
Establishment of Sandwich ELISA for Quality Control in Rotavirus Vaccine Production

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression and Purification of P[4], P[6], P[8]-VP4*
2.2. Monoclonal Antibody Generation and Production
2.3. Enzyme-Linked Immunosorbent Assay
2.4. Vaccine Adsorption Experiment
3. Results
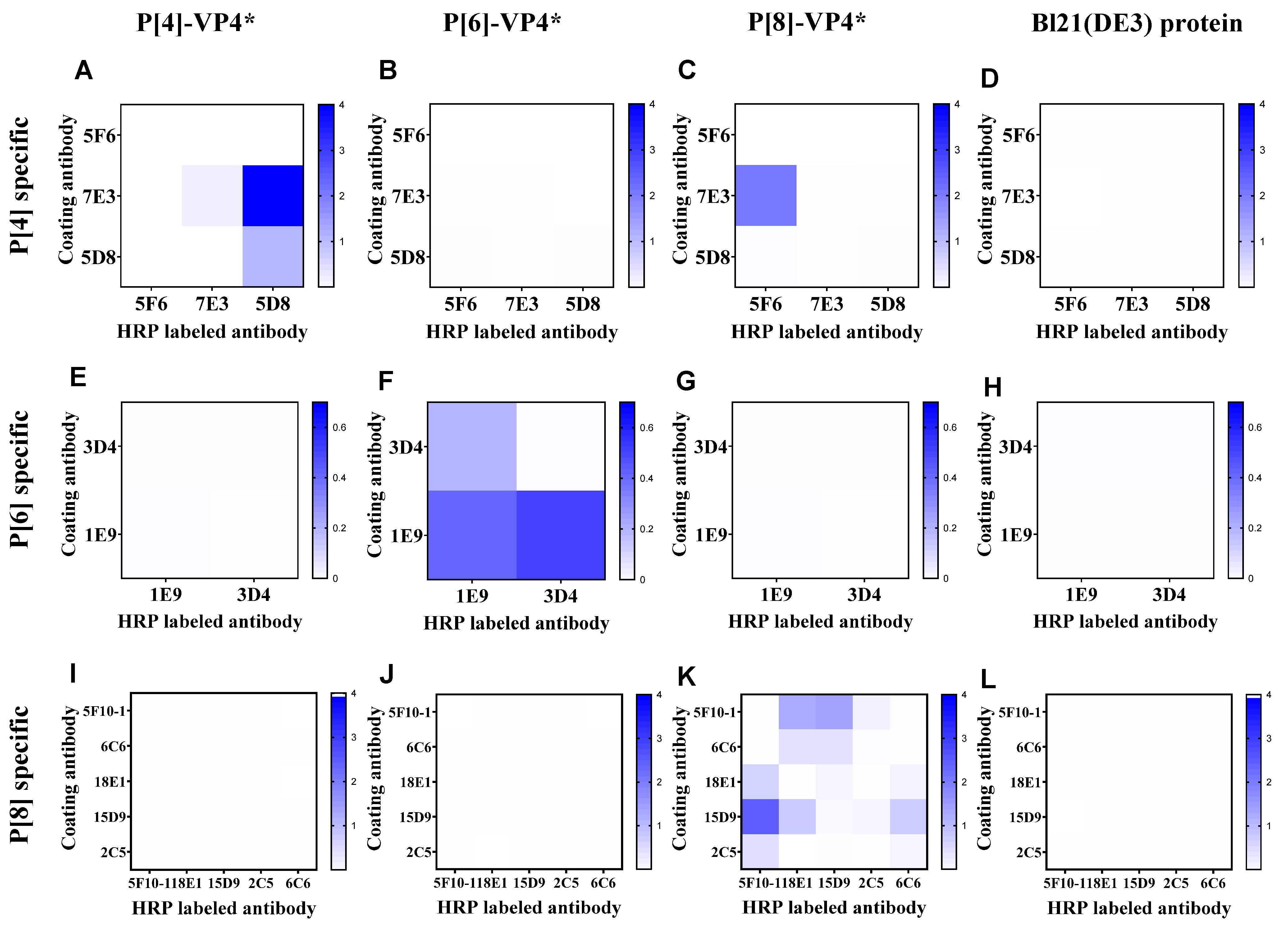
3.1. The Establishment of Double-Antibody Sandwich ELISA
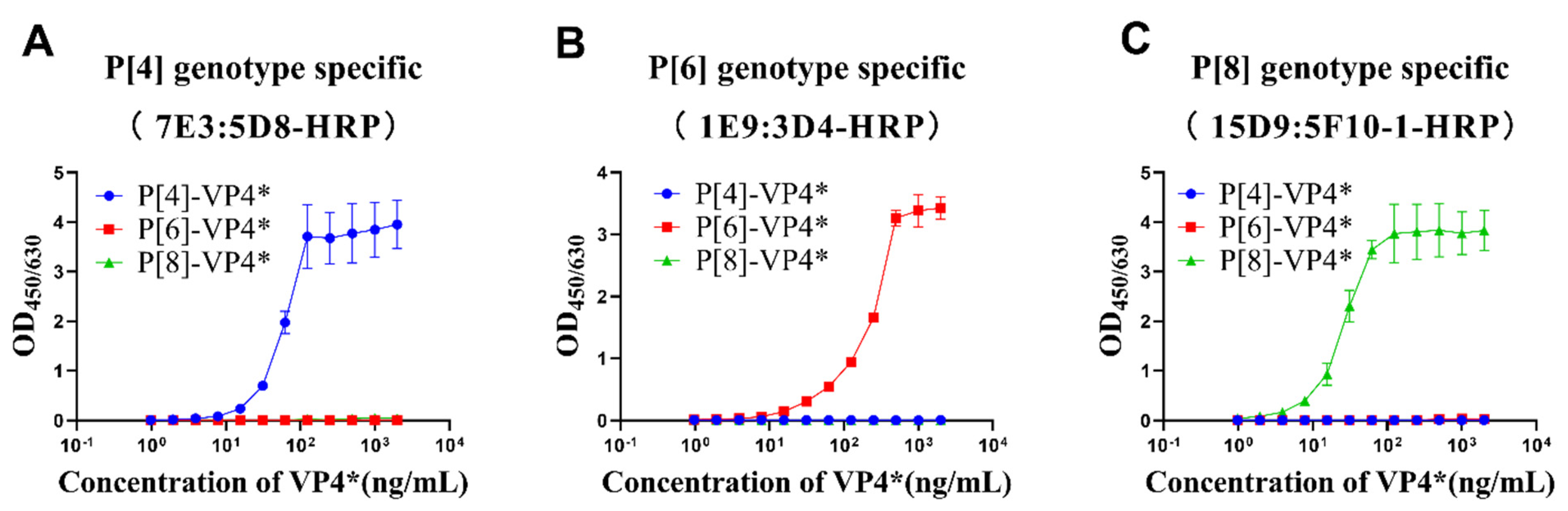
3.2. Detection of VP4* in E. coli Lysate by the Sandwich ELISAs
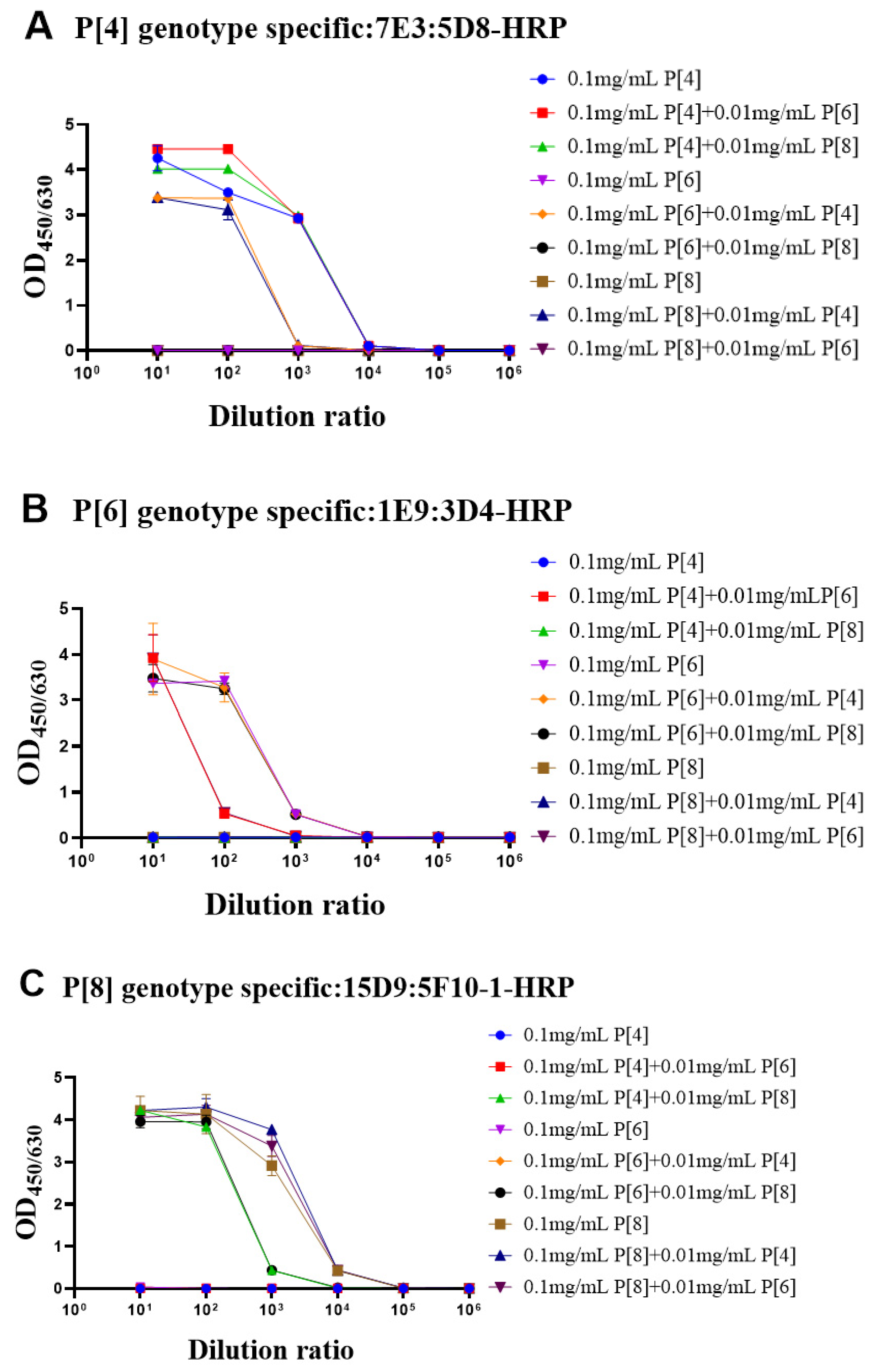
3.3. Identity Evaluation of Purified VP4* Proteins Using the Sandwich ELISAs
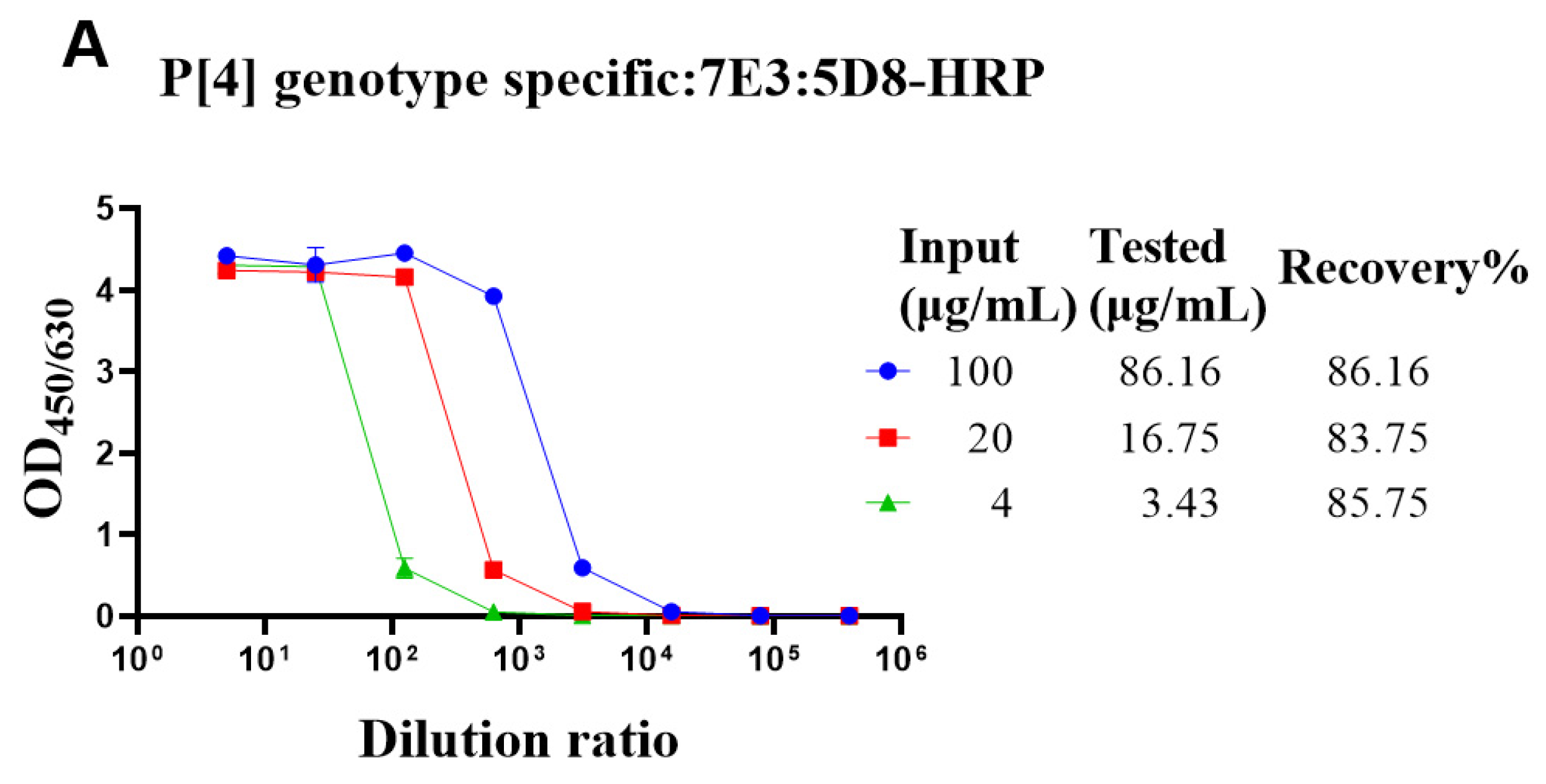
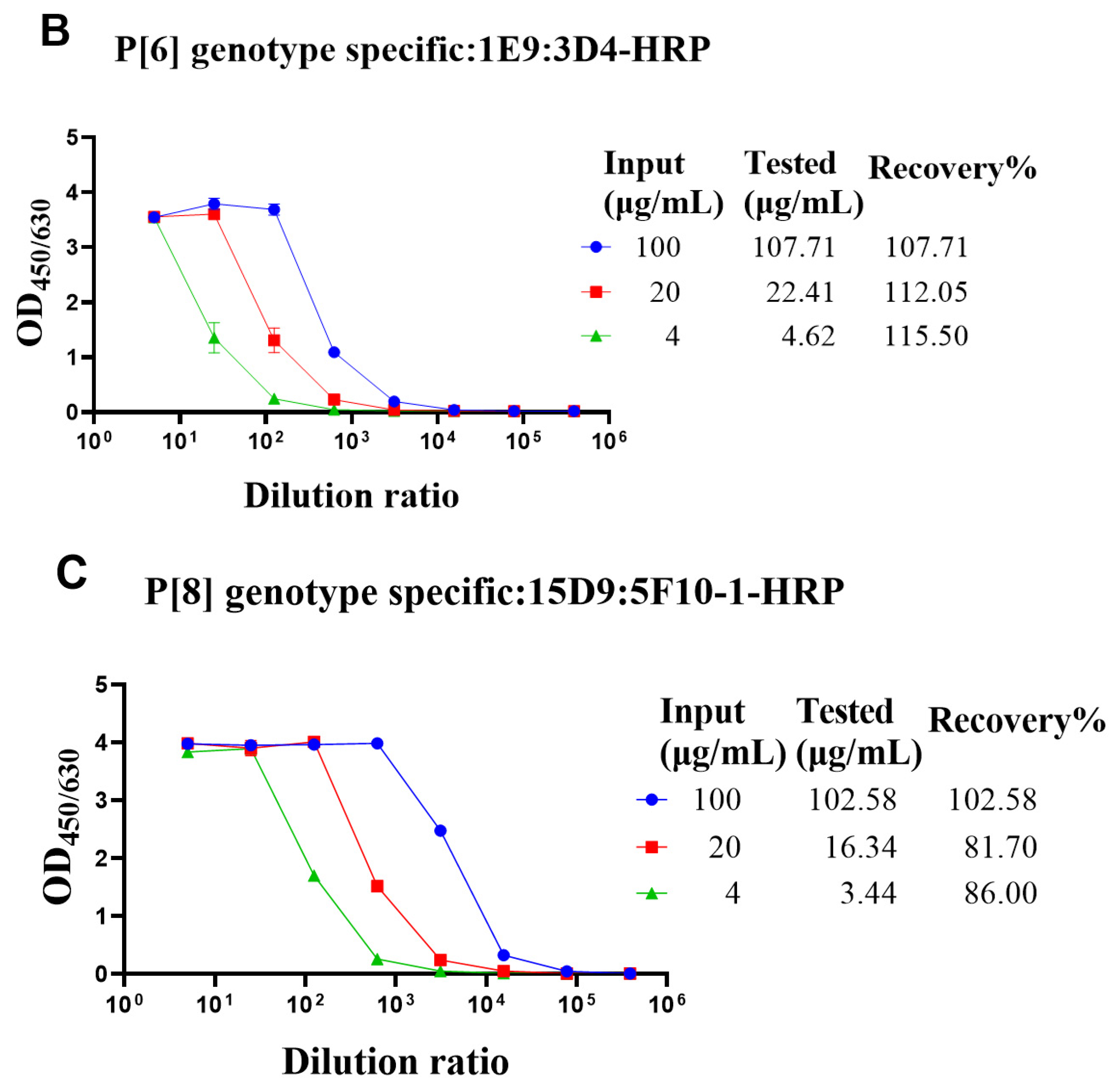
3.4. Determining the Adsorption Rate of VP4* Proteins on Aluminum Adjuvant
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Troeger, C.; Forouzanfar, M.; Rao, P.C.; Khalil, I.; Brown, A.; Reiner, R.C.; Fullman, N.; Thompson, R.L.; Abajobir, A.; Ahmed, M.; et al. Estimates of global, regional, and national morbidity, mortality, and aetiologies of diarrhoeal diseases: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect. Dis. 2017, 17, 909–948. [Google Scholar] [CrossRef] [Green Version]
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Parashar, U.D.; Global, W.H.O.-C. Global, Regional, and National Estimates of Rotavirus Mortality in Children <5 Years of Age, 2000–2013. Clin. Infect. Dis. 2016, 62, S96–S105. [Google Scholar]
