
Flagellin-Fused Protein Targeting M2e and HA2 Induces Innate and T-Cell Responses in Mice of Different Genetic Lines

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Recombinant Protein
2.2. Mice
2.3. Immunization
2.4. Collection of Mouse Lungs
2.5. Phenotyping of Innate Immunity Cells Using Multiparameter Flow Cytometry
2.6. Intracellular Cytokine Staining (ICS) Assay
2.7. Statistical Analysis
3. Results
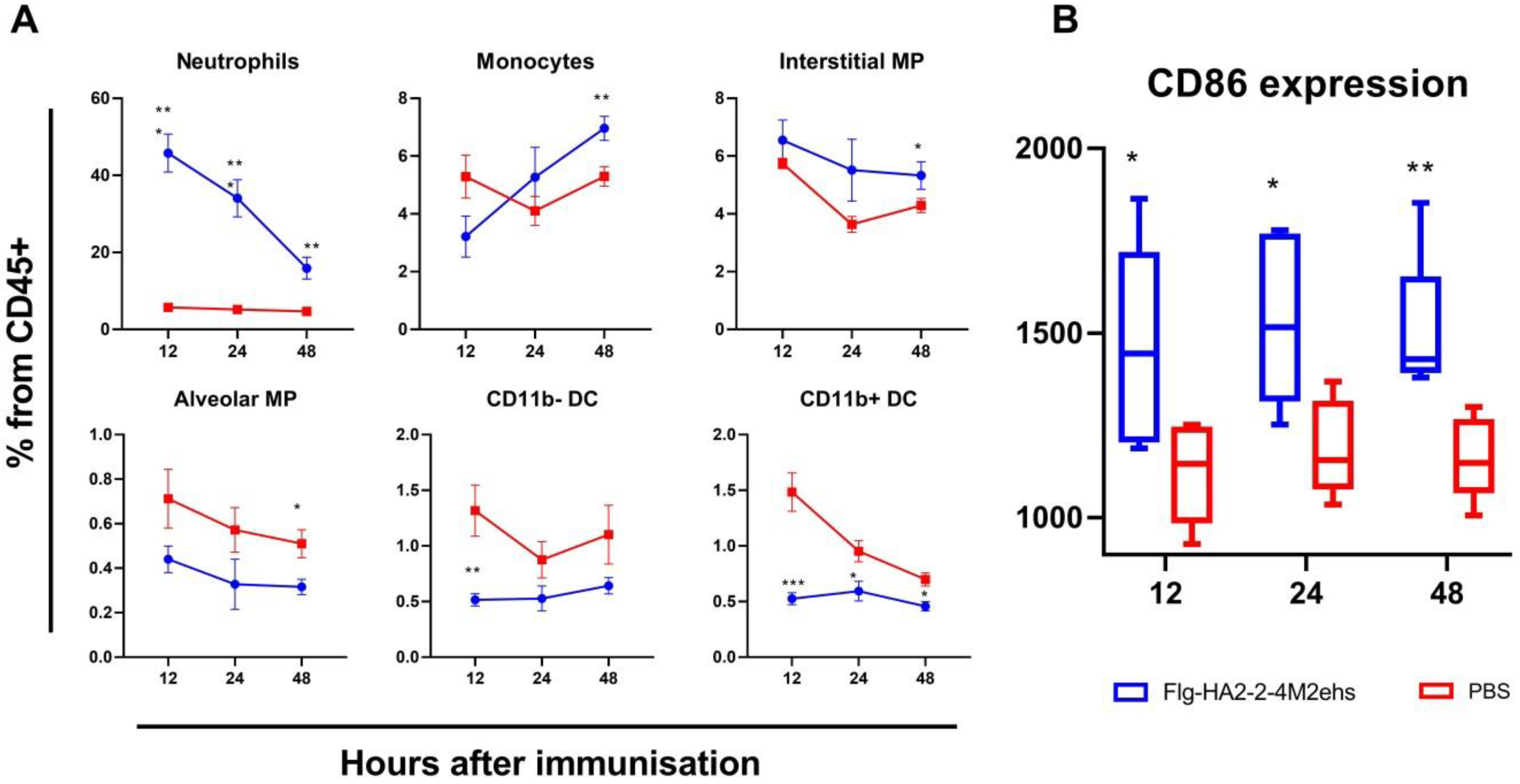
3.1. Dynamics of the Main Populations of Innate Immunity Cells in the Lungs
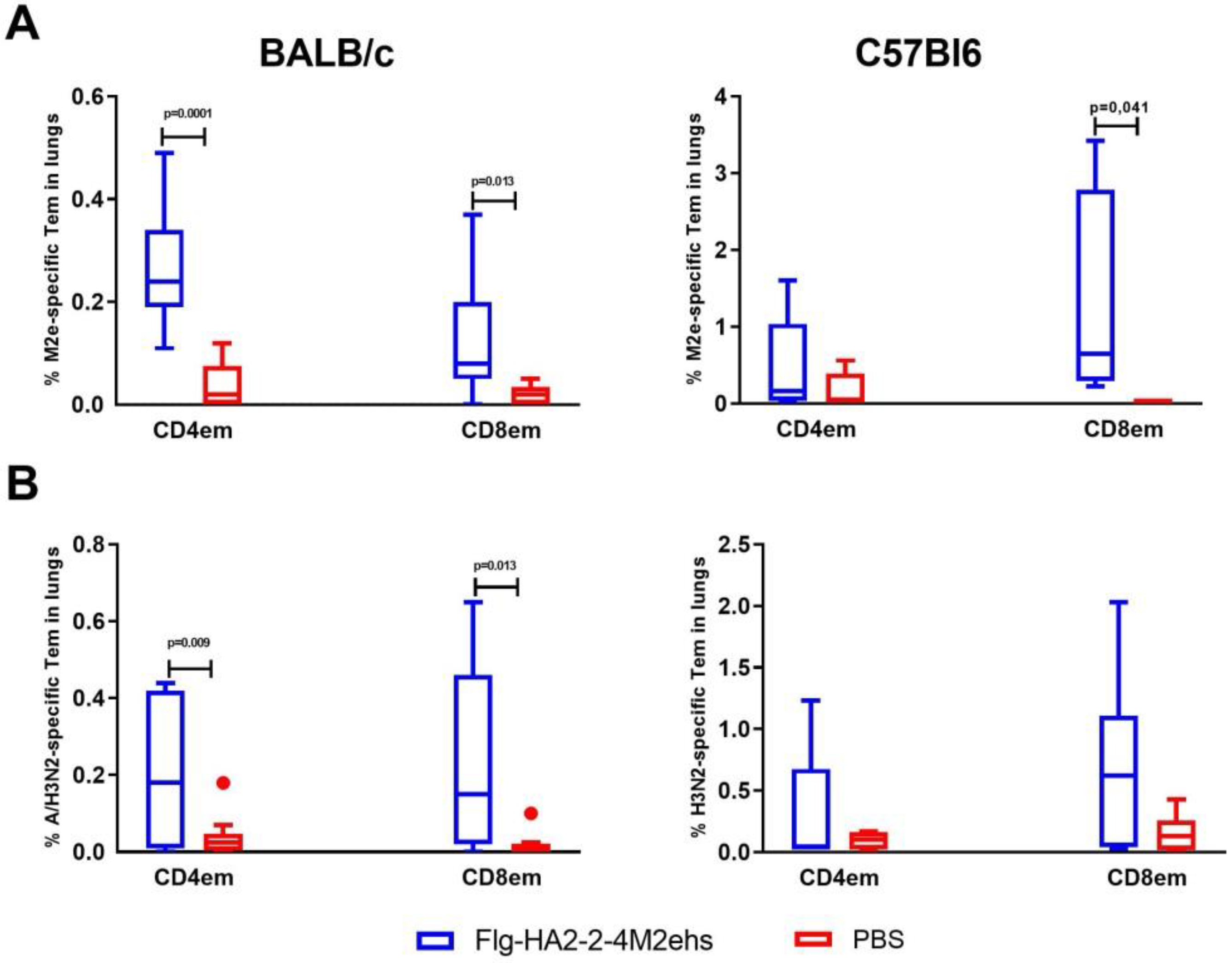
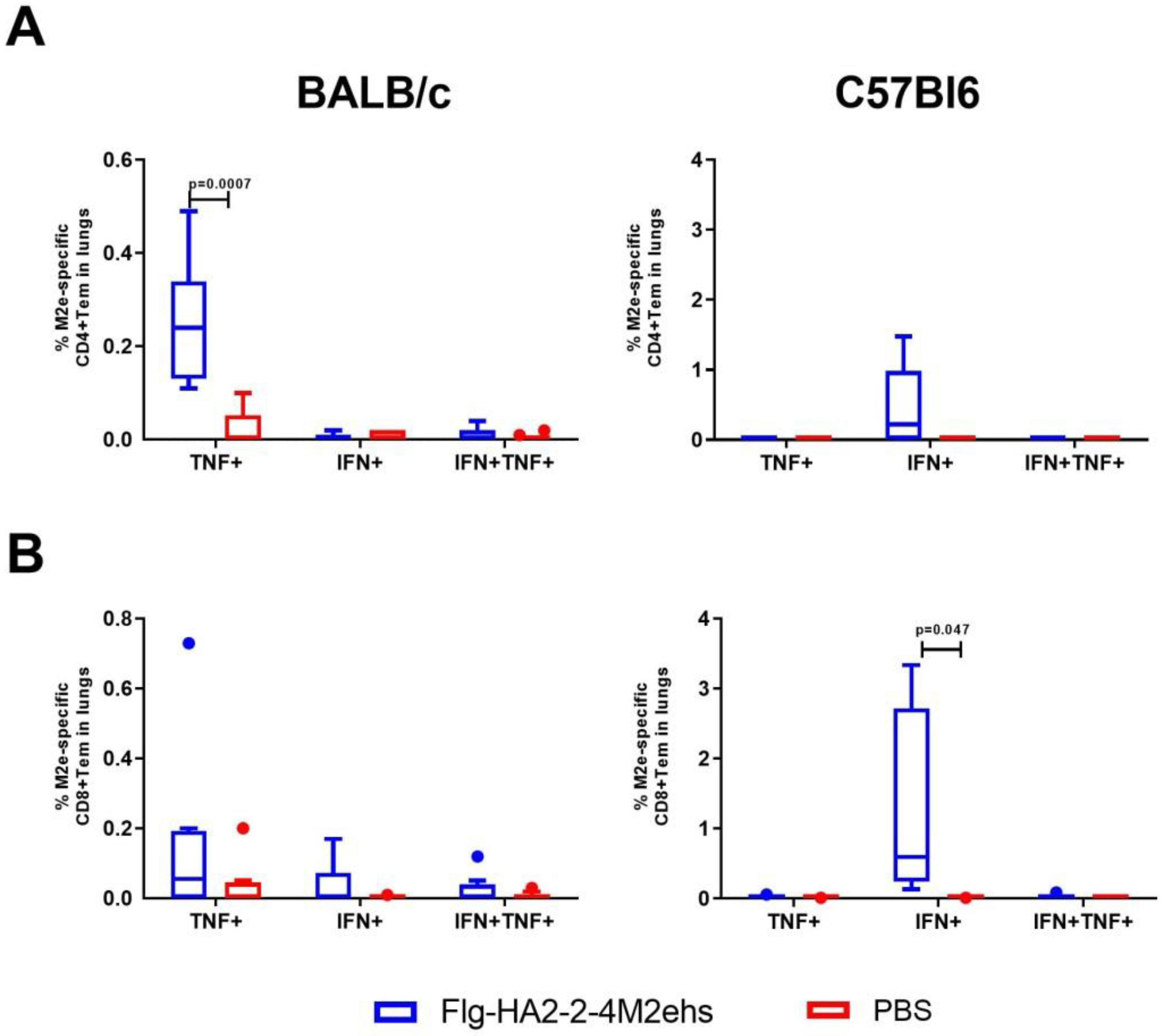
3.2. M2e-Specific T-Cell Response in the Lungs of BALB/c and C57Bl6 Mice after Intranasal Immunization
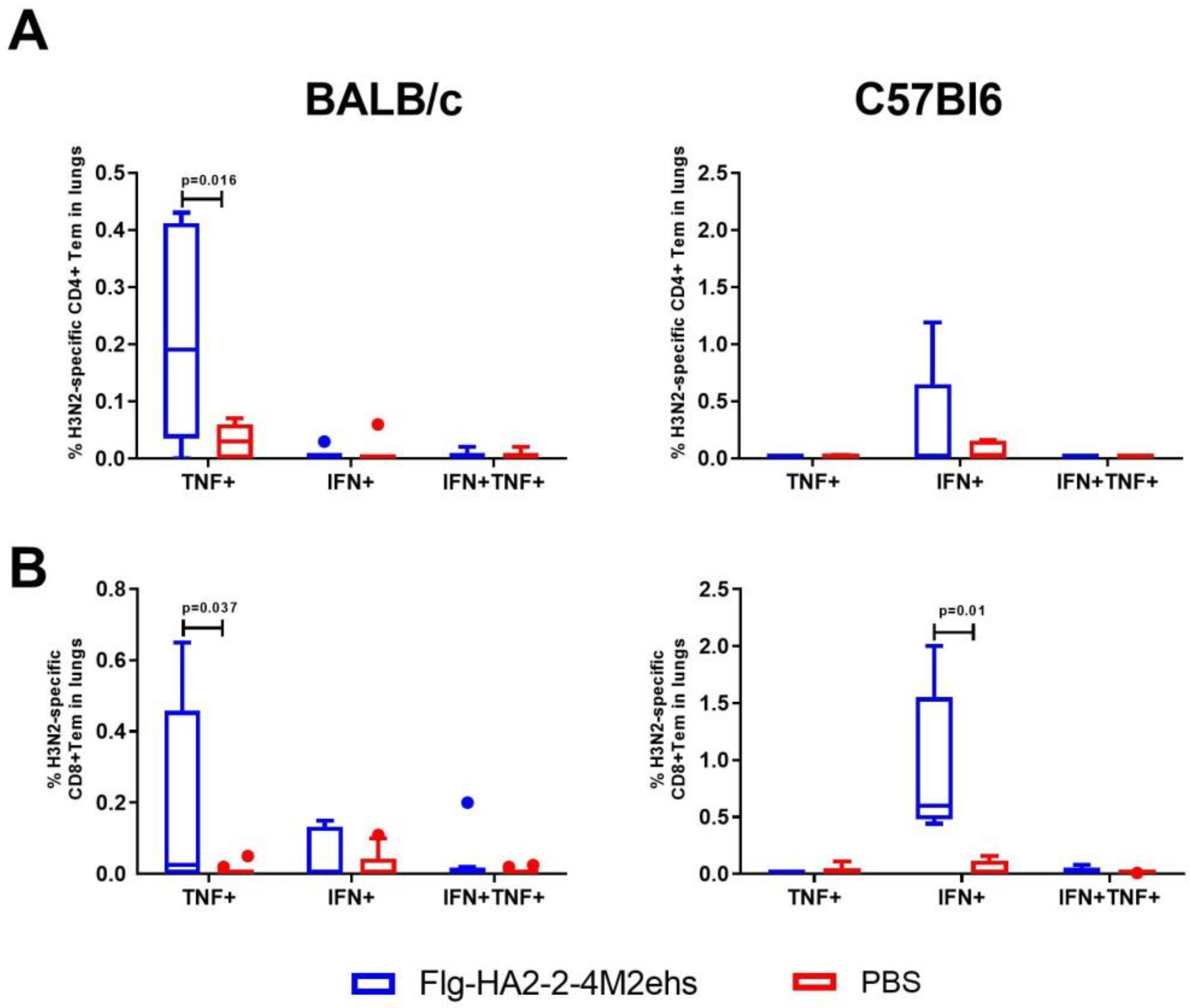
3.3. A/H3N2-Specific T-Cell Response in the Lungs of BALB/c and C57Bl6 Mice after Intranasal Immunization
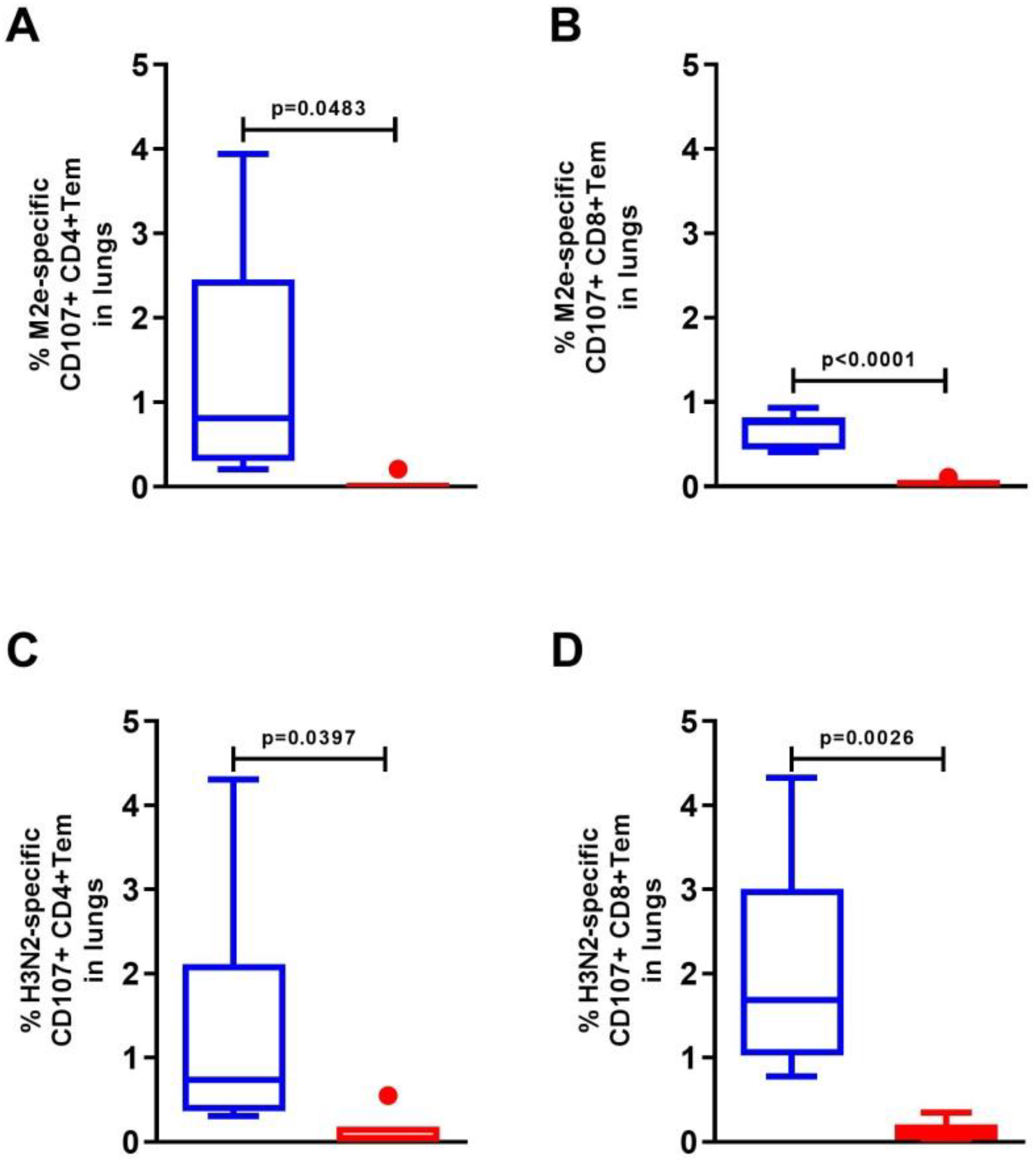
3.4. Expression of the CD107a+ Marker in the Lung Cells of C57Bl/6 Mice after Intranasal Immunization
4. Discussion
5. Conclusions
6. Patent
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HA | hemagglutinin |
| HA2 | second subunit of hemagglutinin |
| M2e | ectodomen M2 protein |
| Flg | flagellin from Salmonella typhimurium |
| IPTG | isopropyl-β-D-1-thiogalactopyranoside |
| PBS | phosphate buffer solution |
| Tem | memory effector T cells |
| MFI | median fluorescence intensity |
| SEM | standard error of the mean |
| RSV | respiratory syncytial virus |
| RV | rhinovirus |
| HMPV | metapneumovirus |
| TLR-5 | toll-like receptor 5 |
| CTL | cytotoxic T cells |
References
- Paules, C.I.; Marston, H.D.; Eisinger, R.W.; Baltimore, D.; Fauci, A.S. The Pathway to a Universal Influenza Vaccine. Immunity 2017, 47, 599–603. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.; Lapuente, D. T Cell Immunity against Influenza: The Long Way from Animal Models Towards a Real-Life Universal Flu Vaccine. Viruses 2021, 13, 199. