuo, H.K.; Peng, W.J.; Shen, Y.H.; Kuo, B.S.; Huang, J.H.; Liu, H.; Ho, Y.H.; Lin, F.; et al. A Multitope SARS-CoV-2 Vaccine Provides Long-Lasting B Cell and T Cell Immunity against Delta and Omicron Variants. J. Clin. Investig. 2022, 132, e157707. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, X.; Song, J.; Wu, J.; Zhu, Y.; Li, M.; Cui, Y.; Chen, Y.; Yang, L.; Liu, J.; et al. Homologous or Heterologous Booster of Inactivated Vaccine Reduces SARS-CoV-2 Omicron Variant Escape from Neutralizing Antibodies. Emerg. Microbes Infect. 2022, 11, 477–481. [Google Scholar] [CrossRef]
- Lauring, A.S.; Tenforde, M.W.; Chappell, J.D.; Gaglani, M.; Ginde, A.A.; Mcneal, T.; Ghamande, S.; Douin, D.J.; Talbot, H.K.; Casey, J.D.; et al. Clinical Severity of, and Effectiveness of MRNA Vaccines against, COVID-19 from Omicron, Delta, and Alpha SARS-CoV-2 Variants in the United States: Prospective Observational Study. BMJ 2022, 376, e069761. [Google Scholar] [CrossRef]
- Luo, S.; Zhang, J.; Kreutzberger, A.J.B.; Eaton, A.; Edwards, R.J.; Jing, C.; Dai, H.-Q.; Sempowski, G.D.; Cronin, K.; Parks, R.; et al. Humanized Antibody Potently Neutralizes All SARS-CoV-2 Variants by a Novel Mechanism. bioRxiv 2022. [Google Scholar] [CrossRef]
- Luo, S.; Zhang, J.; Kreutzberger, A.J.B.; Eaton, A.; Edwards, R.J.; Jing, C.; Dai, H.-Q.; Sempowski, G.D.; Cronin, K.; Parks, R.; et al. An Antibody from Single Human V H -Rearranging Mouse Neutralizes All SARS-CoV-2 Variants Through BA.5 by Inhibiting Membrane Fusion. Sci. Immunol. 2022, 7, eadd5446. [Google Scholar] [CrossRef]
- O’Brien, M.P.; Forleo-Neto, E.; Musser, B.J.; Isa, F.; Chan, K.-C.; Sarkar, N.; Bar, K.J.; Barnabas, R.V.; Barouch, D.H.; Cohen, M.S.; et al. Subcutaneous REGEN-COV Antibody Combination to Prevent COVID-19. N. Engl. J. Med. 2021, 385, 1184–1195. [Google Scholar] [CrossRef]
- Bierle, D.M.; Ganesh, R.; Razonable, R.R. Breakthrough COVID-19 and Casirivimab-Imdevimab Treatment during a SARS-CoV-2 B1.617.2 (Delta) Surge. J. Clin. Virol. 2021, 145, 105026. [Google Scholar] [CrossRef]
- Huang, D.T.; McCreary, E.K.; Bariola, J.R.; Minnier, T.E.; Wadas, R.J.; Shovel, J.A.; Albin, D.; Marroquin, O.C.; Kip, K.E.; Collins, K.; et al. Effectiveness of Casirivimab-Imdevimab and Sotrovimab During a SARS-CoV-2 Delta Variant Surge: A Cohort Study and Randomized Comparative Effectiveness Trial. JAMA Netw. Open 2022, 5, e2220957. [Google Scholar] [CrossRef]
- Cicchitto, G.; Cardillo, L.; de Martinis, C.; Sabatini, P.; Marchitiello, R.; Abate, G.; Rovetti, A.; Cavallera, A.; Apuzzo, C.; Ferrigno, F.; et al. Effects of Casirivimab/Imdevimab Monoclonal Antibody Treatment among Vaccinated Patients Infected by SARS-CoV-2 Delta Variant. Viruses 2022, 14, 650. [Google Scholar] [CrossRef] [PubMed]
- Sasikala, M.; Sadhana, Y.; Vijayasarathy, K.; Gupta, A.; Daram, S.K.; Podduturi, N.C.R.; Reddy, D.N. Comparison of Saliva with Healthcare Workers- and Patient-Collected Swabs in the Diagnosis of COVID-19 in a Large Cohort. BMC Infect. Dis. 2021, 21, 648. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.; Kolata, G. President Trump Received Experimental Antibody Treatment. Available online: https://www.nytimes.com/2020/10/02/health/trump-antibody-treatment.html (accessed on 19 December 2021).