- Parashar, U.D.; Glass, R.I. Rotavirus vaccines—Early success, remaining questions. N. Engl. J. Med. 2009, 360, 1063–1065. [Google Scholar] [CrossRef] [PubMed]
- Cunliffe, N.A.; Witte, D.; Ngwira, B.M.; Todd, S.; Bostock, N.J.; Turner, A.M.; Chimpeni, P.; Victor, J.C.; Steele, A.D.; Bouckenooghe, A.; et al. Efficacy of human rotavirus vaccine against severe gastroenteritis in Malawian children in the first two years of life: A randomized, double-blind, placebo controlled trial. Vaccine 2012, 30 (Suppl. 1), A36–A43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaman, K.; Anh, D.D.; Victor, J.C.; Shin, S.; Yunus, M.; Dallas, M.J.; Podder, G.; Thiem, V.D.; Mai, L.T.P.; Luby, S.P.; et al. Efficacy of pentavalent rotavirus vaccine against severe rotavirus gastroenteritis in infants in developing countries in Asia: A randomised, double-blind, placebo-controlled trial. Lancet 2010, 376, 615–623. [Google Scholar] [CrossRef]
- Yang, H.; Luo, G.X.; Zeng, Y.J.; Li, Y.J.; Yu, S.Y.; Zhao, B.Y.; An, R.; Zhang, S.Y.; Wang, Y.B.; Li, T.D.; et al. The distinct impact of maternal antibodies on the immunogenicity of live and recombinant rotavirus vaccines. Vaccine 2019, 37, 4061–4067. [Google Scholar] [CrossRef]
