
Immunological Aspects of Human Papilloma Virus-Related Cancers Always Says, “I Am like a Box of Complexity, You Never Know What You Are Gonna Get”

 , , ,
, , , 
Abstract
:1. Introduction
1.1. Human Papillomavirus Structure
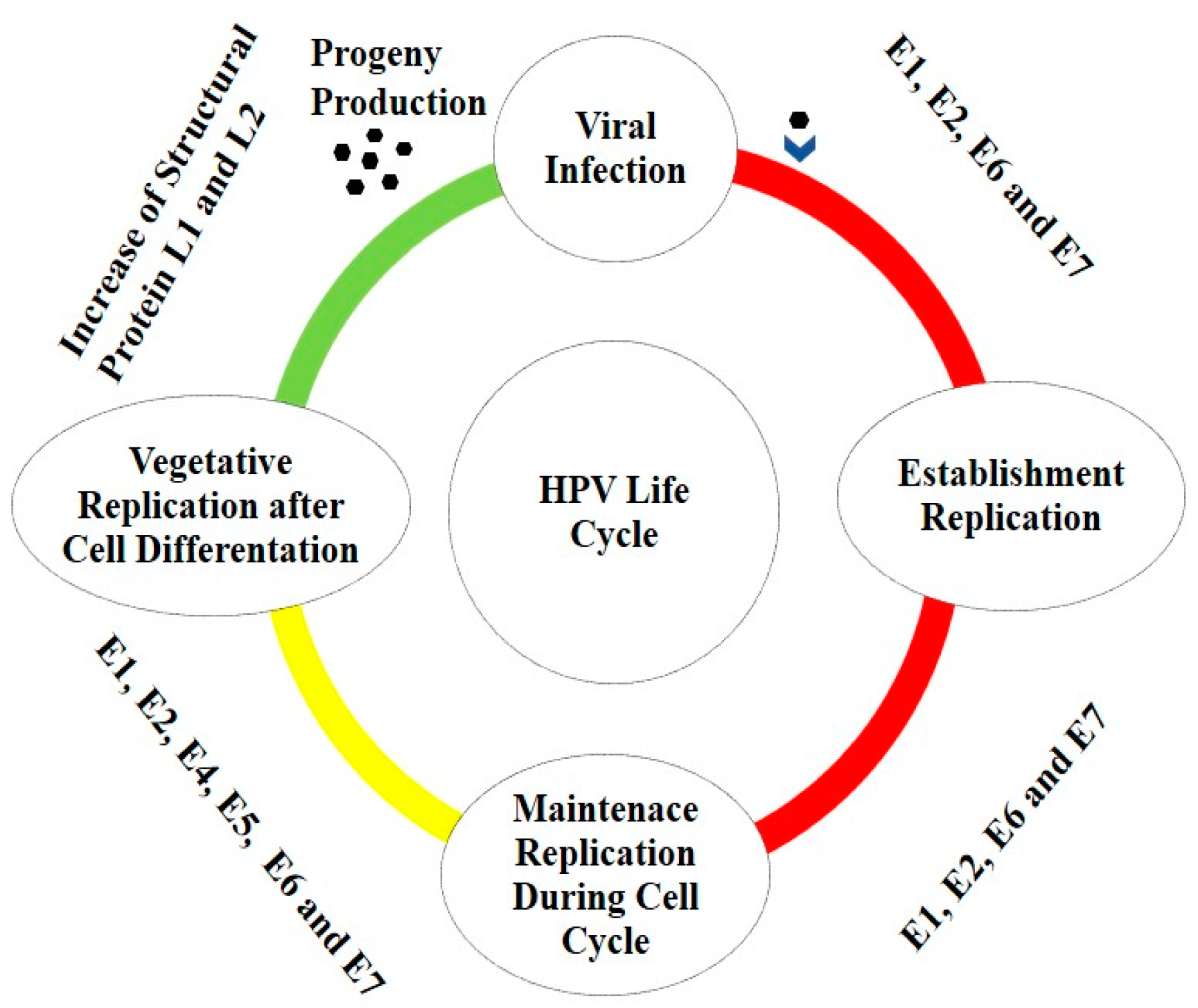
1.2. HPV Life Cycle
1.3. HPV Associated to Cancers
1.4. HPV and T Cells
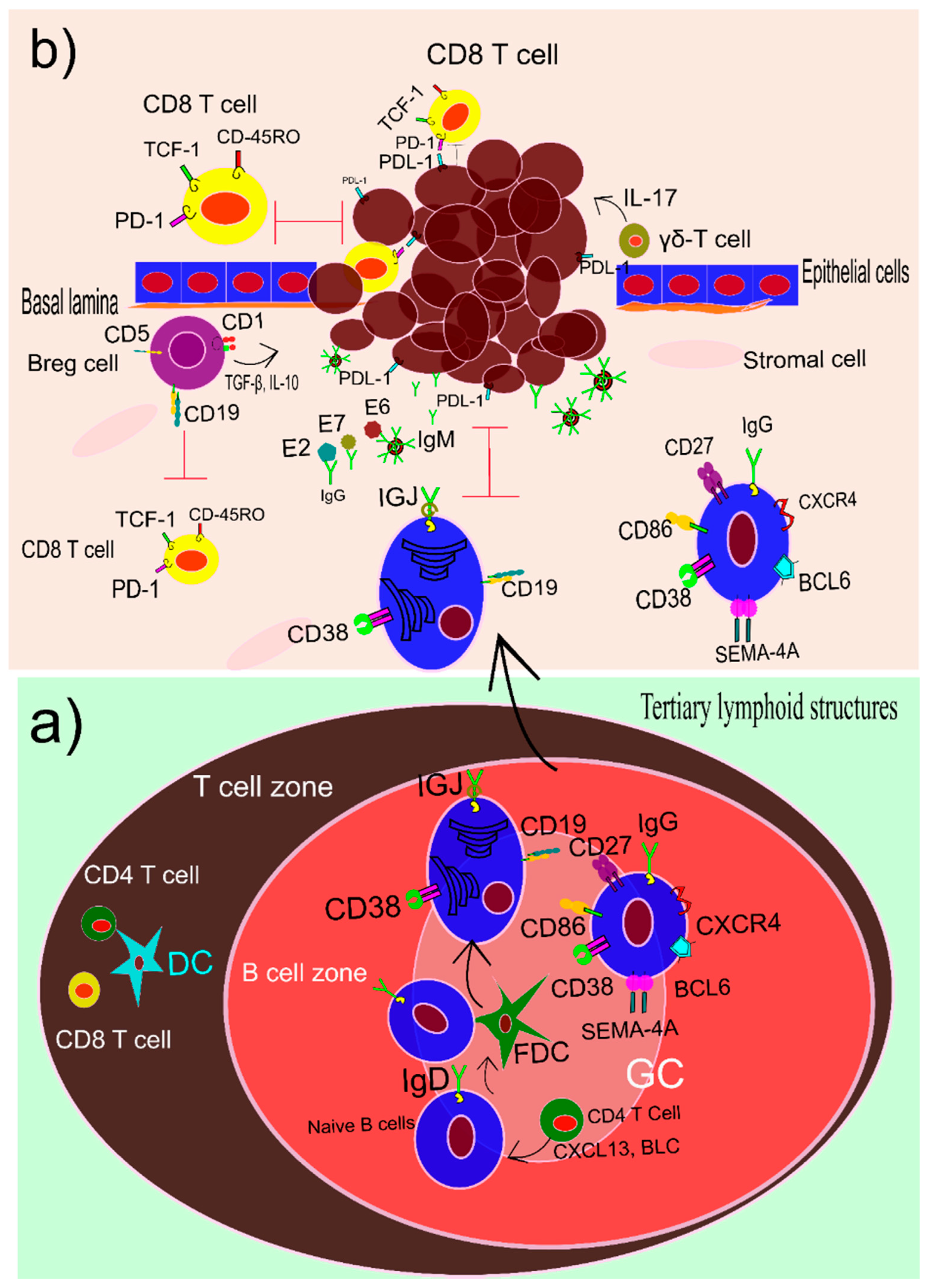
1.5. B Cells and HPV
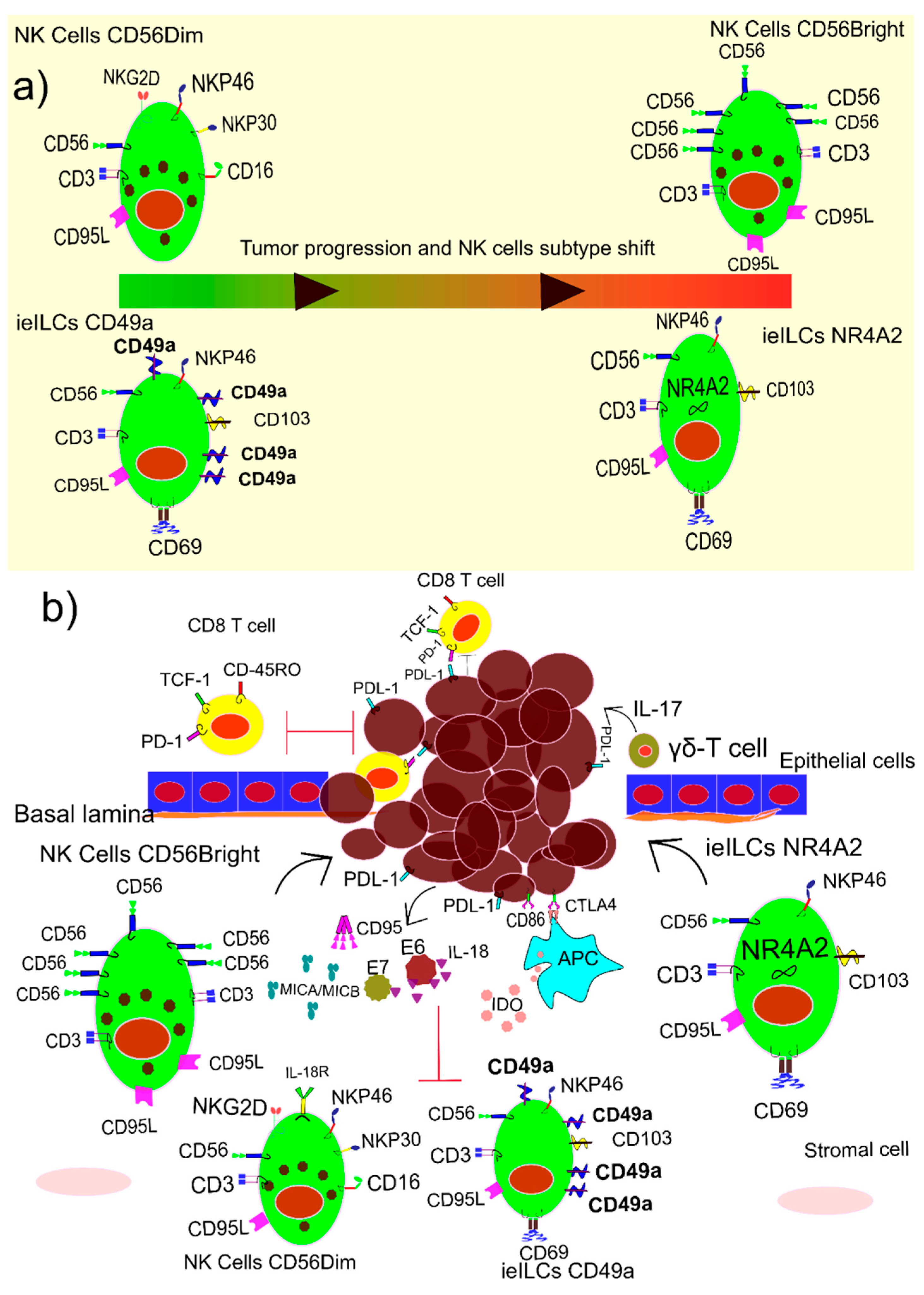
1.6. NK Cells and HPV
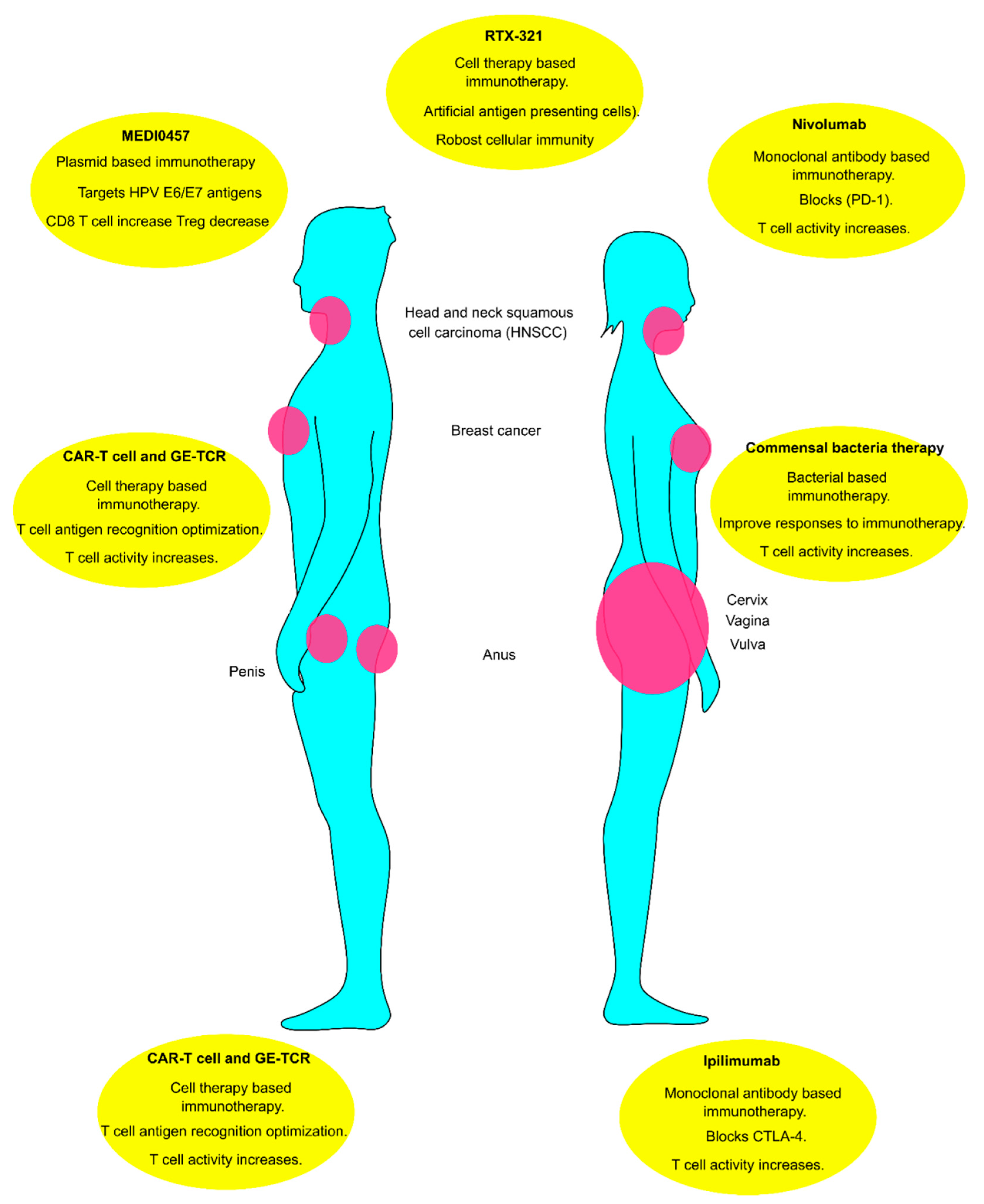
1.7. Immunotherapy of HPV-Related Tumors
2. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chesson, H.W.; Dunne, E.F.; Hariri, S.; Markowitz, L.E. The Estimated Lifetime Probability of Acquiring Human Papillomavirus in the United States. Sex. Transm. Dis. 2014, 41, 660. [Google Scholar] [CrossRef] [PubMed]
- Arrossi, S.; Almonte, M.; Herrero, R.; Gago, J.; Sánchez Antelo, V.; Szwarc, L.; Thouyaret, L.; Paolino, M.; Wiesner, C. Psycho-Social Impact of Positive Human Papillomavirus Testing in Jujuy, Argentina Results from the Psycho-Estampa Study. Prev. Med. Rep. 2020, 18, 101070. [Google Scholar] [CrossRef] [PubMed]
- Handler, M.Z.; Handler, N.S.; Majewski, S.; Schwartz, R.A. Human Papillomavirus Vaccine Trials and Tribulations: Clinical Perspectives. J. Am. Acad. Dermatol. 2015, 73, 743–756. [Google Scholar] [CrossRef] [PubMed]
- Walboomers, J.M.; Jacobs, M.V.; Manos, M.M.; Bosch, F.X.; Kummer, J.A.; Shah, K.V.; Snijders, P.J.; Peto, J.; Meijer, C.J.; Muñoz, N. Human Papillomavirus Is a Necessary Cause of Invasive Cervical Cancer Worldwide. J. Pathol. 1999, 189, 12–19. [Google Scholar] [CrossRef]
- Human Papillomavirus Vaccines: WHO Position Paper, May 2017-Recommendations. Vaccine 2017, 35, 5753–5755. [CrossRef]
- Roman, B.R.; Aragones, A. Epidemiology and Incidence of HPV-Related Cancers of the Head and Neck. J. Surg. Oncol. 2021, 124, 920–922. [Google Scholar] [CrossRef]
- Arbyn, M.; De Sanjosé, S.; Saraiya, M.; Sideri, M.; Palefsky, J.; Lacey, C.; Gillison, M.; Bruni, L.; Ronco, G.; Wentzensen, N.; et al. EUROGIN 2011 Roadmap on Prevention and Treatment of HPV-Related Disease. Int. J. Cancer 2012, 131, 1969–1982. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ding, J.; Zhai, K. Detection of Human Papillomavirus DNA in Patients with Breast Tumor in China. PLoS ONE 2015, 10, e0136050. [Google Scholar] [CrossRef] [Green Version]
- Heng, B.; Glenn, W.K.; Ye, Y.; Tran, B.; Delprado, W.; Lutze-Mann, L.; Whitaker, N.J.; Lawson, J.S. Human Papilloma Virus Is Associated with Breast Cancer. Br. J. Cancer 2009, 101, 1345–1350. [Google Scholar] [CrossRef]
- Sigaroodi, A.; Nadji, S.A.; Naghshvar, F.; Nategh, R.; Emami, H.; Velayati, A.A. Human Papillomavirus Is Associated with Breast Cancer in the North Part of Iran. Sci. World J. 2012, 2012, 837191. [Google Scholar] [CrossRef] [Green Version]
- Bedoya, A.M.; Jaramillo, R.; Baena, A.; Castaño, J.; Olaya, N.; Zea, A.H.; Herrero, R.; Sanchez, G.I. Location and Density of Immune Cells in Precursor Lesions and Cervical Cancer. Cancer Microenviron. 2013, 6, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Wielgos, A.; Pietrzak, B.; Sikora, M.; Martirosian, G.; Suchonska, B.; Gozdowska, J.; Oldakowska-Jedynak, U.; Jabiry-Zieniewicz, Z.; Durlik, M.; Rudnicka, L.; et al. Human Papillomavirus (HPV) DNA Detection Using Self-Sampling Devices in Women Undergoing Long Term Immunosuppressive Therapy. Viruses 2020, 12, 962. [Google Scholar] [CrossRef]