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zheng, D.; Li, C.; Zhang, W.; Xu, W.; Liu, X.; Fang, F.; Chen, Z. Protection against multiple subtypes of influenza viruses by virus-like particle vaccines based on a hemagglutinin conserved epitope. Biomed. Res. Int. 2015, 2015, 901817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scorza, B.F.; Tsvetnitsky, V.; Donnelly, J.J. Universal influenza vaccines: Shifting to better vaccines. Vaccine 2016, 34, 2926–2933. [Google Scholar] [CrossRef] [Green Version]
- van Doorn, E.; Liu, H.; Ben-Yedidia, T.; Hassin, S.; Visontai, I.; Norley, S.; Frijlink, H.W.; Hak, E. Evaluating the immunogenicity and safety of a BiondVax-developed universal influenza vaccine (Multimeric-001) either as a standalone vaccine or as a primer to H5N1 influenza vaccine Phase IIb study protocol. Medicine 2017, 96, e6339. [Google Scholar] [CrossRef] [PubMed]
- Saelens, X. The role of matrix protein 2 ectodomain in the development of universal influenza vaccines. J. Infect. Dis. 2019, 219, S68–S74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pleguezuelos, O.; James, E.; Fernandez, A.; Lopes, V.; Rosas, L.A.; Cervantes-Medina, A.; Cleath, J.; Edwards, K.; Neitzey, D.; Gu, W.; et al. Efficacy of FLU-v, a broad-spectrum influenza vaccine, in a randomized phase IIb human influenza challenge study. NPJ Vaccines 2020, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Shinde, V.; Cai, R.; Plested, J.; Cho, I.; Fiske, J.; Pham, X.; Zhu, M.; Cloney-Clark, S.; Wang, N.; Zhou, H.; et al. Induction of Cross-Reactive Hemagglutination Inhibiting Antibody and Polyfunctional CD4+ T-Cell Responses by a Recombinant Matrix-M–Adjuvanted Hemagglutinin Nanoparticle Influenza Vaccine. Clin. Inf. Dis. 2021, 73, e4278–e4287. [Google Scholar] [CrossRef]
- Li, Y.; Xu, M.; Li, Y.; Gu, W.; Halimu, G.; Li, Y.; Zhang, Z.; Zhou, L.; Liao, H.; Yao, S.; et al. A recombinant protein containing influenza viral conserved epitopes and superantigen induces broad-spectrum protection. eLife 2021, 10, e71725. [Google Scholar] [CrossRef]
- Tsybalova, L.M.; Stepanova, L.A.; Shuklina, M.A.; Mardanova, E.S.; Kotlyarov, R.Y.; Potapchuk, M.V.; Petrov, S.A.; Blokhina, E.A.; Ravin, N.V. Combination of M2e peptide with stalk HA epitopes of influenza A virus enhances protective properties of recombinant vaccine. PLoS ONE 2018, 13, e0201429. [Google Scholar] [CrossRef]