- Fix, A.D.; Harro, C.; McNeal, M.; Dally, L.; Flores, J.; Robertson, G.; Boslego, J.W.; Cryz, S. Safety and immunogenicity of a parenterally administered rotavirus VP8 subunit vaccine in healthy adults. Vaccine 2015, 33, 3766–3772. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Lin, H.J.; Zhang, Y.; Li, M.; Wang, D.N.; Che, Y.J.; Zhu, Y.B.; Li, S.W.; Zhang, J.; Ge, S.X.; et al. Improved characteristics and protective efficacy in an animal model of E-coli-derived recombinant double-layered rotavirus virus-like particles. Vaccine 2014, 32, 1921–1931. [Google Scholar] [CrossRef]
- Wen, X.; Cao, D.; Jones, R.W.; Li, J.; Szu, S.; Hoshino, Y. Construction and characterization of human rotavirus recombinant VP8* subunit parenteral vaccine candidates. Vaccine 2012, 30, 6121–6126. [Google Scholar] [CrossRef] [Green Version]
- Khodabandehloo, M.; Shahrabadi, M.S.; Keyvani, H.; Bambai, B.; Sadigh, Z.A. Recombinant Outer Capsid Glycoprotein (VP7) of Rotavirus Expressed in Insect Cells Induces Neutralizing Antibodies in Rabbits. Iran. J. Public Health 2012, 41, 73–84. [Google Scholar]
- Lappalainen, S.; Tamminen, K.; Vesikari, T.; Blazevic, V. Comparative immunogenicity in mice of rotavirus VP6 tubular structures and virus-like particles. Hum. Vaccines Immunother. 