- Sager, R.; Frei, P.; Steiner, U.C.; Fink, D.; Betschart, C. Genital Dysplasia and Immunosuppression: Why Organ-Specific Therapy Is Important. Inflamm. Intest. Dis. 2019, 4, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Stelzle, D.; Tanaka, L.F.; Lee, K.K.; Ibrahim Khalil, A.; Baussano, I.; Shah, A.S.V.; McAllister, D.A.; Gottlieb, S.L.; Klug, S.J.; Winkler, A.S.; et al. Estimates of the Global Burden of Cervical Cancer Associated with HIV. Lancet Glob. Health 2021, 9, e161–e169. [Google Scholar] [CrossRef]
- Zur Hausen, H. Papillomaviruses and Cancer: From Basic Studies to Clinical Application. Nat. Rev. Cancer 2002, 2, 342–350. [Google Scholar] [CrossRef]
- Ding, D.C.; Chiang, M.H.; Lai, H.C.; Hsiung, C.A.; Hsieh, C.Y.; Chu, T.Y. Methylation of the Long Control Region of HPV16 Is Related to the Severity of Cervical Neoplasia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2009, 147, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Chaiwongkot, A.; Phanuphak, N.; Pankam, T.; Bhattarakosol, P. Human Papillomavirus 16 L1 Gene Methylation as a Potential Biomarker for Predicting Anal Intraepithelial Neoplasia in Men Who Have Sex with Men (MSM). PLoS ONE 2021, 16, e0256852. [Google Scholar] [CrossRef] [PubMed]
- Forcier, M.; Musacchio, N. An Overview of Human Papillomavirus Infection for the Dermatologist: Disease, Diagnosis, Management, and Prevention. Dermatol. Ther. 2010, 23, 458–476. [Google Scholar] [CrossRef]
- Major, A.G.; Pitty, L.P.; Farah, C.S. Cancer Stem Cell Markers in Head and Neck Squamous Cell Carcinoma. Stem Cells Int. 2013, 2013, 319489. [Google Scholar] [CrossRef]
- De Villiers, E.M.; Fauquet, C.; Broker, T.R.; Bernard, H.U.; Zur Hausen, H. Classification of Papillomaviruses. Virology 2004, 324, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Cubie, H.A. Diseases Associated with Human Papillomavirus Infection. Virology 2013, 445, 21–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Béziat, V. Human Genetic Dissection of Papillomavirus-Driven Diseases: New Insight into Their Pathogenesis. Hum. Genet. 2020, 139, 919–939. [Google Scholar] [CrossRef] [PubMed]
- Van Doorslaer, K.; Li, Z.; Xirasagar, S.; Maes, P.; Kaminsky, D.; Liou, D.; Sun, Q.; Kaur, R.; Huyen, Y.; McBride, A.A. The Papillomavirus Episteme: A Major Update to the Papillomavirus Sequence Database. Nucleic Acids Res. 2017, 45, D499–D506. [Google Scholar] [CrossRef]
- Stanley, M. Pathology and Epidemiology of HPV Infection in Females. Gynecol. Oncol. 2010, 117, S5. [Google Scholar] [CrossRef] [PubMed]
- Gheit, T. Mucosal and Cutaneous Human Papillomavirus Infections and Cancer Biology. Front. Oncol. 2019, 9, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunne, E.F.; Park, I.U. HPV and HPV-Associated Diseases. Infect. Dis. Clin. N. Am. 2013, 27, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Vinzón, S.E.; Rösl, F. HPV Vaccination for Prevention of Skin Cancer. Hum. Vaccines Immunother. 2015, 11, 353–357. [Google Scholar] [CrossRef] [Green Version]
- Derkay, C.S.; Bluher, A.E. Update on Recurrent Respiratory Papillomatosis. Otolaryngol. Clin. N. Am. 2019, 52, 669–679. [Google Scholar] [CrossRef]