- Cunha, J.P. What Are the Symptoms of the COVID-19 Delta Variant? Available online: https://www.emedicinehealth.com/what_are_the_symptoms_of_covid19_delta_variant/article_em.htm (accessed on 16 December 2021).
- Connor, B.A.; Couto-Rodriguez, M.; Barrows, J.E.; Gardner, M.; Rogova, M.; O’Hara, N.B.; Nagy-Szakal, D. Monoclonal Antibody Therapy in a Vaccine Breakthrough SARS-CoV-2 Hospitalized Delta (B.1.617.2) Variant Case. Int. J. Infect. Dis. 2021, 110, 232–234. [Google Scholar] [CrossRef] [PubMed]
- Prathiviraj, R.; Chellapandi, P.; Begum, A.; Kiran, G.S.; Selvin, J. Identification of Genotypic Variants and Its Proteomic Mutations of Brazilian SARS-CoV-2 Isolates. Virus Res. 2022, 307, 198618. [Google Scholar] [CrossRef]
- Takashita, E.; Kinoshita, N.; Yamayoshi, S.; Sakai-Tagawa, Y.; Fujisaki, S.; Ito, M.; Iwatsuki-Horimoto, K.; Chiba, S.; Halfmann, P.; Nagai, H.; et al. Efficacy of Antibodies and Antiviral Drugs against COVID-19 Omicron Variant. N. Engl. J. Med. 2022, 386, 995–998. [Google Scholar] [CrossRef] [PubMed]
- Adlhoch, C.; de Carvalho Gomes, H. Sustainability of Surveillance Systems for SARS-CoV-2. Lancet Infect. Dis. 2022, 22, 914–915. [Google Scholar] [CrossRef]
- Nguyen, T.M.; Zhang, Y.; Pandolfi, P.P. Virus against Virus: A Potential Treatment for 2019-NCov (SARS-CoV-2) and Other RNA Viruses. Cell Res. 2020, 30, 189–190. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.M.S.; Mok, C.K.P.; Leung, Y.W.Y.; Ng, S.S.; Chan, K.C.K.; Ko, F.W.; Chen, C.; Yiu, K.; Lam, B.H.S.; Lau, E.H.Y.; et al. Neutralizing Antibodies against the SARS-CoV-2 Omicron Variant BA.1 Following Homologous and Heterologous CoronaVac or BNT162b2 Vaccination. Nat. Med. 2022, 28, 486–489. [Google Scholar] [CrossRef]
- Badano, M.N.; Sabbione, F.; Keitelman, I.; Pereson, M.; Aloisi, N.; Colado, A.; Ramos, M.V.; Ortiz Wilczyñski, J.M.; Pozner, R.G.; Castillo, L.; et al. Humoral Response to the BBIBP-CorV Vaccine over Time in Healthcare Workers with or without Exposure to SARS-CoV-2. Mol. Immunol. 2022, 143, 94–99. [Google Scholar] [CrossRef]
- Martinez, D.R.; Schäfer, A.; Gobeil, S.; Li, D.; De la Cruz, G.; Parks, R.; Lu, X.; Barr, M.; Stalls, V.; Janowska, K.; et al. A Broadly Cross-Reactive Antibody Neutralizes and Protects against Sarbecovirus Challenge in Mice. Sci. Transl. Med. 2022, 14, eabj7125. [Google Scholar] [CrossRef]
- Zhou, H.; Møhlenberg, M.; Thakor, J.C.; Tuli, H.S.; Wang, P.; Assaraf, Y.G.; Dhama, K.; Jiang, S. Sensitivity to Vaccines, Therapeutic Antibodies, and Viral Entry Inhibitors and Advances to Counter the SARS-CoV-2 Omicron Variant. Clin. Microbiol. Rev. 2022, 35, e00014-22. [Google Scholar] [CrossRef] [PubMed]
- Recombinant SARS-CoV-2 Delta-Omicron Variant Identified in the United States. Available online: https://www.news-medical.net/news/20220323/Recombinant-SARS-CoV-2-Delta-Omicron-variant-identified-in-the-United-States.aspx (accessed on 29 September 2022).