2013, 9, 1991–2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Xue, M.; Yu, L.; Luo, G.; Yang, H.; Jia, L.; Zeng, Y.; Li, T.; Ge, S.; Xia, N. Expression and characterization of a novel truncated rotavirus VP4 for the development of a recombinant rotavirus vaccine. Vaccine 2018, 36, 2086–2092. [Google Scholar] [CrossRef] [PubMed]
- Offit, P.A. Rotaviruses—Immunological Determinants of Protection against Infection and Disease. Adv. Virus Res. 1994, 44, 161–202. [Google Scholar] [PubMed]
- Trask, S.; McDonald, S.; Patton, J. Structural insights into the coupling of virion assembly and rotavirus replication. Nat. Rev. Microbiol. 2012, 10, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Li, J.X.; Wang, H.; Li, D.D.; Zhang, Q.; Liu, N. Infection status and circulating strains of rotaviruses in Chinese children younger than 5-years old from 2011 to 2018: Systematic review and meta-analysis. Hum. Vaccines Immunother. 2021, 17, 1811–1817. [Google Scholar] [CrossRef]
- Seheri, L.M.; Magagula, N.B.; Peenze, I.; Rakau, K.; Ndadza, A.; Mwenda, J.M.; Weldegebriel, G.; Steele, A.D.; Mphahlele, M.J. Rotavirus strain diversity in Eastern and Southern African countries before and after vaccine introduction. Vaccine 2018, 36, 7222–7230. [Google Scholar] [CrossRef]