- Egawa, N.; Egawa, K.; Griffin, H.; Doorbar, J. Human Papillomaviruses; Epithelial Tropisms, and the Development of Neoplasia. Viruses 2015, 7, 3863. [Google Scholar] [CrossRef] [Green Version]
- Yoon, C.S.; Kim, K.D.; Park, S.N.; Cheong, S.W. A6 Integrin Is the Main Receptor of Human Papillomavirus Type 16 VLP. Biochem. Biophys. Res. Commun. 2001, 283, 668–673. [Google Scholar] [CrossRef]
- Bousarghin, L.; Touzé, A.; Sizaret, P.-Y.; Coursaget, P. Human Papillomavirus Types 16, 31, and 58 Use Different Endocytosis Pathways to Enter Cells. J. Virol. 2003, 77, 3846–3850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schelhaas, M.; Shah, B.; Holzer, M.; Blattmann, P.; Kühling, L.; Day, P.M.; Schiller, J.T.; Helenius, A. Entry of Human Papillomavirus Type 16 by Actin-Dependent, Clathrin- and Lipid Raft-Independent Endocytosis. PLoS Pathog. 2012, 8, e1002657. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.R.; Ramalho, A.C.; Marques, M.; Ribeiro, D. The Interplay between Antiviral Signalling and Carcinogenesis in Human Papillomavirus Infections. Cancers 2020, 12, 646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosper, P.F.; Bradley, S.; Luo, L.; Kimple, R.J. Biology of HPV Mediated Carcinogenesis and Tumor Progression. Semin. Radiat. Oncol. 2021, 31, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Lugović-Mihić, L.; Djaković, H.C.I.; Kuna, M.; Šešerko, A. The Influence of Psychological Stress on HPV Infection Manifestations and Carcinogenesis. Cell. Physiol. Biochem. 2021, 55, 71–88. [Google Scholar] [CrossRef]
- Moody, C.A.; Laimins, L.A. Human Papillomavirus Oncoproteins: Pathways to Transformation. Nat. Rev. Cancer 2010, 10, 550–560. [Google Scholar] [CrossRef]
- Maglennon, G.A.; McIntosh, P.; Doorbar, J. Persistence of Viral DNA in the Epithelial Basal Layer Suggests a Model for Papillomavirus Latency Following Immune Regression. Virology 2011, 414, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Burchell, A.N.; Winer, R.L.; de Sanjosé, S.; Franco, E.L. Chapter 6: Epidemiology and Transmission Dynamics of Genital HPV Infection. Vaccine 2006, 24 (Suppl. 3), S52–S61. [Google Scholar] [CrossRef]
- O’Rourke, K. Cutaneous β-HPV May Predict Cutaneous Squamous Cell Carcinoma. Cancer 2022, 128, 646. [Google Scholar] [CrossRef]
- Egawa, N.; Doorbar, J. The Low-Risk Papillomaviruses. Virus Res. 2017, 231, 119–127. [Google Scholar] [CrossRef]
- Ryndock, E.J.; Meyers, C. A Risk for Non-Sexual Transmission of Human Papillomavirus? Expert Rev. Anti. Infect. Ther. 2014, 12, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Scott-Wittenborn, N.; Fakhry, C. Epidemiology of HPV Related Malignancies. Semin. Radiat. Oncol. 2021, 31, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Crosbie, E.J.; Einstein, M.H.; Franceschi, S.; Kitchener, H.C. Human Papillomavirus and Cervical Cancer. Lancet 2013, 382, 889–899. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Oton-Gonzalez, L.; Mazziotta, C.; Lanzillotti, C.; Iaquinta, M.R.; Tognon, M.; Martini, F. Simultaneous Detection and Viral DNA Load Quantification of Different Human Papillomavirus Types in Clinical Specimens by the High Analytical Droplet Digital PCR Method. Front. Microbiol. 2020, 11, 2818. [Google Scholar] [CrossRef] [PubMed]