- Varabyou, A.; Pockrandt, C.; Salzberg, S.L.; Pertea, M. Rapid Detection of Inter-Clade Recombination in SARS-CoV-2 with Bolotie. Genetics 2020, 218, iyab074. [Google Scholar] [CrossRef] [PubMed]
- Lacek, K.A.; Rambo-Martin, B.L.; Batra, D.; Zheng, X.; Sakaguchi, H.; Peacock, T.; Keller, M.; Wilson, M.M.; Sheth, M.; Davis, M.L.; et al. Identification of a Novel SARS-CoV-2 Delta-Omicron Recombinant Virus in the United States. bioRxiv 2022. [Google Scholar] [CrossRef]
- Mahase, E. Unknown Covid-19: What Do We Know about the Delta Omicron Recombinant Variant? BMJ Br. Med. J. 2022, 376. [Google Scholar] [CrossRef]
- Bolze, A.; Basler, T.; White, S.; Rossi, A.D.; Wyman, D.; Roychoudhury, P.; Greninger, A.L.; Hayashibara, K.; Beatty, M.; Shah, S.; et al. Evidence for SARS-CoV-2 Delta and Omicron Co-Infections and Recombination. medRxiv 2022. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Deng, A.; Li, K.; Hu, Y.; Li, Z.; Xiong, Q.; Liu, Z.; Guo, Q.; Zou, L.; Zhang, H.; et al. Viral Infection and Transmission in a Large, Well-Traced Outbreak Caused by the SARS-CoV-2 Delta Variant. Nat. Commun. 2022, 13, 460. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ginn, H.M.; Dejnirattisai, W.; Supasa, P.; Wang, B.; Tuekprakhon, A.; Nutalai, R.; Zhou, D.; Mentzer, A.J.; Zhao, Y.; et al. Reduced Neutralization of SARS-CoV-2 B.1.617 by Vaccine and Convalescent Serum. Cell 2021, 184, 4220.e13–4236.e13. [Google Scholar] [CrossRef]
- Bates, T.A.; Leier, H.C.; Lyski, Z.L.; Goodman, J.R.; Curlin, M.E.; Messer, W.B.; Tafesse, F.G. Age-Dependent Neutralization of SARS-CoV-2 and P.1 Variant by Vaccine Immune Serum Samples. JAMA 2021, 326, 868–869. [Google Scholar] [CrossRef]
- Munro, A.P.S.; Janani, L.; Cornelius, V.; Aley, P.K.; Babbage, G.; Baxter, D.; Bula, M.; Cathie, K.; Chatterjee, K.; Dodd, K.; et al. Safety and Immunogenicity of Seven COVID-19 Vaccines as a Third Dose (Booster) Following Two Doses of ChAdOx1 NCov-19 or BNT162b2 in the UK (COV-BOOST): A Blinded, Multicentre, Randomised, Controlled, Phase 2 Trial. Lancet 2021, 398, 2258–2276. [Google Scholar] [CrossRef]
- Patel, M.K. Booster Doses and Prioritizing Lives Saved. N. Engl. J. Med. 2021, 385, 2476–2477. [Google Scholar] [CrossRef]
- Choi, A.; Koch, M.; Wu, K.; Chu, L.; Ma, L.; Hill, A.; Nunna, N.; Huang, W.; Oestreicher, J.; Colpitts, T.; et al. Safety and Immunogenicity of SARS-CoV-2 Variant MRNA Vaccine Boosters in Healthy Adults: An Interim Analysis. Nat. Med. 2021, 27, 2025–2031. [Google Scholar] [CrossRef] [PubMed]
- Jacob John, T. How India’s Second Covid Wave Started, What Went Wrong, and What Can Be Done: Top Virologist Answers. Available online: https://www.indiatoday.in/coronavirus-outbreak/story/india-second-covid-wave-start-went-wrong-top-virologist-answers-details-1795743-2021-04-28 (accessed on 20 May 2021).