- Stepanova, L.A.; Mardanova, E.S.; Shuklina, M.A.; Blokhina, E.A.; Kotlyarov, R.Y.; Potapchuk, M.V.; Kovaleva, A.A.; Vidyaeva, I.G.; Korotkov, A.V.; Eletskaya, E.I.; et al. Flagellin-fused protein targeting M2e and HA2 induces potent humoral and T-cell respones and protects mice against various influenza viruses a subtypes. J. Biomed. Sci. 2018, 25, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuklina, M.A.; Stepanova, L.A.; Kovaleva, A.A.; Korotkov, A.V.; Shaldzhyan, A.A.; Zaitseva, M.V.; Eletskaya, E.I.; Tsybalova, L.M. Intranasal immunization with a recombinant protein based on the M2e peptide and second subunit of influenza A viral hemagglutinin fragment induces a cross-protective humoral and T cell response in mice. Med. Immunol. 2020, 22, 357–370. [Google Scholar] [CrossRef] [Green Version]
- Jegerlehner, A.; Schmitz, N.; Storni, T.; Bachmann, M.F. Influenza a vaccine based on the extracellular domain of M2: Weak protection mediated via antibodydependent NK cell activity. J. Immunol. 2004, 172, 5598–5605. [Google Scholar] [CrossRef] [Green Version]
- El Bakkouri, K.; Descamps, F.; de Filette, M.; Smet, A.; Festjens, E.; Birkett, A.; van Rooijen, N.; Verbeek, S.; Fiers, W.; Saelens, X. Universal vaccine based on ectodomain of matrix protein 2 of influenza a: Fc receptors and alveolar macrophages mediate protection. J. Immunol. 2011, 186, 1022–1031. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.-N.; Kim, M.-C.; Lee, Y.-T.; Kim, Y.-J.; Kang, S.-M. Mechanisms of Cross-protection by Influenza Virus M2-based Vaccines. Immune Netw. 2015, 15, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolpe, A.; Schepens, B.; Fiers, W.; Saelens, X. M2-based influenza vaccines: Recent advances and clinical potential. Expert Rev. Vaccines 2017, 16, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Gerhard, W.; Mozdzanowska, K.; Zharikova, D. Prospects for universal influenza virus vaccine. Emerg. Infect. Dis. 2006, 12, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S.F.; Graham, B.S.; Mascola, J.R.; McDermott, A.B. Is it possible to develop a “universal” influenza virus vaccine? Immunogenetic considerations underlying B-cell biology in the development of a pan-subtype influenza A vaccine targeting the hemagglutinin stem. Cold Spring Harb. Perspect. Biol. 2018, 10, a029413. [Google Scholar] [CrossRef]
- Andrews, S.F.; Joyce, M.G.; Chambers, M.J.; Gillespie, R.A.; Kanekiyo, M.; Leung, K.; Yang, E.S.; Tsybovsky, Y.; Wheatley, A.K.; Crank, M.C.; et al. Preferential induction of cross-group influenza A hemagglutinin stem-specific memory B cells after H7N9 immunization in humans. Sci. Immunol. 2017, 2, eaan2676. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.T.; Tan, G.S.; Hai, R.; Pica, N.; Petersen, E.; Moran, T.M.; Palese, P. Broadly protective monoclonal antibodies against H3 influenza viruses following sequential immunization with different hemagglutinins. PLoS Pathog. 2010, 6, e1000796. [Google Scholar] [CrossRef]
- Bommakanti, G.; Citron, M.P.; Hepler, R.W.; Callahan, C.; Heidecker, G.J.; Najar, T.A.; Lu, X.; Joyce, J.G.; Shiver, J.W.; Casimiro, D.R.; et al. Design of an HA2-based Escherichia coli expressed influenza immunogen that protects mice from pathogenic challenge. Proc. Natl. Acad. Sci. USA 2010, 107, 13701–13706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneemann, A.; Speir, J.A.; Tan, G.S.; Khayat, R.; Ekiert, D.C.; Matsuoka, Y.; Wilson, I.A. A virus-like particle that elicits cross-reactive antibodies to the conserved stem of influenza virus hemagglutinin. J. Virol. 2012, 86, 11686–11697. [Google Scholar] [CrossRef] [Green Version]
- Staneková, Z.; Adkins, I.; Kosová, M.; Janulíková, J.; Šebo, P.; Varečková, E. Heterosubtypic protection against influenza A induced by adenylate cyclase toxoids delivering conserved HA2 subunit of hemagglutinin. Antivir. Res. 2013, 97, 24–35. [Google Scholar] [CrossRef]