- Lazare, C.; Xiao, S.; Meng, Y.; Wang, C.; Li, W.; Wang, Y.; Chen, G.; Wei, J.; Hu, J.; Xue, M.; et al. Evaluation of Cervical Intraepithelial Neoplasia Occurrence Following the Recorded Onset of Persistent High-Risk Human Papillomavirus Infection: A Retrospective Study on Infection Duration. Front. Oncol. 2019, 9, 976. [Google Scholar] [CrossRef]
- Maucort-Boulch, D.; Plummer, M.; Castle, P.E.; Demuth, F.; Safaeian, M.; Wheeler, C.M.; Schiffman, M. Predictors of Human Papillomavirus Persistence among Women with Equivocal or Mildly Abnormal Cytology. Int. J. Cancer 2010, 126, 684–691. [Google Scholar] [CrossRef]
- Li, K.; Yin, R.; Wang, D.; Li, Q. Human Papillomavirus Subtypes Distribution among 2309 Cervical Cancer Patients in West China. Oncotarget 2017, 8, 28502–28509. [Google Scholar] [CrossRef] [Green Version]
- Steele, J.C.; Mann, C.H.; Rookes, S.; Rollason, T.; Murphy, D.; Freeth, M.G.; Gallimore, P.H.; Roberts, S. T-Cell Responses to Human Papillomavirus Type 16 among Women with Different Grades of Cervical Neoplasia. Br. J. Cancer 2005, 93, 248–259. [Google Scholar] [CrossRef] [Green Version]
- Krishna, S.; Ulrich, P.; Wilson, E.; Parikh, F.; Narang, P.; Yang, S.; Read, A.K.; Kim-Schulze, S.; Park, J.G.; Posner, M.; et al. Human Papilloma Virus Specific Immunogenicity and Dysfunction of CD8 + T Cells in Head and Neck Cancer. Cancer Res. 2018, 78, 6159–6170. [Google Scholar] [CrossRef] [Green Version]
- Tawfeik, A.M.; Mora, A.; Osman, A.; Moneer, M.M.; El-Sheikh, N.; Elrefaei, M. Frequency of CD4+ Regulatory T Cells, CD8+ T Cells, and Human Papilloma Virus Infection in Egyptian Women with Breast Cancer. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420966822. [Google Scholar] [CrossRef]
- Béziat, V.; Rapaport, F.; Hu, J.; Titeux, M.; Bonnet des Claustres, M.; Bourgey, M.; Griffin, H.; Bandet, É.; Ma, C.S.; Sherkat, R.; et al. Humans with Inherited T Cell CD28 Deficiency Are Susceptible to Skin Papillomaviruses but Are Otherwise Healthy. Cell 2021, 184, 3812–3828.e30. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, C.S.; Kissick, H.T.; Patel, M.R.; Cardenas, M.A.; Prokhnevska, N.; Obeng, R.C.; Nasti, T.H.; Griffith, C.C.; Im, S.J.; Wang, X.; et al. Functional HPV-Specific PD-1 + Stem-like CD8 T Cells in Head and Neck Cancer. Nature 2021, 597, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Hewavisenti, R.; Ferguson, A.; Wang, K.; Jones, D.; Gebhardt, T.; Edwards, J.; Zhang, M.; Britton, W.; Yang, J.; Hong, A.; et al. CD103+ Tumor-Resident CD8+ T Cell Numbers Underlie Improved Patient Survival in Oropharyngeal Squamous Cell Carcinoma. J. Immunother. Cancer 2020, 8, e000452. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, N.; Ding, M.; Yang, Y.; Wang, Z.; Huang, L.; Zhu, W.; Mellor, A.L.; Hou, X.; Zhou, C.; et al. CD40 Accelerates the Antigen-Specific Stem-Like Memory CD8+ T Cells Formation and Human Papilloma Virus (HPV)-Positive Tumor Eradication. Front. Immunol. 2020, 11, 1012. [Google Scholar] [CrossRef] [PubMed]
- Maskey, N.; Thapa, N.; Maharjan, M.; Shrestha, G.; Maharjan, N.; Cai, H.; Liu, S. Infiltrating CD4 and CD8 Lymphocytes in HPV Infected Uterine Cervical Milieu. Cancer Manag. Res. 2019, 11, 7647–7655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litwin, T.R.; Irvin, S.R.; Chornock, R.L.; Sahasrabuddhe, V.V.; Stanley, M.; Wentzensen, N. Infiltrating T-Cell Markers in Cervical Carcinogenesis: A Systematic Review and Meta-Analysis. Br. J. Cancer 2020, 124, 831–841. [Google Scholar] [CrossRef]