| Sub Variants | Gene | Amino Acid Mutation | Mutation Impact | Omicron Variant Lineages |
|---|---|---|---|---|
| 21K (Omicron) | S | E484A | In other variants, mutations to this amino acid have been linked with antigenic escape, as well as mutations to other amino acids at this position [91]. | BA.1 |
| H655Y, N679K, P681H | May increase transmissibility [92]. | |||
| Q498R, N501Y | The binding to the ACE2 is increased by a significant amount [93]. | |||
| A67V, T95I, Y145D, H69, L212I, G339D, S371L, S373P, V70, S375F, K417N, N440K, G446S, G142, S477N, T478K, Q493R, G496S, V143, N501Y, Y505H, T547K, D614G, Y144, N764K, D796Y, N856K, Q954H, N211, N969K, L981F | ||||
| N | P13L | |||
| R203K, G204R | Viral loads and subgenomic RNA expression are increased [94,95]. | |||
| E31-, R32-, S33- | ||||
| ORF1a | K856R, S2083-, L2084I, I3758V, A2710T, T3255I, P3395H | |||
| G3676, L3674, S3675 | These mutations have been hypothesized to promote the evasion of innate immunity by impairing cells’ capacity to break down components of the virus [96]. | |||
| ORF1B | P314L, I1566V | |||
| ORF9b | P10S, E27-, N28-, A29- | By interacting with TOM70 and NEMO, ORF9b suppresses the innate immune response to infection, resulting in the generation of IFN [97,98]. | ||
| E | T91I | |||
| M | D3, Q19, A63T | |||
| 21L (Omicron) | S | T19I, V213G, T376A, D405N, S371F, R408S: (6 additional spike mutations) A27S, G142D, G339D, S373P, L24, S375F, K417N, N440K, P25, S477N, T478K, E484A, P26, Y505H, Q493R, Q498R, N501Y, N969K, D614G, H655Y, N679K, Q954H, P681H, N764K, D796Y | BA.2 | |
| N | P13L, E31, S413R, R32, S33, R203K, G204R | |||
| ORF1a | L3201F | May have originated in South Africa but it is more common in other countries than in the country of origin. | ||
| S135R, T842I, G1307S, F3677, L3027F, T3090I, T3255I, G3676, P3395H, S3675 | ||||
| ORF1b | P314L, T2163I, R1315C, I1566V | |||
| ORF3a | T223I | |||
| ORF6 | D61L | |||
| ORF9b | P10S, A29, E27, N28 | |||
| E | T9I | |||
| M | Q19E, A63T | |||
| 22A (Omicron) | S | F486V, R493Q | Due to the mutation and reversion, binding affinity to ACE2 is reduced [89]. | BA.4 |
| T19I, L24, N969K, P25, P26, Q954H, A27S, H69, D796Y, V70, N764K, G339D, S371F, P681H, S373P, S375F, N679K, T376A, D405N, H655Y, R408S, K417N, D614G, N440K, L452R, S477N, Y505H, T478K, E484A, F486V, Q498R, N501Y, | ||||
| N | S413R, P13L, E31, G204R, R32, S33, P151S, R203K | |||
| ORF1a | F3677, S135R, K141, G3676, S142, F143, S3675, T842I, G1307S, P3395H, L3027F, T3090I, T3255I | |||
| ORF1b | P314L, T2163I, R1315C, I1566V | |||
| ORF3a | T223I | |||
| ORF6 | D61L | |||
| ORF7b | L11F | |||
| ORF9b | P10S, E27-, N28-, A29- | |||
| E | T9I | |||
| M | Q19E, A63T | |||