- Abebe, A.; Getahun, M.; Mapaseka, S.L.; Beyene, B.; Assefa, E.; Teshome, B.; Tefera, M.; Kebede, F.; Habtamu, A.; Haile-Mariam, T.; et al. Impact of rotavirus vaccine introduction and genotypic characteristics of rotavirus strains in children less than 5 years of age with gastroenteritis in Ethiopia: 2011–2016. Vaccine 2018, 36, 7043–7047. [Google Scholar] [CrossRef]
- Damtie, D.; Melku, M.; Tessema, B.; Vlasova, A.N. Prevalence and Genetic Diversity of Rotaviruses among under-Five Children in Ethiopia: A Systematic Review and Meta-Analysis. Viruses 2020, 12, 62. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, S.; Hickey, J.; McAdams, D.; White, J.; Sitrin, R.; Khandke, L.; Cryz, S.; Joshi, S.; Volkin, D. Effect of Aluminum Adjuvant and Preservatives on Structural Integrity and Physicochemical Stability Profiles of Three Recombinant Subunit Rotavirus Vaccine Antigens. J. Pharm. Sci. 2020, 109, 476–487. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Song, F.; Luo, G.; Yang, H.; Li, C.; Liu, W.; Li, T.; Zhang, S.; Wang, Y.; Ge, S.; et al. Generation and characterization of mouse monoclonal antibodies against the VP4 protein of Group A Human Rotaviruses. submitted.
- Xue, M.; Yu, L.; Che, Y.; Lin, H.; Zeng, Y.; Fang, M.; Li, T.; Ge, S.; Xia, N. Characterization and protective efficacy in an animal model of a novel truncated rotavirus VP8 subunit parenteral vaccine candidate. Vaccine 2015, 33, 2606–2613. [Google Scholar] [CrossRef]