- Chen, Z.; Dong, D.; Zhu, Y.; Pang, N.; Ding, J. The Role of Tim-3/Galectin-9 Pathway in T-Cell Function and Prognosis of Patients with Human Papilloma Virus-Associated Cervical Carcinoma. FASEB J. 2021, 35, e21401. [Google Scholar] [CrossRef]
- Ao, C.; Zeng, K. The Role of Regulatory T Cells in Pathogenesis and Therapy of Human Papillomavirus-Related Diseases, Especially in Cancer. Infect. Genet. Evol. 2018, 65, 406–413. [Google Scholar] [CrossRef]
- Liu, C.; Lu, J.; Tian, H.; Du, W.; Zhao, L.; Feng, J.; Yuan, D.; Li, Z. Increased Expression of PD-L1 by the Human Papillomavirus 16 E7 Oncoprotein Inhibits Anticancer Immunity. Mol. Med. Rep. 2017, 15, 1063–1070. [Google Scholar] [CrossRef] [Green Version]
- Raverdeau, M.; Cunningham, S.P.; Harmon, C.; Lynch, L. Γδ T Cells in Cancer: A Small Population of Lymphocytes with Big Implications. Clin. Transl. Immunol. 2019, 8, e01080. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, B.; Ma, F.; Tong, F.; Yan, B.; Liu, T.; Xie, H.; Song, L.; Yu, S.; Wei, L. Characteristics of B Lymphocyte Infiltration in HPV + Head and Neck Squamous Cell Carcinoma. Cancer Sci. 2021, 112, 1402–1416. [Google Scholar] [CrossRef] [PubMed]
- Gui, S.; O’Neill, W.Q.; Teknos, T.N.; Pan, Q. Plasma Cell Marker, Immunoglobulin J Polypeptide, Predicts Early Disease-Specific Mortality in HPV+ HNSCC. J. Immunother. Cancer 2021, 9, e001259. [Google Scholar] [CrossRef] [PubMed]
- Ruffin, A.T.; Cillo, A.R.; Tabib, T.; Liu, A.; Onkar, S.; Kunning, S.R.; Lampenfeld, C.; Atiya, H.I.; Abecassis, I.; Kürten, C.H.L.; et al. B Cell Signatures and Tertiary Lymphoid Structures Contribute to Outcome in Head and Neck Squamous Cell Carcinoma. Nat. Commun. 2021, 12, 3349. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Shen, S.; Miyauchi, S.; Dominick Sanders, P.; Franiak-Pietryga, I.; Mell, L.; Silvio Gutkind, J.; Cohen, E.E.W.; Califano, J.A.; Sharabi, A.B. B Cells Improve Overall Survival in HPV-Associated Squamous Cell Carcinomas and Are Activated by Radiation and PD-1 Blockade. Clin. Cancer Res. 2020, 26, 3345–3359. [Google Scholar] [CrossRef] [Green Version]
- Wieland, A.; Patel, M.R.; Cardenas, M.A.; Eberhardt, C.S.; Hudson, W.H.; Obeng, R.C.; Griffith, C.C.; Wang, X.; Chen, Z.G.; Kissick, H.T.; et al. Defining HPV-Specific B Cell Responses in Patients with Head and Neck Cancer. Nature 2021, 597, 274–278. [Google Scholar] [CrossRef]
- Meng, J.W.; Song, J.H. Association between Interleukin-2, Interleukin-10, Secretory Immunoglobulin A and Immunoglobulin G Expression in Vaginal Fluid and Human Papilloma Virus Outcome in Patients with Cervical Lesions. Oncol. Lett. 2019, 18, 5543. [Google Scholar] [CrossRef] [Green Version]
- Hladíková, K.; Koucký, V.; Bouček, J.; Laco, J.; Grega, M.; Hodek, M.; Zábrodský, M.; Vošmik, M.; Rozkošová, K.; Vošmiková, H.; et al. Tumor-Infiltrating B Cells Affect the Progression of Oropharyngeal Squamous Cell Carcinoma via Cell-to-Cell Interactions with CD8 + T Cells. J. Immunother. Cancer 2019, 7, 261. [Google Scholar] [CrossRef]
- Lechner, A.; Schlößer, H.A.; Thelen, M.; Wennhold, K.; Rothschild, S.I.; Gilles, R.; Quaas, A.; Siefer, O.G.; Huebbers, C.U.; Cukuroglu, E.; et al. Tumor-Associated B Cells and Humoral Immune Response in Head and Neck Squamous Cell Carcinoma. Oncoimmunology 2019, 8, 1535293. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhu, Y.; Du, R.; Pang, N.; Zhang, F.; Dong, D.; Ding, J.; Ding, Y. Role of Regulatory B Cells in the Progression of Cervical Cancer. Mediat. Inflamm. 2019, 2019, 6519427. [Google Scholar] [CrossRef]