- Stepanova, L.A.; Sergeeva, M.V.; Shuklina, M.A.; Shaldzhyan, A.A.; Potapchuk, M.V.; Korotkov, A.V.; Tsybalova, L.M. A Fusion Protein Based on the Second Subunit of Hemagglutinin of Influenza A/H2N2 Viruses Provides Cross Immunity. Acta Nat. 2016, 8, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.-Y.; Wang, S.-C.; Ko, Y.-A.; Lin, K.-I.; Ma, C.; Cheng, T.-J.R.; Wong, C.-H. Chimeric hemagglutinin vaccine elicits broadly protective CD4 and CD8 T cell responses against multiple influenza strains and subtypes. Proc. Natl. Acad. Sci. USA 2020, 117, 17757–17763. [Google Scholar] [CrossRef]
- Yu, Y.R.A.; O’Koren, E.G.; Hotten, D.F.; Kan, M.J.; Kopin, D.; Nelson, E.R.; Que, L.; Gunn, M.D. A protocol for the comprehensive flow cytometric analysis of immune cells in normal and inflamed murine non-lymphoid tissues. PLoS ONE 2016, 11, e0150606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, Y.H.; Seong, B.L. Call for a paradigm shift in the design of universal influenza vaccines by harnessing multiple correlates of protection. Expert Opin. Drug Discov. 2020, 15, 1441–1455. [Google Scholar] [CrossRef]
- Tscharke, D.C.; Croft, N.P.; Doherty, P.C.; La Gruta, N.L. Sizing up the key determinants of the CD8(+) T cell response. Nat. Rev. Immunol. 2015, 15, 705–716. [Google Scholar] [CrossRef]
- Lukacs, N.W.; Malinczak, C.-A. Harnessing Cellular Immunity for Vaccination against Respiratory Viruses. Vaccines 2020, 8, 783. [Google Scholar] [CrossRef]
- Hillaire, M.L.B.; Osterhaus, A.D.M.E.; Rimmelzwaan, G.F. Induction of virus-specific cytotoxic T lymphocytes as a basis for the development of broadly protective influenza vaccines. J. Biomed. Biotechnol. 2011, 2011, 939860. [Google Scholar] [CrossRef]
- McKinstry, K.K.; Strutt, T.M.; Kuang, Y.; Brown, D.M.; Sell, S.; Dutton, R.W.; Swain, S.L. Memory CD4+ T cells protect against influenza through multiple synergizing mechanisms. J. Clin. Investig. 2012, 122, 2847–2856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, T.M.; Li, C.K.; Chui, C.S.; Huang, A.K.; Perkins, M.; Liebner, J.C.; Lambkin-Williams, R.; Gilbert, A.; Oxford, J.; Nicholas, B.; et al. Preexisting influenza-specific CD4+ T cells correlate with disease protection against influenza challenge in humans. Nat. Med. 2012, 18, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Begom, S.; Bermingham, A.; Hoschler, K.; Adamson, W.; Carman, W.; Bean, T.; Barclay, W.; Deeks, J.J.; Lalvani, A. Cellular immune correlates of protection against symptomatic pandemic influenza. Nat. Med. 2013, 19, 1305–1312. [Google Scholar] [CrossRef]