- Chen, L.; Ruan, F.; Sun, Y.; Chen, H.; Liu, M.; Zhou, J.; Qin, K. Establishment of sandwich ELISA for detecting the H7 subtype influenza A virus. J. Med. Virol. 2019, 91, 1168–1171. [Google Scholar] [CrossRef] [PubMed]
- Nakane, P.K.; Kawaoi, A. Peroxidase-labeled antibody. A new method of conjugation. J. Histochem. Cytochem. 1974, 22, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- McAdams, D.; Lakatos, K.; Estrada, M.; Chen, D.; Plikaytis, B.; Sitrin, R.; White, J.A. Quantification of trivalent non-replicating rotavirus vaccine antigens in the presence of aluminum adjuvant. J. Immunol. Methods 2021, 494, 113056. [Google Scholar] [CrossRef] [PubMed]
- Soni, D.; Sahoo, I.; Mallya, A.D.; Kamthe, P.; Sahai, A.; Goel, S.K.; Kulkarni, P.S.; Dhere, R.M. Development of competitive inhibition ELISA as an effective potency test to analyze human rabies vaccines and assessment of the antigenic epitope of rabies glycoprotein. J. Immunol. Methods 2021, 492, 112939. [Google Scholar] [CrossRef]
- Harmsen, M.M.; Seago, J.; Perez, E.; Charleston, B.; Eble, P.L.; Dekker, A. Isolation of Single-Domain Antibody Fragments That Preferentially Detect Intact (146S) Particles of Foot-and-Mouth Disease virus for Use in Vaccine Quality Control. Front. Immunol. 2017, 8, 960. [Google Scholar] [CrossRef] [Green Version]