- Loick, S.M.; Fröhlich, A.; Gabrielpillai, J.; Franzen, A.; Vogt, T.J.; Dietrich, J.; Wiek, C.; Scheckenbach, K.; Strieth, S.; Landsberg, J.; et al. DNA Methylation and MRNA Expression of OX40 (TNFRSF4) and GITR (TNFRSF18, AITR) in Head and Neck Squamous Cell Carcinoma Correlates with HPV Status, Mutational Load, an Interferon-γ Signature, Signatures of Immune Infiltrates, and Survival. J. Immunother. 2021, 45, 194–206. [Google Scholar] [CrossRef]
- Lisco, A.; Hsu, A.P.; Dimitrova, D.; Proctor, D.M.; Mace, E.M.; Ye, P.; Anderson, M.V.; Hicks, S.N.; Grivas, C.; Hammoud, D.A.; et al. Treatment of Relapsing HPV Diseases by Restored Function of Natural Killer Cells. N. Engl. J. Med. 2021, 385, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jin, S.; Li, X.; Liu, L.; Xi, L.; Wang, F.; Zhang, S. Human Papillomavirus Type 16 Disables the Increased Natural Killer Cells in Early Lesions of the Cervix. J. Immunol. Res. 2019, 2019, 9182979. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Hoya, A.; Soto-Cruz, I. NK Cell Regulation in Cervical Cancer and Strategies for Immunotherapy. Cells 2021, 10, 3104. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Nieves, U.Y.; Tay, J.K.; Saumyaa, S.; Horowitz, N.B.; Shin, J.H.; Mohammad, I.A.; Luca, B.; Mundy, D.C.; Gulati, G.S.; Bedi, N.; et al. Landscape of Innate Lymphoid Cells in Human Head and Neck Cancer Reveals Divergent NK Cell States in the Tumor Microenvironment. Proc. Natl. Acad. Sci. USA 2021, 118, e2101169118. [Google Scholar] [CrossRef] [PubMed]
- Cella, M.; Robinette, M.L. Intraepithelial ILC1-like Cells: Front-Line Fighters in Human Head and Neck Squamous Cell Carcinoma. Proc. Natl. Acad. Sci. USA 2021, 118, e2110875118. [Google Scholar] [CrossRef]
- Garcia-Iglesias, T.; del Toro-Arreola, A.; Albarran-Somoza, B.; del Toro-Arreola, S.; Sanchez-Hernandez, P.E.; Ramirez-Dueñas, M.; Balderas-Peña, L.M.A.; Bravo-Cuellar, A.; Ortiz-Lazareno, P.C.; Daneri-Navarro, A. Low NKp30, NKp46 and NKG2D Expression and Reduced Cytotoxic Activity on NK Cells in Cervical Cancer and Precursor Lesions. BMC Cancer 2009, 9, 186. [Google Scholar] [CrossRef] [Green Version]
- Julian, R.; Savani, M.; Bauman, J.E. Immunotherapy Approaches in HPV-Associated Head and Neck Cancer. Cancers 2021, 13, 5889. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, Q.; Zhang, Y.; Zhang, Q.; Zheng, Z.; Liu, S.; Liu, Z.; Meng, L.; Xin, Y.; Jiang, X. Immunotherapy Advances in Locally Advanced and Recurrent/Metastatic Head and Neck Squamous Cell Carcinoma and Its Relationship With Human Papillomavirus. Front. Immunol. 2021, 12, 2333. [Google Scholar] [CrossRef]
- Galvis, M.M.; Borges, G.A.; de Oliveira, T.B.; de Toledo, I.P.; Castilho, R.M.; Guerra, E.N.S.; Kowalski, L.P.; Squarize, C.H. Immunotherapy Improves Efficacy and Safety of Patients with HPV Positive and Negative Head and Neck Cancer: A Systematic Review and Meta-Analysis. Crit. Rev. Oncol. Hematol. 2020, 150, 102966. [Google Scholar] [CrossRef]