- Altenburg, A.F.; Rimmelzwaan, G.F.; de Vries, R.D. Virus-specific T cells as correlate of (cross-)protective immunity against influenza. Vaccine 2015, 33, 500–506. [Google Scholar] [CrossRef]
- Eliasson, D.G.; Omokanye, A.; SchoËn, K.; Wenzel, U.A.; Bernasconi, V.; Bemark, M.; Kolpe, A.; Bakkouri, K.E.; Ysenbaert, T.; Deng, L.; et al. M2etetramer-specific memory CD4 T cells are broadly protective against influenza infection. Mucosal Immunol. 2018, 11, 273–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martorelli, D.; Muraro, E.; Merlo, A.; Turrini, R.; Rosato, A.; Dolcetti, R. Role of CD4+ cytotoxic T lymphocytes in the control of viral diseases and cancer. Int. Rev. Immunol. 2010, 29, 371–402. [Google Scholar] [CrossRef]
- Juno, J.A.; van Bockel, D.; Stephen, J.; Kent, S.J.; Anthony, D.; Kelleher, A.D.; Zaunders, J.J.; Munier, C.M.L. Cytotoxic CD4 T Cells-Friend or Foe during Viral Infection? Front. Immunol. 2017, 23, 19. [Google Scholar] [CrossRef] [Green Version]
- Meckiff, B.J.; Ladell, K.; McLaren, J.E.; Ryan, G.B.; Leese, A.M.; James, E.A.; Price, D.A.; Long, H.M. Primary EBV Infection Induces an Acute Wave of Activated Antigen-Specific Cytotoxic CD4 + T Cells. J. Immunol. 2019, 203, 1276–1287. [Google Scholar] [CrossRef] [Green Version]
- van de Ven, K.; de Heij, F.; van Dijken, H.; Ferreira, J.A.; de Jonge, J. Systemic and respiratory T-cells induced by seasonal H1N1 influenza protect against pandemic H2N2 in ferrets. Commun. Biol. 2020, 3, 564. [Google Scholar] [CrossRef]
- Stewart, D.; Fulton, W.D.; Wilson, C.; Monitto, C.L.; Paidas, C.N.; Reeves, R.H.; de Maio, A. Genetic contribution to the septic response in a mouse model. Shock 2002, 18, 342–347. [Google Scholar] [CrossRef]