- Shank-Retzlaff, M.; Wang, F.; Morley, T.; Anderson, C.; Hamm, M.; Brown, M.; Rowland, K.; Pancari, G.; Zorman, J.; Lowe, R.; et al. Correlation between Mouse Potency and In Vitro Relative Potency for Human Papillomavirus Type 16 Virus-Like Particles and Gardasil (R) Vaccine Samples. Hum. Vaccines 2005, 1, 191–197. [Google Scholar] [CrossRef]
- Gonzalez, E.; Reyes, F.; Otero, O.; Camacho, F.; Cuello, M.; Ramirez, F.; Acevedo, R. Monoclonal Antibodies Against the Capsular Polysaccharides A, C, Y, W, and X of Neisseria meningitidis: A Platform for the Quality Control of Meningococcal Vaccines. Methods Mol. Biol. 2019, 1969, 181–203. [Google Scholar]
- Reyes, F.; Otero, O.; Camacho, F.; Amin, N.; Ramirez, F.; Valdes, Y.; Acevedo, R.; Garcia, L.; Cardoso, D.; Cuello, M. A novel monoclonal antibody to Neisseria meningitidis serogroup X capsular polysaccharide and its potential use in quantitation of meningococcal vaccines. Biologicals 2014, 42, 312–315. [Google Scholar] [CrossRef]
- Coombes, L.; Tierney, R.; Rigsby, P.; Sesardic, D.; Stickings, P. In vitro antigen ELISA for quality control of tetanus vaccines. Biologicals 2012, 40, 466–472. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine Composition Total VP4* (μg/mL) | P[4]-VP4* | P[6]-VP4* | P[8]-VP4* | P[4]+P[6]+P[8]-VP4* | ||||
|---|---|---|---|---|---|---|---|---|
| 60 | 120 | 60 | 120 | 60 | 120 | 180 | 360 | |
| Unadsorbed P[4]-VP4* (μg/mL) | 0.32 | 0.59 | / | / | / | / | 0.40 | 1.50 |