- Xu, Y.; Zhu, G.; Maroun, C.A.; Wu, I.X.Y.; Huang, D.; Seiwert, T.Y.; Liu, Y.; Mandal, R.; Zhang, X. Programmed Death-1/Programmed Death-Ligand 1-Axis Blockade in Recurrent or Metastatic Head and Neck Squamous Cell Carcinoma Stratified by Human Papillomavirus Status: A Systematic Review and Meta-Analysis. Front. Immunol. 2021, 12, 1118. [Google Scholar] [CrossRef]
- Aggarwal, C.; Cohen, R.B.; Morrow, M.P.; Kraynyak, K.A.; Sylvester, A.J.; Knoblock, D.M.; Bauml, J.M.; Weinstein, G.S.; Lin, A.; Boyer, J.; et al. Immunotherapy Targeting HPV 16/18 Generates Potent Immune Responses in HPV-Associated Head and Neck Cancer. Clin. Cancer Res. 2019, 25, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, J.J.; Levy, D.A.; Nguyen, S.A.; Knochelmann, H.M.; Day, T.A. Impact of PD-L1 Expression and Human Papillomavirus Status in Anti-PD1/PDL1 Immunotherapy for Head and Neck Squamous Cell Carcinoma-Systematic Review and Meta-Analysis. Head Neck 2020, 42, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Ferris, R.L.; Spanos, W.C.; Leidner, R.; Gonçalves, A.; Martens, U.M.; Kyi, C.; Sharfman, W.; Chung, C.H.; Devriese, L.A.; Gauthier, H.; et al. Original Research: Neoadjuvant Nivolumab for Patients with Resectable HPV-Positive and HPV-Negative Squamous Cell Carcinomas of the Head and Neck in the CheckMate 358 Trial. J. Immunother. Cancer 2021, 9, e002568. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Chen, L.; Song, Y.; Ma, L.; Xiao, P.; Chen, L.; Zhen, H.; Han, R.; Chen, X.; Sun, S.; et al. Induction of Co-Inhibitory Molecule CTLA-4 by Human Papillomavirus E7 Protein through Downregulation of Histone Methyltransferase JHDM1B Expression. Virology 2019, 538, 111–118. [Google Scholar] [CrossRef]
- Dorta-Estremera, S.; Hegde, V.L.; Slay, R.B.; Sun, R.; Yanamandra, A.V.; Nicholas, C.; Nookala, S.; Sierra, G.; Curran, M.A.; Sastry, K.J. Targeting Interferon Signaling and CTLA-4 Enhance the Therapeutic Efficacy of Anti-PD-1 Immunotherapy in Preclinical Model of HPV+ Oral Cancer. J. Immunother. Cancer 2019, 7, 252. [Google Scholar] [CrossRef]
- Doran, S.L.; Stevanović, S.; Adhikary, S.; Gartner, J.J.; Jia, L.; Kwong, M.L.M.; Faquin, W.C.; Hewitt, S.M.; Sherry, R.M.; Yang, J.C.; et al. T-Cell Receptor Gene Therapy for Human Papillomavirus–Associated Epithelial Cancers: A First-in-Human, Phase I/II Study. J. Clin. Oncol. 2019, 37, 2759. [Google Scholar] [CrossRef]
- Wang, X.; Sandberg, M.L.; Martin, A.D.; Negri, K.R.; Gabrelow, G.B.; Nampe, D.P.; Wu, M.L.; McElvain, M.E.; Toledo Warshaviak, D.; Lee, W.H.; et al. Potent, Selective CARs as Potential T-Cell Therapeutics for HPV-Positive Cancers. J. Immunother. 2021, 44, 292. [Google Scholar] [CrossRef]
- Nagarsheth, N.B.; Norberg, S.M.; Sinkoe, A.L.; Adhikary, S.; Meyer, T.J.; Lack, J.B.; Warner, A.C.; Schweitzer, C.; Doran, S.L.; Korrapati, S.; et al. TCR-Engineered T Cells Targeting E7 for Patients with Metastatic HPV-Associated Epithelial Cancers. Nat. Med. 2021, 27, 419–425. [Google Scholar] [CrossRef]
- Norberg, S.; Nagarsheth, N.; Sinkoe, A.; Adhikary, S.; Meyer, T.; Lack, J.; Kanakry, J.A.; Bagheri, M.; Schweitzer, C.; Astrow, S.H.; et al. Safety and Clinical Activity of Gene-Engineered T-Cell Therapy Targeting HPV-16 E7 for Epithelial Cancers. J. Clin. Oncol. 2020, 38, 101. [Google Scholar] [CrossRef]