- Lee, J.; Kim, J.; Lee, J.; Shin, S.J.; Shin, E.-C. DNA immunization of Mycobacterium tuberculosis resuscitation-promoting factor B elicits polyfunctional CD8+ T cell responses. Clin. Exp. Vaccine Res. 2014, 3, 235–243. [Google Scholar] [CrossRef] [Green Version]
- Perdomo-Celis, F.; Velilla, P.A.; Taborda, N.A.; Rugeles, T.M. An altered cytotoxic program of CD8+ T-cells in HIV-infected patients despite HAART induced viral suppression. PLoS ONE 2019, 14, e0210540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, R.; Coulon, P.-G.A.; Prakash, S.; Dhanushkodi, N.R.; Roy, S.; Nguyen, A.M.; Alomari, N.I.; Mai, U.T.; Amezquita, C.; Ye, C.; et al. Human epitopes identified from herpes simplex virus tegument protein VP11/12 (UL46) recall multifunctional effector memory CD4+ TEM cells in asymptomatic individuals and protect from ocular herpes infection and disease in “humanized” HLA-DR transgenic mice. J. Virol. 2020, 94, e01991-19. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.M.; Lee, S.; Garcia-Hernandez, M.D.L.L.; Swain, S.L. Multifunctional CD4 cells expressing gamma interferon and perforin mediate protection against lethal influenza virus infection. J. Virol. 2012, 86, 6792–6803. [Google Scholar] [CrossRef] [Green Version]
- Savic, M.; Dembinski, J.L.; Laake, I.; Hungnes, O.; Cox, R.; Oftung, F.; Trogstad, L.; Mjaaland, S. Distinct T and NK cell populations may serve as immune correlates of protection against symptomatic pandemic influenza A(H1N1) virus infection during pregnancy. PLoS ONE 2017, 12, e0188055. [Google Scholar] [CrossRef] [Green Version]
- L’Huillier, A.G.; Ferreira, V.H.; Hirzel, C.; Nellimarla, S.; Ku, T.; Natori, Y.; Humar, A.; Kumar, D. T-cell responses following Natural Influenza Infection or Vaccination in Solid Organ Transplant Recipients. Sci. Rep. 2020, 10, 10104. [Google Scholar] [CrossRef]
- Hillaire, M.L.; Rimmelzwaan, G.F.; Kreijtz, J.H. Clearance of influenza virus infections by T cells: Risk of collateral damage? Curr. Opin. Virol. 2013, 3, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Thomas, P.G. Balancing immune protection and immune pathology by CD8(1) T cell responses to influenza infection. Front. Immunol. 2016, 7, 25. [Google Scholar] [CrossRef] [Green Version]