| Unadsorbed P[6]-VP4* (μg/mL) | / | / | 0.39 | 1.52 | / | / | 0.80 | 2.00 |
| Unadsorbed P[8]-VP4* (μg/mL) | / | / | / | / | 0.22 | 0.92 | 0.23 | 0.97 |
| Adsorption rate (%) | 99.47 | 99.51 | 99.35 | 98.73 | 99.63 | 99.23 | 99.21 | 98.76 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Luo, G.; Zeng, Y.; Song, F.; Yang, H.; Zhang, S.; Wang, Y.; Li, T.; Ge, S.; Xia, N. Establishment of Sandwich ELISA for Quality Control in Rotavirus Vaccine Production. Vaccines 2022, 10, 243. https://doi.org/10.3390/vaccines10020243
Li C, Luo G, Zeng Y, Song F, Yang H, Zhang S, Wang Y, Li T, Ge S, Xia N. Establishment of Sandwich ELISA for Quality Control in Rotavirus Vaccine Production. Vaccines. 2022; 10(2):243. https://doi.org/10.3390/vaccines10020243
Chicago/Turabian StyleLi, Cao, Guoxing Luo, Yuanjun Zeng, Feibo Song, Han Yang, Shiyin Zhang, Yingbin Wang, Tingdong Li, Shengxiang Ge, and Ningshao Xia. 2022. "Establishment of Sandwich ELISA for Quality Control in Rotavirus Vaccine Production" Vaccines 10, no. 2: 243. https://doi.org/10.3390/vaccines10020243
APA StyleLi, C., Luo, G., Zeng, Y., Song, F., Yang, H., Zhang, S., Wang, Y., Li, T., Ge, S., & Xia, N. (2022). Establishment of Sandwich ELISA for Quality Control in Rotavirus Vaccine Production. Vaccines, 10(2), 243. https://doi.org/10.3390/vaccines10020243