- Luo, M.; Dastagir, S.S.; Zhang, X.; Schmidt, A.; Marques, B.; Lyford, T.J.; Blanco, B.; Turka, L.A.; Wickham, T.J.; Chen, T.F. Abstract PO044: RTX-321, an Allogeneic Red Blood Cell-Based Artificial Antigen Presenting Cell, Expressing MHC I-Peptide, 4-1BBL and IL-12, Engages Primary Human HPV-Specific T Cells and Boosts Other General Immune Responses. Cancer Immunol. Res. 2021, 9, PO044. [Google Scholar] [CrossRef]
- Doerflinger, S.Y.; Throop, A.L.; Herbst-Kralovetz, M.M. Bacteria in the Vaginal Microbiome Alter the Innate Immune Response and Barrier Properties of the Human Vaginal Epithelia in a Species-Specific Manner. J. Infect. Dis. 2014, 209, 1989–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakabayashi, R.; Nakahama, Y.; Nguyen, V.; Espinoza, J.L. The Host-Microbe Interplay in Human Papillomavirus-Induced Carcinogenesis. Microorganisms 2019, 7, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onywera, H.; Williamson, A.L.; Mbulawa, Z.Z.A.; Coetzee, D.; Meiring, T.L. The Cervical Microbiota in Reproductive-Age South African Women with and without Human Papillomavirus Infection. Papillomavirus Res. 2019, 7, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Lebeau, A.; Bruyere, D.; Roncarati, P.; Peixoto, P.; Hervouet, E.; Cobraiville, G.; Taminiau, B.; Masson, M.; Gallego, C.; Mazzucchelli, G.; et al. HPV Infection Alters Vaginal Microbiome through Down-Regulating Host Mucosal Innate Peptides Used by Lactobacilli as Amino Acid Sources. Nat. Commun. 2022, 13, 1076. [Google Scholar] [CrossRef]
- Matson, V.; Fessler, J.; Bao, R.; Chongsuwat, T.; Zha, Y.; Alegre, M.L.; Luke, J.J.; Gajewski, T.F. The Commensal Microbiome Is Associated with Anti-PD-1 Efficacy in Metastatic Melanoma Patients. Science 2018, 359, 104–108. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HPV Neoplasia Potential | High-Risk (HR) | Low-Risk (LR) |
|---|---|---|
| HPV Types | 16, 18, 23, 31, 33, 35, 39, 45, 51, 52, 53, 56, 58, 66, 68, 73, and 82 | 40, 42, 43, 44, 53, 54, 61, 72, 73, and 81 |
| Lesions | Intraepithelial neoplasia and cervical cancer | Intraepithelial neoplasia, genital or cutaneus warts |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soleymaninejadian, E.; Zelini, P.; Cassaniti, I.; Baldanti, F.; Dominoni, M.; Gritti, A.; Gardella, B. Immunological Aspects of Human Papilloma Virus-Related Cancers Always Says, “I Am like a Box of Complexity, You Never Know What You Are Gonna Get”. Vaccines 2022, 10, 731. https://doi.org/10.3390/vaccines10050731
Soleymaninejadian E, Zelini P, Cassaniti I, Baldanti F, Dominoni M, Gritti A, Gardella B. Immunological Aspects of Human Papilloma Virus-Related Cancers Always Says, “I Am like a Box of Complexity, You Never Know What You Are Gonna Get”. Vaccines. 2022; 10(5):731. https://doi.org/10.3390/vaccines10050731
Chicago/Turabian StyleSoleymaninejadian, Ehsan, Paola Zelini, Irene Cassaniti, Fausto Baldanti, Mattia Dominoni, Andrea Gritti, and Barbara Gardella. 2022. "Immunological Aspects of Human Papilloma Virus-Related Cancers Always Says, “I Am like a Box of Complexity, You Never Know What You Are Gonna Get”" Vaccines 10, no. 5: 731. https://doi.org/10.3390/vaccines10050731
APA StyleSoleymaninejadian, E., Zelini, P., Cassaniti, I., Baldanti, F., Dominoni, M., Gritti, A., & Gardella, B. (2022). Immunological Aspects of Human Papilloma Virus-Related Cancers Always Says, “I Am like a Box of Complexity, You Never Know What You Are Gonna Get”. Vaccines, 10(5), 731. https://doi.org/10.3390/vaccines10050731