- Dhakal, S.; Loube, J.; Misplon, J.A.; Lo, C.-Y.; Creisher, P.S.; Mulka, K.R.; Deshpande, S.; Mitzner, W.; Klein, S.L.; Epstein, S.L. Effect of an Adenovirus-Vectored Universal Influenza Virus Vaccine on Pulmonary Pathophysiology in a Mouse Model. J. Virol. 2021, 95, e02359-20. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse Number | ||||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |
| M2e-specific CD8+Tem | ||||||
| CD107a+ | + | + | + | + | − | + |
| IFN-γ+ | − | ++ | ++ | − | + | − |
| IFN+CD107a+ | + | + | − | + | + | + |
| H3N2-specific CD8+Tem | ||||||
| CD107a+ | ++ | ++ | + | ++ | + | ++ |
| IFN-γ+ | − | ++ | ++ | − | − | + |
| IFN+CD107a+ | + | + | − | − | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stepanova, L.A.; Shuklina, M.A.; Vasiliev, K.A.; Kovaleva, A.A.; Vidyaeva, I.G.; Zabrodskaya, Y.A.; Korotkov, A.V.; Tsybalova, L.M. Flagellin-Fused Protein Targeting M2e and HA2 Induces Innate and T-Cell Responses in Mice of Different Genetic Lines. Vaccines 2022, 10, 2098. https://doi.org/10.3390/vaccines10122098
Stepanova LA, Shuklina MA, Vasiliev KA, Kovaleva AA, Vidyaeva IG, Zabrodskaya YA, Korotkov AV, Tsybalova LM. Flagellin-Fused Protein Targeting M2e and HA2 Induces Innate and T-Cell Responses in Mice of Different Genetic Lines. Vaccines. 2022; 10(12):2098. https://doi.org/10.3390/vaccines10122098
Chicago/Turabian StyleStepanova, Liudmila A., Marina A. Shuklina, Kirill A. Vasiliev, Anna A. Kovaleva, Inna G. Vidyaeva, Yana A. Zabrodskaya, Alexandr V. Korotkov, and Liudmila M. Tsybalova. 2022. "Flagellin-Fused Protein Targeting M2e and HA2 Induces Innate and T-Cell Responses in Mice of Different Genetic Lines" Vaccines 10, no. 12: 2098. https://doi.org/10.3390/vaccines10122098
APA StyleStepanova, L. A., Shuklina, M. A., Vasiliev, K. A., Kovaleva, A. A., Vidyaeva, I. G., Zabrodskaya, Y. A., Korotkov, A. V., & Tsybalova, L. M. (2022). Flagellin-Fused Protein Targeting M2e and HA2 Induces Innate and T-Cell Responses in Mice of Different Genetic Lines. Vaccines, 10(12), 2098. https://doi.org/10.3390/vaccines10122098