| 22B (Omicron) | S | T19I, L24-, N969K, P25-, P26-, Q954H, A27S, H69-, D796Y, V70-, G142D, V213G, N764K, G339D, S371F, P681H, S373P, S375F, T376A, N679K, D405N, R408S, H655Y, D614G, K417N, N440K, L452R, S477N, Y505H, T478K, E484A, F486V, N501Y, Q498R | A reversion at R493Q and the F486V mutation may have caused the marked reduction in ACE2 binding affinity in 22A (Omicron) and 22B (Omicron) compared to 21K (Omicron) [89]. | BA.5 |
| N | P13L, E31, S413R, R32-, S33-, G204R, R203K | |||
| ORF1a | S135R, T842I, F3677, G1307S, L3027F, G3676, T3090I, T3255I, S3675, P3395H | |||
| ORF1b | P314L, T2163I, R1315C, I1566V | |||
| ORF3a | T223I | |||
| ORF9b | P10S, A29, E27, N28 | |||
| E | T9I | |||
| M | A63T, D3N, Q19E | |||
| 22C (Omicron) | S | N969K, T19I, L24, Q954H, P25, P26, A27S, D796Y, G142D, V213G, N764K, G339D, S704L, S371F, S373P, P681H, S375F, T376A, N679K, D405N, R408S, K417N, H655Y, N440K, L452Q, D614G, S477N, T478K, Y505H, E484A, Q493R, Q498R, N501Y | BA.2.12.1 | |
| N | P13L, E31, G204R, S413R, R32, R203K, S33 | |||
| ORF1a | S135R, T842I, F3677, G1307S, L3027F, G3676, T3090I, L3201F, S3675, T3255I, P3395H | L3201 originated in South Africa as a wild type but it is common in other countries. | ||
| ORF1b | P314L, R1315C, T2163I, I1566V | |||
| ORF3a | T223I | |||
| ORF6 | D61L | |||
| ORF9b | P10S, A29, E27, N28 | |||
| E | T9I | |||
| M | Q19E, A63T | |||
| 22D (Omicron) | S | T19I, L24, N969K, P25, P26, Q954H, A27S, D796Y, G142D, N764K, K147E, D796K, P681H, W152R, N679K, F157L, I210V, H655Y, I210V, V213G, D614G, G257S, G339H, Y505H, SS373P, S375F, N501Y, T376A, D405N, Q498R, R408S, K417N, R493Q, N440K, G446S, E484A, N460K, S477N, T478K | Mutations in N460K, G446S, G339H, and R493Q may cause 21L(Omicron)-induced immunity to be overcome. May also have higher ACE-2 binding affinity than 22A (Omicron)/22B (Omicron) [89]. | BA.2.75 |
| N | P13L, E31, S413R, R32, S33, R203K, G204R | |||
| ORF1a | S135R, T842I, N4060S, S1221L, G1307S, F3677, P1640S, L3027F, G3676, T3090I, L3201F, S3675, T3255I, P3395H | |||
| ORF1b | P314L, G662S, T2163I, R1315C, I1566V | |||
| ORF3a | T223I | |||
| ORF6 | D61L | |||
| ORF9b | P10S, A29, E27, N28 | |||
| E | T9I, T11A | |||
| M | Q19E, A63T |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chavda, V.P.; Bezbaruah, R.; Deka, K.; Nongrang, L.; Kalita, T. The Delta and Omicron Variants of SARS-CoV-2: What We Know So Far. Vaccines 2022, 10, 1926. https://doi.org/10.3390/vaccines10111926
Chavda VP, Bezbaruah R, Deka K, Nongrang L, Kalita T. The Delta and Omicron Variants of SARS-CoV-2: What We Know So Far. Vaccines. 2022; 10(11):1926. https://doi.org/10.3390/vaccines10111926
Chicago/Turabian StyleChavda, Vivek P., Rajashri Bezbaruah, Kangkan Deka, Lawandashisha Nongrang, and Tutumoni Kalita. 2022. "The Delta and Omicron Variants of SARS-CoV-2: What We Know So Far" Vaccines 10, no. 11: 1926. https://doi.org/10.3390/vaccines10111926
APA StyleChavda, V. P., Bezbaruah, R., Deka, K., Nongrang, L., & Kalita, T. (2022). The Delta and Omicron Variants of SARS-CoV-2: What We Know So Far. Vaccines, 10(11), 1926. https://doi.org/10.3390/vaccines10111926